
The Effects of Plant Health Status on the Community Structure and Metabolic Pathways of Rhizosphere Microbial Communities Associated with Solanum lycopersicum




Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Area
2.2. Field Description and Sampling
2.3. Extraction and Shotgun Sequencing of DNA Obtained from Soil Samples
2.4. Data Analysis
2.5. Statistical Analysis
3. Results
3.1. Shotgun Sequencing of the Microbiome in Tomato Rhizosphere and Bulk Soil
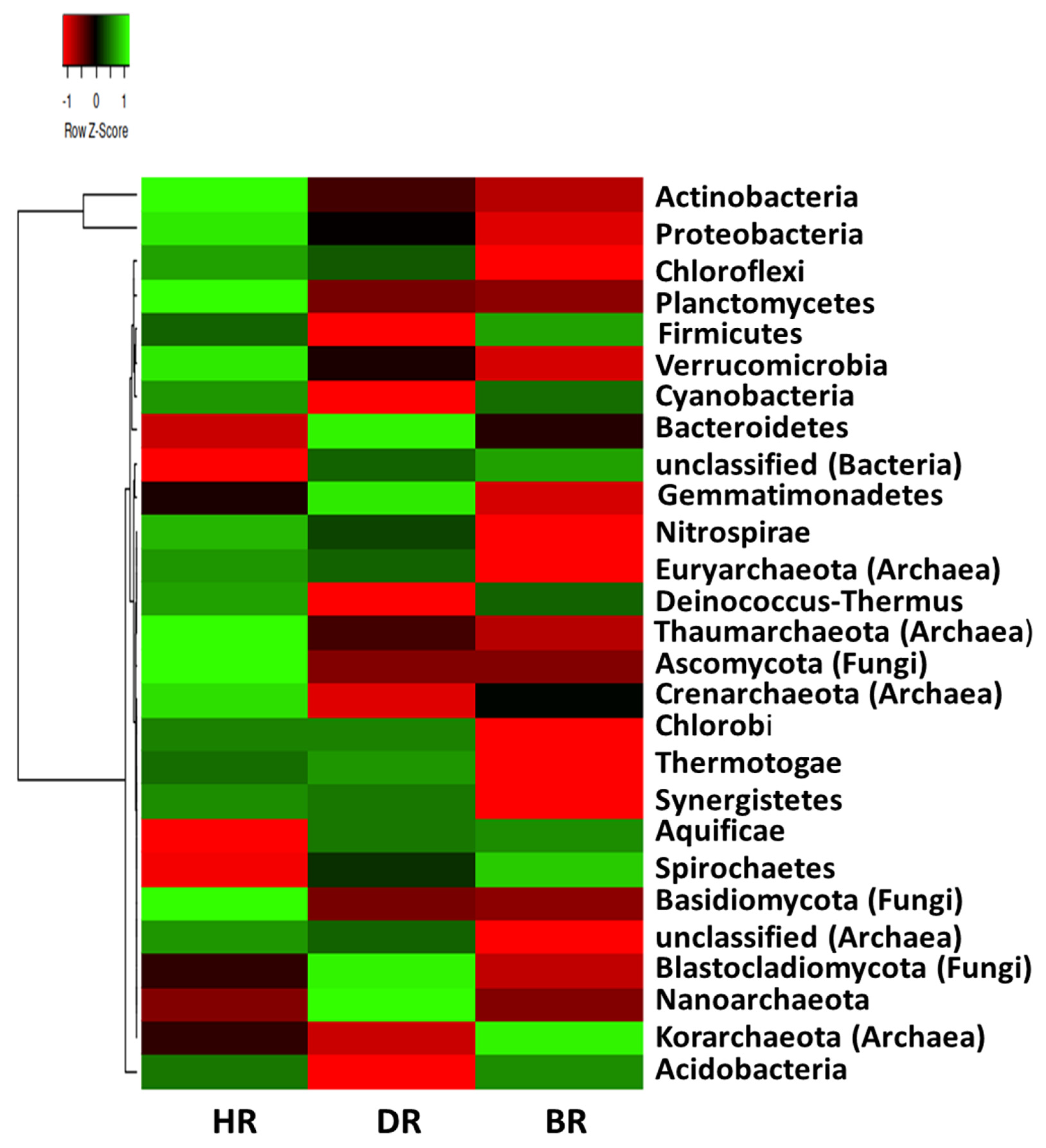
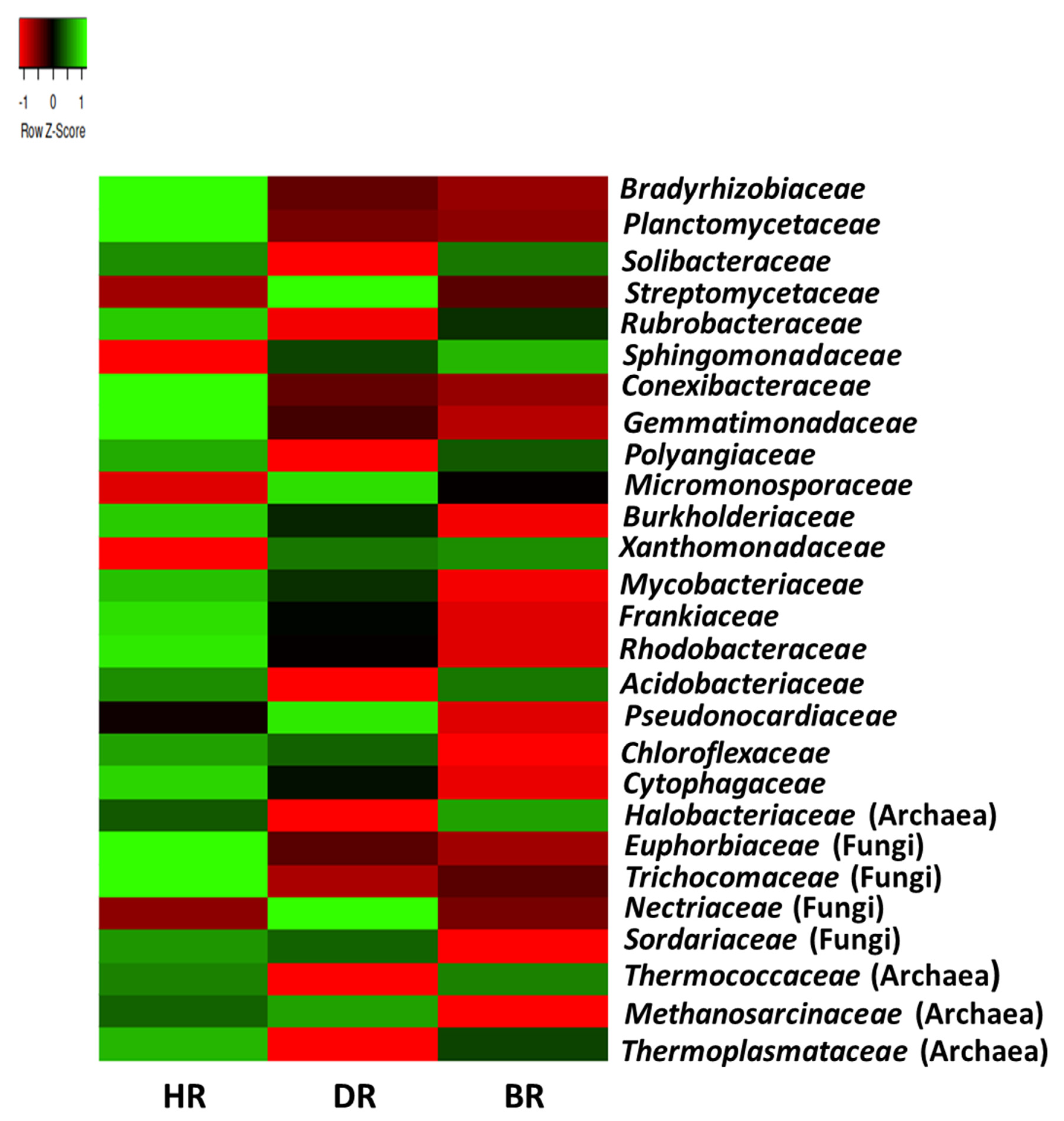
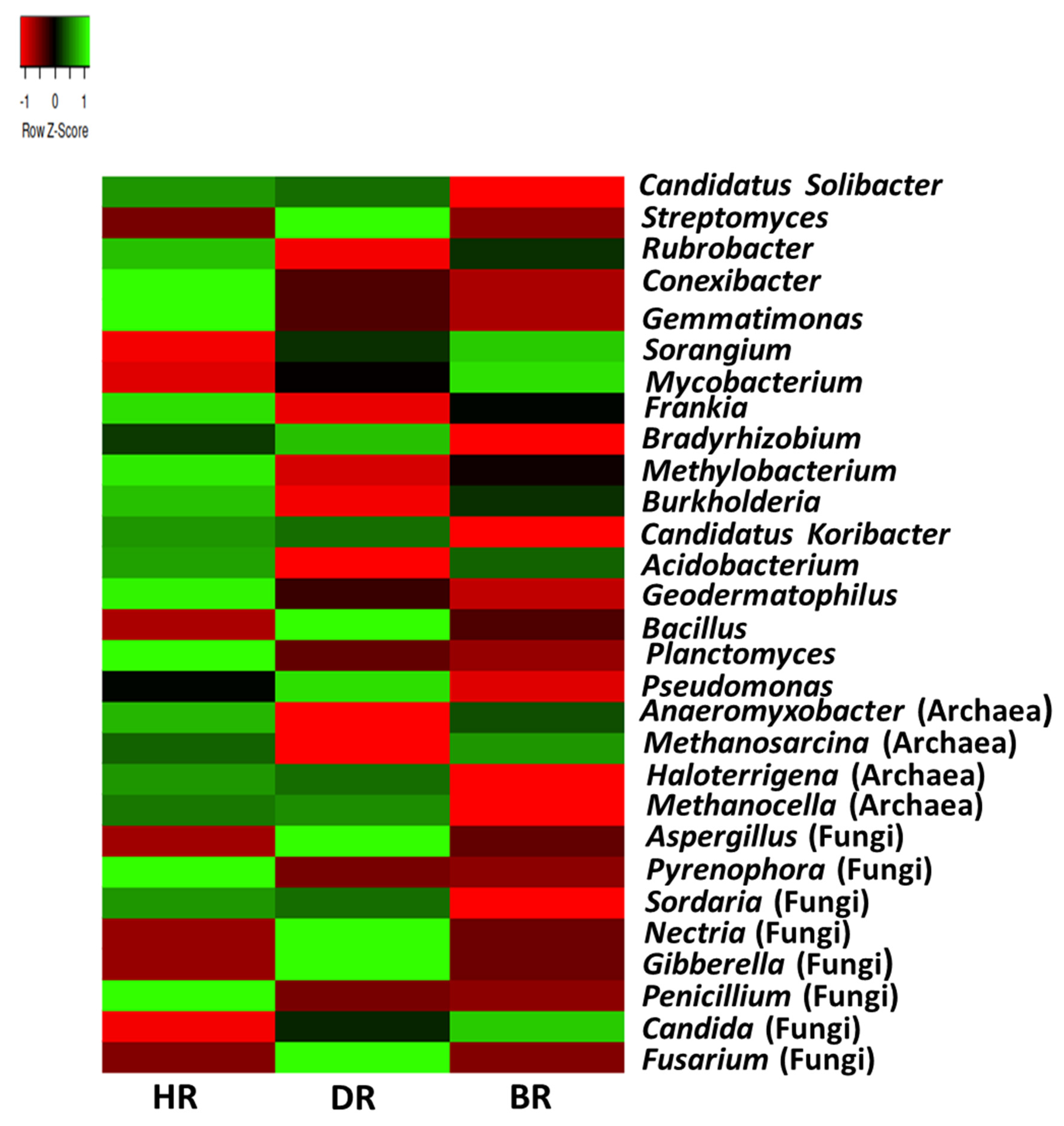
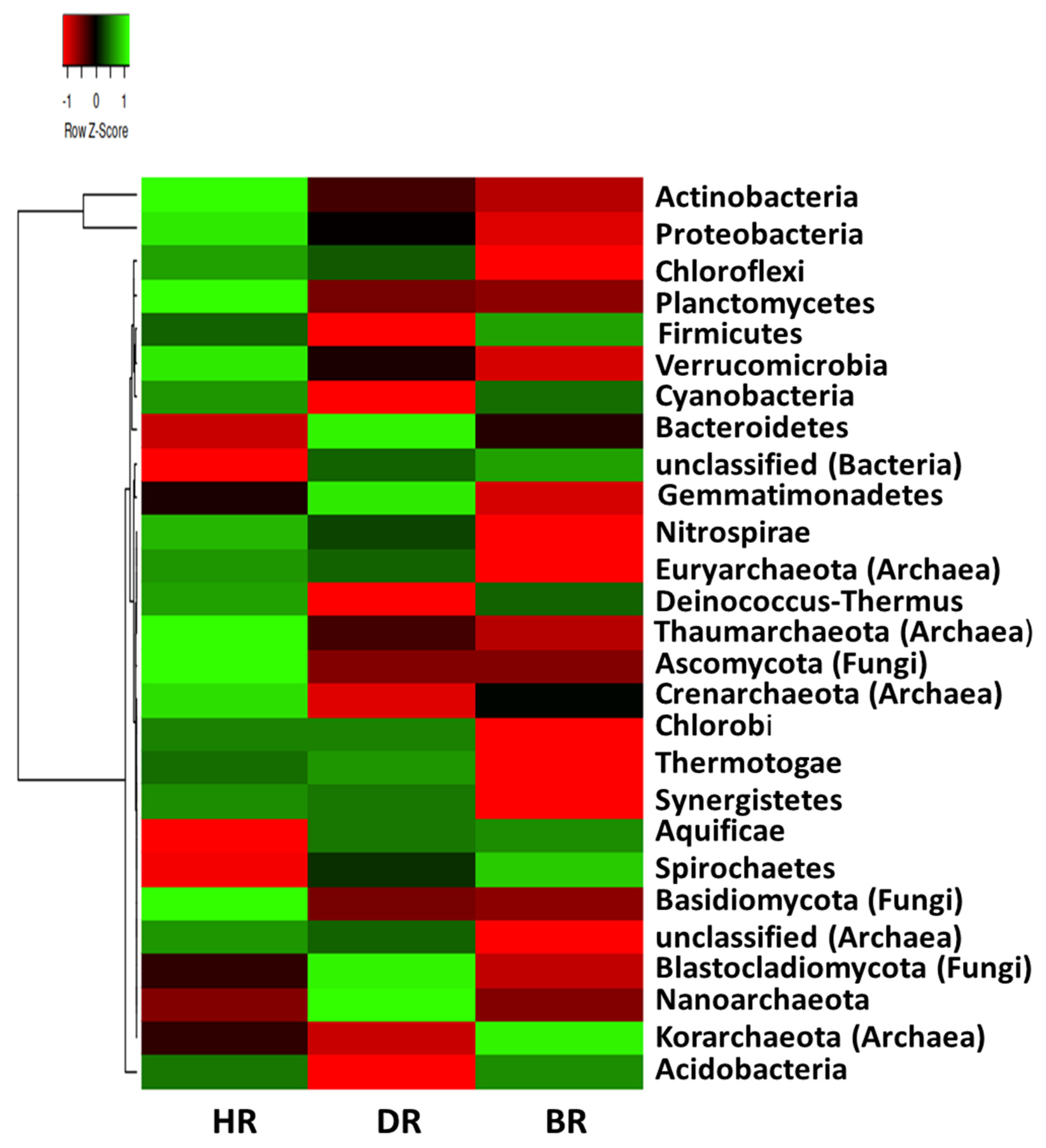
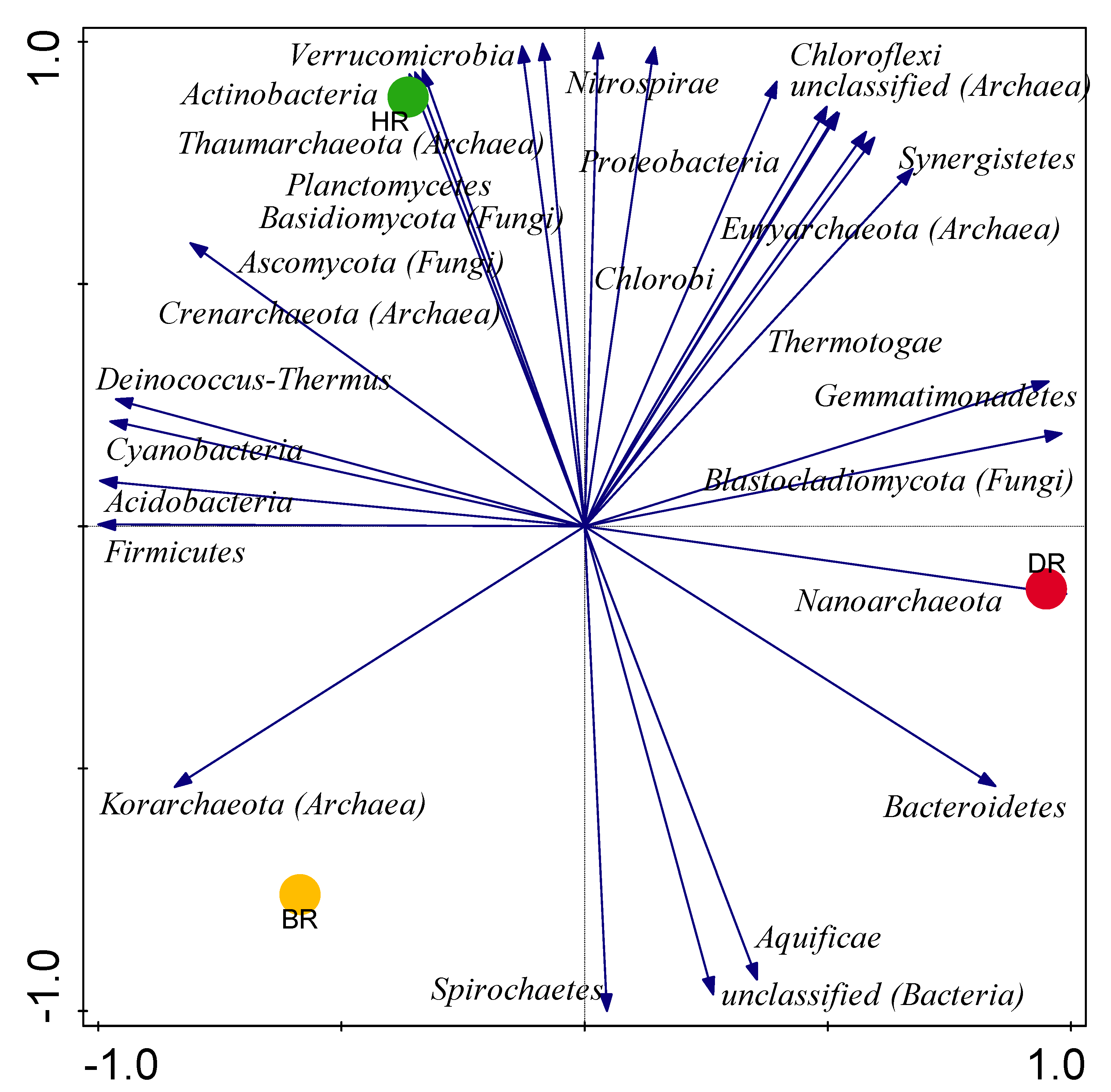
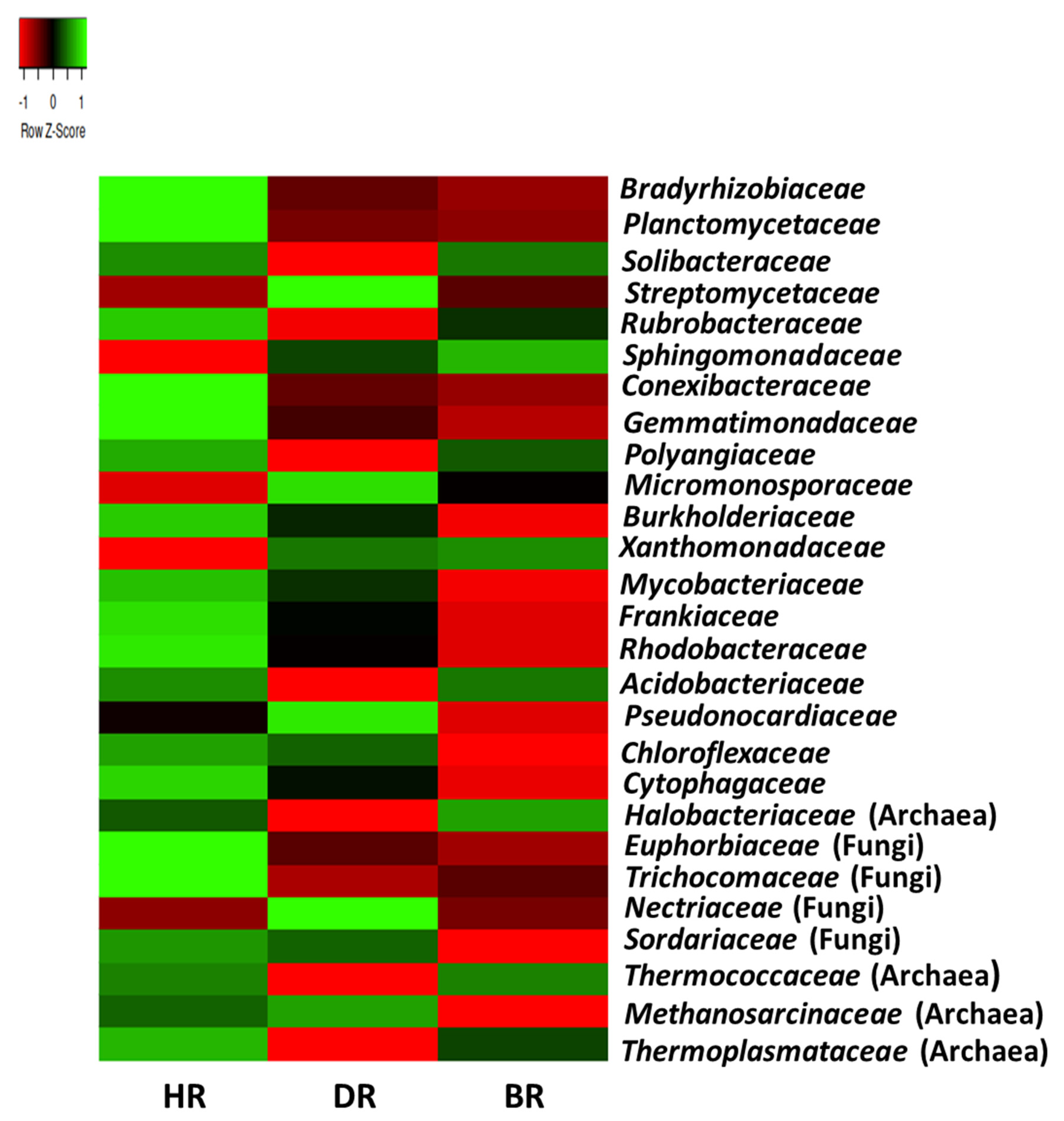
3.2. Distribution of Major Rhizosphere Soil Microbiome Phyla across the Tomato Plant Sites
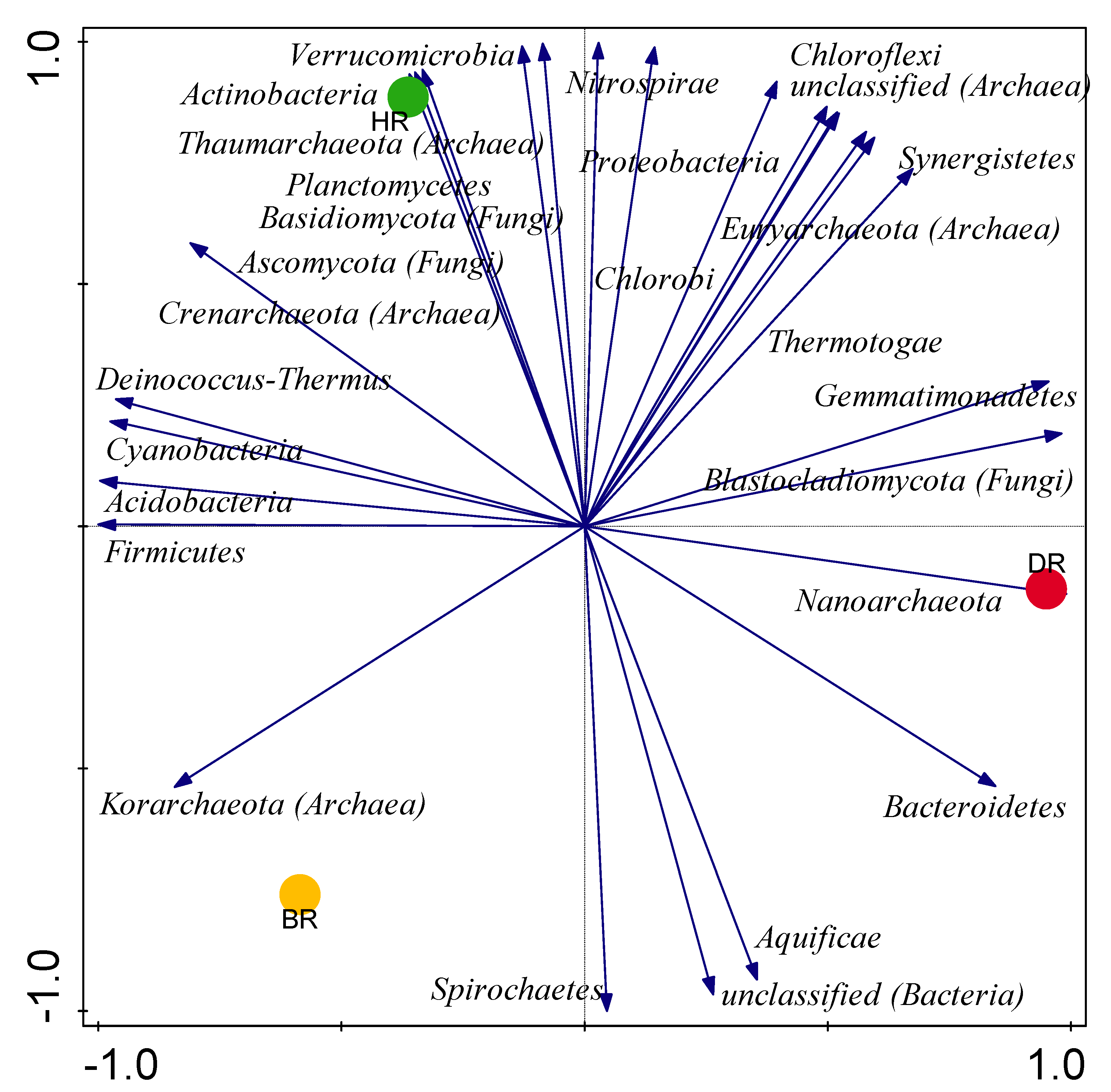
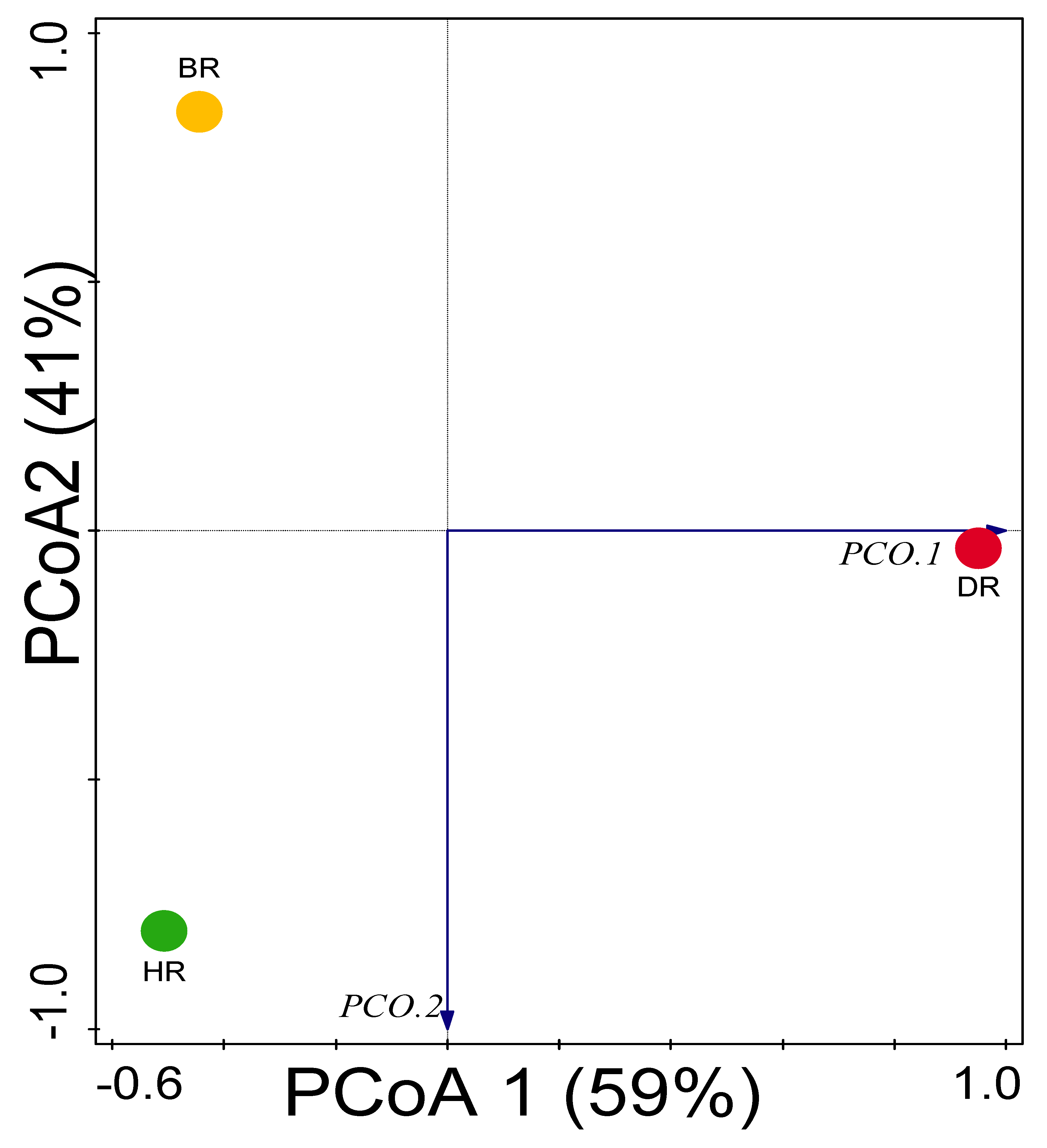
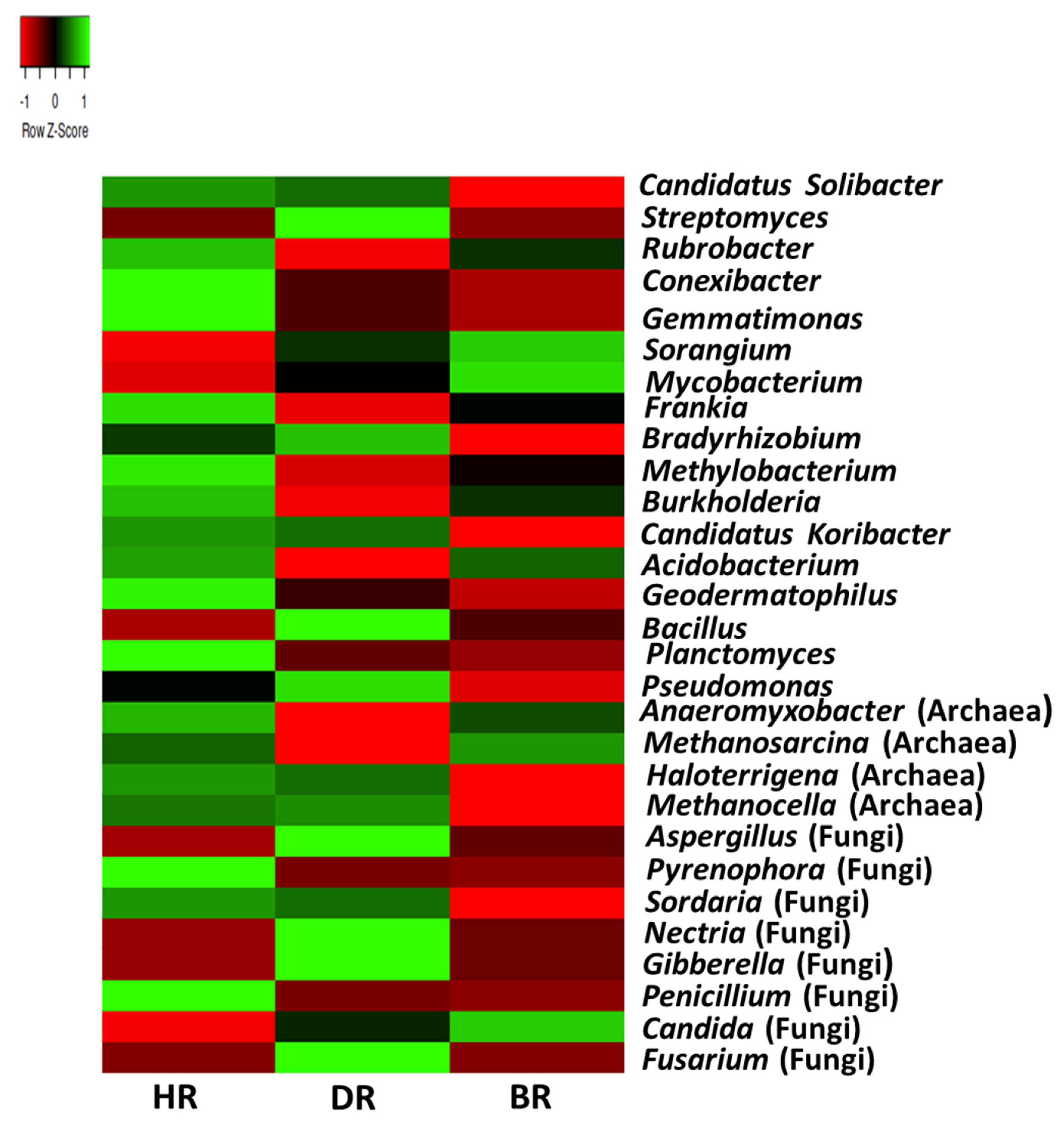
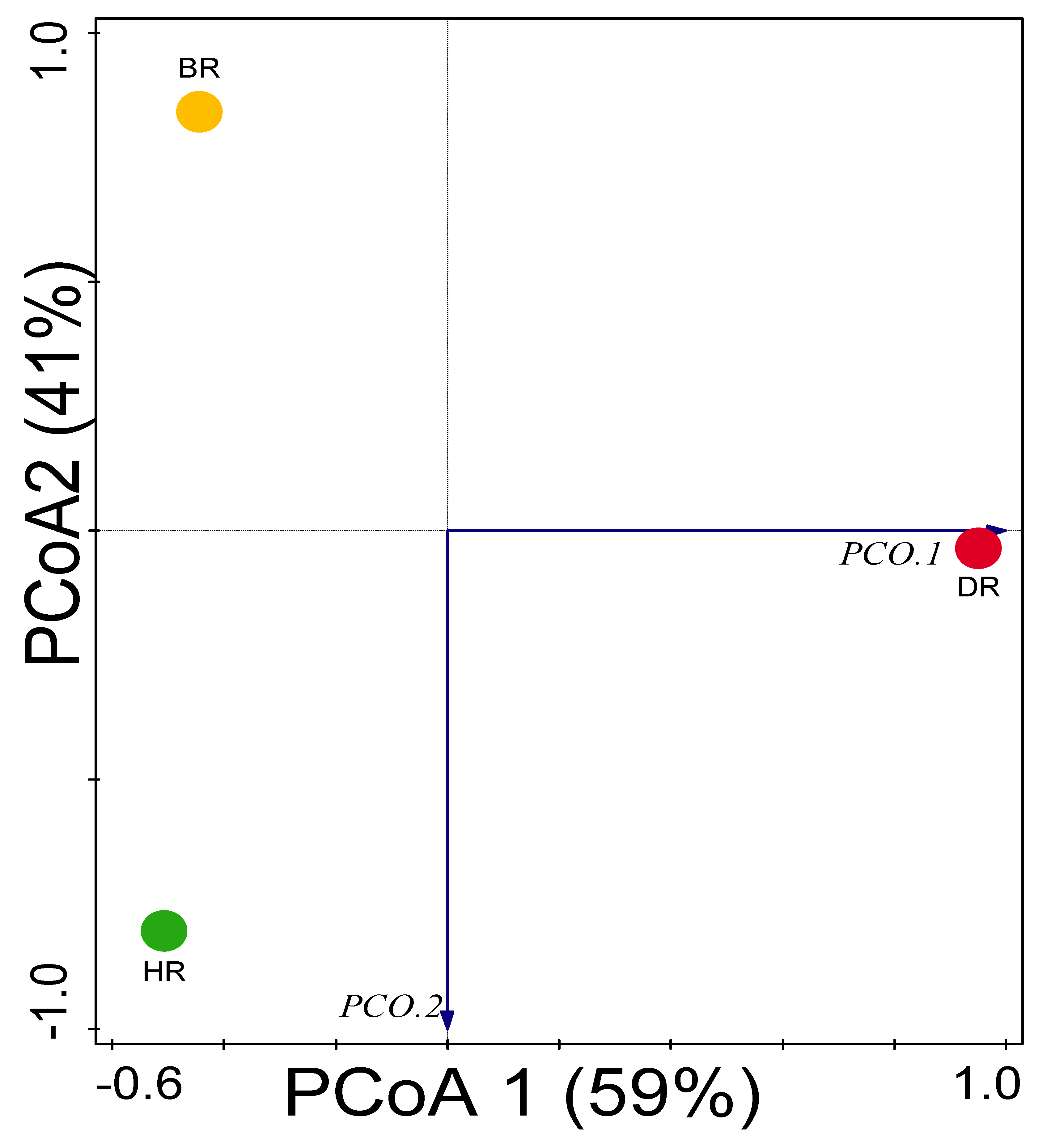
3.3. Alpha (α) and Beta (β) Diversity
3.4. Major Metabolic Pathways of Microbial Communities Associated with the Tomato Rhizosphere and Bulk Soils
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Zhu, A.; Tan, H.; Cao, L.; Zhang, R. Engineering Banana Endosphere Microbiome to improve Fusarium wilt resistance in Banana. Microbiome 2019, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Lin, X.; Tian, J.; Ji, L.; Chen, Y.; Tran, L.-S.P.; Tian, C. Research Advances of Beneficial Microbiota Associated with Crop Plants. Int. J. Mol. Sci. 2020, 21, 1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babalola, O.O.; Adedayo, A.A.; Fadiji, A.E. Metagenomic Survey of Tomato Rhizosphere Microbiome Using the Shotgun Approach. Microbiol. Resour. Announc. 2022, 11, e01131-21. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Sun, A.; Jiao, X.-Y.; Chen, Q.; Wu, A.-L.; Zheng, Y.; Lin, Y.-X.; He, J.-Z.; Hu, H.-W. Microbial communities in crop phyllosphere and root endosphere are more resistant than soil microbiota to fertilization. Soil Biol. Biochem. 2021, 153, 108113. [Google Scholar] [CrossRef]
- Vaishnav, A.; Shukla, A.K.; Sharma, A.; Kumar, R.; Choudhary, D.K. Endophytic Bacteria in Plant Salt Stress Tolerance: Current and Future Prospects. J. Plant Growth Regul. 2019, 38, 650–668. [Google Scholar] [CrossRef]
- de Vries Franciska, T.; Griffiths Rob, I.; Knight Christopher, G.; Nicolitch, O.; Williams, A. Harnessing rhizosphere microbiomes for drought-resilient crop production. Science 2020, 368, 270–274. [Google Scholar] [CrossRef]
- Porter, S.S.; Bantay, R.; Friel, C.A.; Garoutte, A.; Gdanetz, K.; Ibarreta, K.; Moore, B.M.; Shetty, P.; Siler, E.; Friesen, M.L. Beneficial microbes ameliorate abiotic and biotic sources of stress on plants. Funct. Ecol. 2020, 34, 2075–2086. [Google Scholar] [CrossRef] [Green Version]
- Lyu, D.; Backer, R.; Robinson, W.G.; Smith, D.L. Plant Growth-Promoting Rhizobacteria for Cannabis Production: Yield, Cannabinoid Profile and Disease Resistance. Front. Microbiol. 2019, 10, 1761. [Google Scholar] [CrossRef] [Green Version]
- Konappa, N.; Krishnamurthy, S.; Arakere, U.C.; Chowdappa, S.; Ramachandrappa, N.S. Efficacy of indigenous plant growth-promoting rhizobacteria and Trichoderma strains in eliciting resistance against bacterial wilt in a tomato. Egypt. J. Biol. Pest Control 2020, 30, 106. [Google Scholar] [CrossRef]
- Saia, S.; Aissa, E.; Luziatelli, F.; Ruzzi, M.; Colla, G.; Ficca, A.G.; Cardarelli, M.; Rouphael, Y. Growth-promoting bacteria and arbuscular mycorrhizal fungi differentially benefit tomato and corn depending upon the supplied form of phosphorus. Mycorrhiza 2020, 30, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Alori, E.T.; Babalola, O.O. Microbial Inoculants for Improving Crop Quality and Human Health in Africa. Front. Microbiol. 2018, 9, 2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dukare, A.; Paul, S. Biological control of Fusarium wilt and growth promotion in pigeon pea (Cajanus cajan) by antagonistic rhizobacteria, displaying multiple modes of pathogen inhibition. Rhizosphere 2021, 17, 100278. [Google Scholar] [CrossRef]
- Sahu, P.K.; Singh, D.P.; Prabha, R.; Meena, K.K.; Abhilash, P.C. Connecting microbial capabilities with the soil and plant health: Options for agricultural sustainability. Ecol. Indic. 2019, 105, 601–612. [Google Scholar] [CrossRef]
- Fones, H.N.; Bebber, D.P.; Chaloner, T.M.; Kay, W.T.; Steinberg, G.; Gurr, S.J. Threats to global food security from emerging fungal and oomycete crop pathogens. Nat. Food 2020, 1, 332–342. [Google Scholar] [CrossRef]
- Wei, Z.; Gu, Y.; Friman, V.-P.; Kowalchuk, G.A.; Xu, Y.; Shen, Q.; Jousset, A. Initial soil microbiome composition and functioning predetermine future plant health. Sci. Adv. 2019, 5, eaaw0759. [Google Scholar] [CrossRef] [Green Version]
- Soumare, A.; Boubekri, K.; Lyamlouli, K.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L. From Isolation of Phosphate Solubilizing Microbes to Their Formulation and Use as Biofertilizers: Status and Needs. Front. Bioeng. Biotechnol. 2020, 7, 425. [Google Scholar] [CrossRef]
- De Corato, U. Soil microbiota manipulation and its role in suppressing soil-borne plant pathogens in organic farming systems under the light of microbiome-assisted strategies. Chem. Biol. Technol. Agric. 2020, 7, 17. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O. The endosphere microbial communities, a great promise in agriculture. Int. Microbiol 2021, 24, 1–17. [Google Scholar] [CrossRef]
- Wei, X.; Wang, X.; Cao, P.; Gao, Z.; Chen, A.J.; Han, J. Microbial Community Changes in the Rhizosphere Soil of Healthy and Rusty Panax ginseng and Discovery of Pivotal Fungal Genera Associated with Rusty Roots. BioMed Res. Int. 2020, 2020, 8018525. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Feng, H.; Zhang, D.; Feng, Z.; Zhao, L.; Zhang, Y.; Deakin, G.; Peng, J.; Zhu, H.; Xu, X. Composition of Rhizosphere Microbial Communities Associated With Healthy and Verticillium Wilt Diseased Cotton Plants. Front. Microbiol. 2021, 12, 618169. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, J.-T.; Wang, W.-H.; Tsui, C.K.; Cai, L. Changes in Bacterial and Fungal Microbiomes Associated with Tomatoes of Healthy and Infected by Fusarium oxysporum f. sp. lycopersici. Microb. Ecol. 2021, 81, 1004–1017. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.-J.; Wang, L.-L.; Li, Q.; Shang, Q.-M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef] [PubMed]
- Ajilogba, C.F.; Babalola, O.O. Integrated Management Strategies for Tomato Fusarium Wilt. Biocontrol Sci. 2013, 18, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajilogba, C.F.; Babalola, O.O.; Ahmad, F. Antagonistic Effects of Bacillus species in Biocontrol of Tomato Fusarium Wilt. Stud. Ethno-Med. 2013, 7, 205–216. [Google Scholar] [CrossRef]
- Ahmad, F.; Babalola, O.O.; Tak, H.I. Potential of MALDI-TOF mass spectrometry as a rapid detection technique in plant pathology: Identification of plant-associated microorganisms. Anal. Bioanal. Chem 2012, 404, 1247–1255. [Google Scholar] [CrossRef]
- Akhtar, K.P.; Ullah, N.; Saleem, M.Y.; Iqbal, Q.; Asghar, M.; Khan, A.R. Evaluation of tomato genotypes for early blight disease resistance caused by Alternaria solani in Pakistan. Plant Pathol. 2019, 101, 1159–1170. [Google Scholar] [CrossRef]
- Mazumdar, P.; Singh, P.; Kethiravan, D.; Ramathani, I.; Ramakrishnan, N. Late blight in tomato: Insights into the pathogenesis of the aggressive pathogen Phytophthora infestans and future research priorities. Planta 2021, 253, 119. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, A.; Zhou, J.; Zhang, W.; Li, P. Comparison of bacterial communities in soil samples with and without tomato bacterial wilt caused by Ralstonia solanacearum species complex. BMC Microbiol. 2020, 20, 89. [Google Scholar] [CrossRef]
- Sun, G.; Feng, C.; Guo, J.; Zhang, A.; Xu, Y.; Wang, Y.; Day, B.; Ma, Q. The tomato Arp2/3 complex is required for resistance to the powdery mildew fungus Oidium neolycopersici. Plant Cell Environ. 2019, 42, 2664–2680. [Google Scholar] [CrossRef]
- Khalil Bagy, H.M.M.; Abo-Elyousr, K.A.M. Antibacterial activity of some essential oils on bacterial spot disease of tomato plant caused by Xanthomonas axonopodis pv. vesicatoria. Int. J. Plant Pathol. 2019, 8, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.-W.; Chun, S.C.; Chandrasekaran, M. Preparation and In Vitro Characterization of Chitosan Nanoparticles and Their Broad-Spectrum Antifungal Action Compared to Antibacterial Activities against Phytopathogens of Tomato. Agronomy 2019, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Safari, Z.S.; Ding, P.; Nakasha, J.J.; Yusoff, S.F. Controlling Fusarium oxysporum Tomato Fruit Rot under Tropical Condition Using Both Chitosan and Vanillin. Coatings 2021, 11, 367. [Google Scholar] [CrossRef]
- Istifadah, N.; Firman, A.R.; Desiana, M.F. Effectiveness of compost and microbial-enriched compost to suppress powdery mildew and early blight diseases in tomato. J. Anim. Plant Sci. 2020, 30, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Jahnová, J.; Činčalová, L.; Sedlářová, M.; Jedelská, T.; Sekaninová, J.; Mieslerová, B.; Luhová, L.; Barroso, J.B.; Petřivalský, M. Differential modulation of S-nitrosoglutathione reductase and reactive nitrogen species in wild and cultivated tomato genotypes during development and powdery mildew infection. Plant Physiol. Biochem. 2020, 155, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Fadiji, A.E.; Babalola, O.O. Metagenomics methods for the study of plant-associated microbial communities: A review. J. Microbiol. Methods 2020, 170, 105860. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Kim, Y.; Kim, J.M.; Chu, B.; Joa, J.-H.; Sang, M.K.; Song, J.; Weon, H.-Y. A preliminary examination of bacterial, archaeal, and fungal communities inhabiting different rhizocompartments of tomato plants under real-world environments. Sci. Rep. 2019, 9, 9300. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Xu, X.; Huo, Y.; Xiao, Y. Trichoderma-Inoculation and Mowing Synergistically Altered Soil Available Nutrients, Rhizosphere Chemical Compounds and Soil Microbial Community, Potentially Driving Alfalfa Growth. Front. Microbiol. 2019, 9, 3241. [Google Scholar] [CrossRef]
- Liu, L.; Huang, X.; Zhang, J.; Cai, Z.; Jiang, K.; Chang, Y. Deciphering the relative importance of soil and plant traits on the development of rhizosphere microbial communities. Soil Biol. Biochem. 2020, 148, 107909. [Google Scholar] [CrossRef]
- López, S.M.Y.; Pastorino, G.N.; Fernández-González, A.J.; Franco, M.E.E.; Fernández-López, M.; Balatti, P.A. The endosphere bacteriome of diseased and healthy tomato plants. Arch. Microbiol. 2020, 202, 2629–2642. [Google Scholar] [CrossRef]
- Zheng, X.; Liu, B.; Zhu, Y.; Wang, J.; Zhang, H.; Wang, Z. Bacterial community diversity associated with the severity of bacterial wilt disease in tomato fields in Southeast China. Can. J. Microbiol. 2019, 65, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-M.; Kong, H.G.; Song, G.C.; Ryu, C.-M. Disruption of Firmicutes and Actinobacteria abundance in tomato rhizosphere causes the incidence of bacterial wilt disease. ISME J. 2021, 15, 330–347. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xiong, W.; Zhang, R.; Hang, X.; Wang, D.; Li, R.; Shen, Q. Continuous application of different organic additives can suppress tomato disease by inducing the healthy rhizospheric microbiota through alterations to the bulk soil microflora. Plant Soil 2018, 423, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Zhang, L.; Nie, D.; Kuramae, E.E.; Shen, B.; Shen, Q. Bacterial Tomato Pathogen Ralstonia solanacearum Invasion Modulates Rhizosphere Compounds and Facilitates the Cascade Effect of Fungal Pathogen Fusarium solani. Microorganisms 2020, 8, 806. [Google Scholar] [CrossRef]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Kent, W.J. BLAT—The BLAST-Like Alignment Tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Wilke, A.; Harrison, T.; Wilkening, J.; Field, D.; Glass, E.M.; Kyrpides, N.; Mavrommatis, K.; Meyer, F. The M5nr: A novel non-redundant database containing protein sequences and annotations from multiple sources and associated tools. BMC Bioinform. 2012, 13, 141. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Rupek, P.; Richter, D.C.; Urich, T.; Gilbert, J.A.; Meyer, F.; Wilke, A.; Huson, D.H. Functional analysis of metagenomes and metatranscriptomes using SEED and KEGG. BMC Bioinform. 2011, 12, S21. [Google Scholar] [CrossRef] [Green Version]
- Khomtchouk, B.B.; Hennessy, J.R.; Wahlestedt, C. Shinyheatmap: Ultra fast low memory heatmap web interface for big data genomics. PLoS ONE 2017, 12, e0176334. [Google Scholar] [CrossRef] [Green Version]
- Hamel, C.; Gan, Y.; Sokolski, S.; Bainard, L.D. High frequency cropping of pulses modifies soil nitrogen level and the rhizosphere bacterial microbiome in 4-year rotation systems of the semiarid prairie. Agric. Ecosyst. Environ. Appl. Soil Ecol. 2018, 126, 47–56. [Google Scholar] [CrossRef]
- Carrell, A.; Frank, C. Bacterial endophyte communities in the foliage of coast redwood and giant sequoia. Front. Microbiol. 2015, 6, 1008. [Google Scholar] [CrossRef] [PubMed]
- Raimi, A.; Adeleke, R.; Roopnarain, A. Soil fertility challenges and Biofertiliser as a viable alternative for increasing smallholder farmer crop productivity in sub-Saharan Africa. Cogent Food Agric. 2017, 3, 1400933. [Google Scholar] [CrossRef]
- Rolfe, S.A.; Griffiths, J.; Ton, J. Crying out for help with root exudates: Adaptive mechanisms by which stressed plants assemble health-promoting soil microbiomes. Curr. Opin. Microbiol. 2019, 49, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Li, L.; Wang, X.; You, J.; Li, J.; Chen, X. Elevational is the main factor controlling the soil microbial community structure in alpine tundra of the Changbai Mountain. Sci. Rep. 2020, 10, 12442. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Kanu, J.O.; Babalola, O.O. Metagenomic profiling of rhizosphere microbial community structure and diversity associated with maize plant as affected by cropping systems. Int. Microbiol. 2021, 24, 325–335. [Google Scholar] [CrossRef]
- Nessner Kavamura, V.; Taketani, R.G.; Lançoni, M.D.; Andreote, F.D.; Mendes, R.; Soares De Melo, I. Water Regime Influences Bulk Soil and Rhizosphere of Cereus jamacaru Bacterial Communities in the Brazilian Caatinga Biome. PLoS ONE 2013, 8, e73606. [Google Scholar] [CrossRef]
- Pang, Z.; Dong, F.; Liu, Q.; Lin, W.; Hu, C.; Yuan, Z. Soil Metagenomics Reveals Effects of Continuous Sugarcane Cropping on the Structure and Functional Pathway of Rhizospheric Microbial Community. Front. Microbiol. 2021, 12, 369. [Google Scholar] [CrossRef]
- Jung, J.; Kim, J.-S.; Taffner, J.; Berg, G.; Ryu, C.-M. Archaea, tiny helpers of land plants. Comput. Struct. Biotechnol. J. 2020, 18, 2494–2500. [Google Scholar] [CrossRef]
- Hussain, H.I.; Kasinadhuni, N.; Arioli, T. The effect of seaweed extract on tomato plant growth, productivity and soil. J. Appl. Phycol. 2021, 33, 1305–1314. [Google Scholar] [CrossRef]
- Wu, A.-L.; Jiao, X.-Y.; Fan, F.-F.; Wang, J.-S.; Guo, J.; Dong, E.-W.; Wang, L.-G.; Shen, X.-M. Effect of continuous sorghum cropping on the rhizosphere microbial community and the role of Bacillus amyloliquefaciens in altering the microbial composition. Plant Growth Regul. 2019, 89, 299–308. [Google Scholar] [CrossRef]
- Enebe, M.C.; Babalola, O.O. Effects of inorganic and organic treatments on the microbial community of maize rhizosphere by a shotgun metagenomics approach. Ann. Microbiol. 2020, 70, 49. [Google Scholar] [CrossRef]
- Dong, C.; Wang, L.; Li, Q.; Shang, Q. Epiphytic and Endophytic Fungal Communities of Tomato Plants. Hortic. Plant J. 2021, 7, 38–48. [Google Scholar] [CrossRef]
- Omomowo, I.O.; Adedayo, A.A.; Omomowo, O.I. Biocontrol potential of rhizospheric fungi from Moringa oleifera, their phytochemicals and secondary metabolite assessment against spoilage fungi of sweet orange (Citrus sinensis). Asian J. Appl. Sci. 2020, 8, 1–14. [Google Scholar] [CrossRef]
- Rampersad, S. Pathogenomics and Management of Fusarium Diseases in Plants. Pathogens 2020, 9, 340. [Google Scholar] [CrossRef] [PubMed]
- Ayangbenro, A.S.; Babalola, O.O. Reclamation of arid and semi-arid soils: The role of plant growth-promoting archaea and bacteria. Curr. Plant Biol. 2021, 25, 100173. [Google Scholar] [CrossRef]
- Schmidt, J.E.; Kent, A.D.; Brisson, V.L.; Gaudin, A.C.M. Agricultural management and plant selection interactively affect rhizosphere microbial community structure and nitrogen cycling. Microbiome 2019, 7, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbasi, S.; Sadeghi, A.; Omidvari, M.; Tahan, V. The stimulators and responsive genes to induce systemic resistance against pathogens: An exclusive focus on tomato as a model plant. Biocatal. Agric. Biotechnol. 2021, 33, 101993. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; Khedr, A.A.; Mehiar, F.; El-Kady, E.M.; Baazeem, A.; Shimizu, M. Suppression of Pseudomonas syringae pv. tomato infection by rhizosphere fungi. Pest Manag. Sci. 2021, 77, 4350–4356. [Google Scholar] [CrossRef]
- Kong, H.G.; Song, G.C.; Sim, H.-J.; Ryu, C.-M. Achieving similar root microbiota composition in neighbouring plants through airborne signalling. ISME J. 2021, 15, 397–408. [Google Scholar] [CrossRef]
- Nwachukwu, B.C.; Babalola, O.O. Perspectives for sustainable agriculture from the microbiome in plant rhizosphere. Plant Biotechnol. Rep. 2021, 15, 259–278. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, Z.; Zhu, Y.; Wang, J.; Liu, B. Effects of a microbial restoration substrate on plant growth and rhizosphere bacterial community in a continuous tomato cropping greenhouse. Sci. Rep. 2020, 10, 13729. [Google Scholar] [CrossRef] [PubMed]
- Taffner, J.; Cernava, T.; Erlacher, A.; Berg, G. Novel insights into plant-associated archaea and their functioning in arugula (Eruca sativa Mill.). J. Adv. Res. 2019, 19, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Mayak, S.; Tirosh, T.; Glick, B.R. Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol. Biochem. 2004, 42, 565–572. [Google Scholar] [CrossRef]
- Goswami, L.; Nath, A.; Sutradhar, S.; Bhattacharya, S.S.; Kalamdhad, A.; Vellingiri, K.; Kim, K.-H. Application of drum compost and vermicompost to improve soil health, growth, and yield parameters for tomato and cabbage plants. J. Environ. Manag. 2017, 200, 243–252. [Google Scholar] [CrossRef]
- Bielsa, B.; Hewitt, S.; Reyes-Chin-Wo, S.; Dhingra, A.; Rubio-Cabetas, M.J. Identification of water use efficiency related genes in ‘Garnem’ almond-peach rootstock using time-course transcriptome analysis. PLoS ONE 2018, 13, e0205493. [Google Scholar] [CrossRef]
- Dawood, M.F.A.; Tahjib-Ul-Arif, M.; Sohag, A.A.M.; Abdel Latef, A.A.H.; Ragaey, M.M. Mechanistic Insight of Allantoin in Protecting Tomato Plants Against Ultraviolet C Stress. Plants 2020, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.W.; Kuramae, E.E.; Navarrete, A.A.; van Veen, J.A.; Tsai, S.M. Taxonomical and functional microbial community selection in soybean rhizosphere. ISME J. 2014, 8, 1577–1587. [Google Scholar] [CrossRef] [PubMed]
- Santana, M.M.; Dias, T.; Gonzalez, J.M.; Cruz, C. Transformation of organic and inorganic sulfur– adding perspectives to new players in soil and rhizosphere. Soil Biol. Biochem. 2021, 160, 108306. [Google Scholar] [CrossRef]
- Bodale, I.; Mihalache, G.; Achiţei, V.; Teliban, G.-C.; Cazacu, A.; Stoleru, V. Evaluation of the Nutrients Uptake by Tomato Plants in Different Phenological Stages Using an Electrical Conductivity Technique. Agriculture 2021, 11, 292. [Google Scholar] [CrossRef]
- Stigter, K.A.; Plaxton, W.C. Molecular Mechanisms of Phosphorus Metabolism and Transport during Leaf Senescence. Plants 2015, 4, 773–798. [Google Scholar] [CrossRef] [Green Version]
- Akinola, S.; Ayangbenro, A.; Babalola, O. Metagenomic Insight into the Community Structure of Maize-Rhizosphere Bacteria as Predicted by Different Environmental Factors and Their Functioning within Plant Proximity. Microorganisms 2021, 9, 1419. [Google Scholar] [CrossRef] [PubMed]
- Emenecker, R.J.; Strader, L.C. Auxin-Abscisic Acid Interactions in Plant Growth and Development. Biomolecules 2020, 10, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Statistics | HR | DR | BR | p-Value |
|---|---|---|---|---|---|
| Phylum | Simpson_1-D | 0.7083 ± 0.07 | 0.6801 ± 0.06 | 0.6735 ± 0.06 | 0.94 |
| Shannon_H | 1.695 ± 0.24 | 1.598 ± 0.2 | 1.586 ± 0.23 | ||
| Evenness_e^H/S | 0.181 ± 0.09 | 0.183 ± 0.08 | 0.201 ± 0.09 | ||
| Genus | Simpson_1-D | 0.924 ± 0.04 | 0.9216 ± 0.04 | 0.924 ± 0.04 | 0.78 |
| Shannon_H | 2.745 ± 0.20 | 2.741 ± 0.23 | 2.739 ± 0.023 | ||
| Evenness_e^H/S | 0.5334 ± 0.10 | 0.5365 ± 0.10 | 0.5347 ± 0.10 |
| SEED Subsystem Level 3 | HR (%) | DR (%) | BR(%) | p-Value |
|---|---|---|---|---|
| Carbohydrate metabolism | ||||
| Trehalose Biosynthesis | 0.6378 ± 0.01 | 0.5787 ± 0.01 | 0.5744 ± 0.01 | 0.06 |
| TCA Cycle | 0.5744 ± 0.001 | 0.5648 ± 0.001 | 0.5577 ± 0.01 | 0.05 |
| Glycolysis and Gluconeogenesis | 0.5056 ± 0.01 | 0.4250 ± 0.01 | 0.4900 ± 0.01 | 0.05 |
| Glycogen metabolism | 0.3266 ± 0.002 | 0.3227 ± 0.01 | 0.2801 ± 0.01 | 0.06 |
| Glycerol and Glycerol-3-phosphate Uptake and Utilization | 0.2517 ± 0.00 | 0.2348 ± 0.003 | 0.2298 ± 0.003 | 0.05 |
| Glycolysis and Gluconeogenesis, including Archaeal enzymes | 0.2373 ± 0.004 | 0.2227 ± 0.005 | 0.2327 ± 0.003 | 0.25 |
| Nitrogen metabolism | ||||
| Allantoin Utilization | 0.0923 ± 0.03 | 0.0708 ± 0.02 | 0.0882 ± 0.00 | 0.05 |
| Ammonia assimilation | 0.5551 ± 0.003 | 0.5321 ± 0.002 | 0.5305 ± 0.004 | 0.07 |
| Nitrate and nitrite ammonification | 0.4344 ± 0.005 | 0.3446 ± 0.004 | 0.3403 ± 0.000 | 0.06 |
| Nitric oxide synthase | 0.1810 ± 0.004 | 0.1432 ± 0.0.004 | 0.1625 ± 0.004 | 0.03 |
| Nitrogen fixation | 0.0555 ± 0.004 | 0.0518 ± 0.000 | 0.0350 ± 0.003 | 0.06 |
| Denitrification | 0.0754 ± 0.002 | 0.0452 ± 0.002 | 0.0434 ± 0.003 | 0.06 |
| Sulfur metabolism | ||||
| Inorganic Sulfur Assimilation | 0.3981 ± 0.001 | 0.3920 ± 0.004 | 0.3975 ± 0.04 | 0.39 |
| Sulfur oxidation | 0.1209 ± 0.001 | 0.1154 ± 0.001 | 0.1100 ± 0.001 | 0.03 |
| Alkanesulfonates Utilization | 0.0365 ± 0.001 | 0.0309 ± 0.002 | 0.0294 ± 0.001 | 0.11 |
| Alkanesulfonate assimilation | 0.1805 ± 0.001 | 0.1660 ± 0.006 | 0.1625 ± 0.003 | 0.06 |
| Phosphorus metabolism | ||||
| Phosphate metabolism | 0.8512 ± 0.003 | 0.8141 ± 0.006 | 0.8023 ± 0.005 | 0.04 |
| Pentose phosphate pathway | 0.4227 ± 0.01 | 0.3211 ± 0.01 | 0.4085 ± 0.01 | 0.05 |
| P-uptake (cyanobacteria) | 0.1184 ± 0.004 | 0.1135 ± 0.001 | 0.1088 ± 0.002 | 0.11 |
| Alkylphosphonate utilization | 0.1030 ± 0.003 | 0.0915 ± 0.002 | 0.0850 ± 0.002 | 0.04 |
| Phosphonate metabolism | 0.0116 ± 0.000 | 0.0119 ± 0.001 | 0.0103 ± 0.001 | 0.25 |
| Phosphoenolpyruvate phosphomutase | 0.0094 ± 0.001 | 0.0091 ± 0.001 | 0.0086 ± 0.001 | 0.56 |
| Secondary metabolism | ||||
| Auxin biosynthesis | 0.0989 ± 0.002 | 0.0952 ± 0.000 | 0.0921 ± 0.001 | 0.06 |
| Phytoalexin biosynthesis | 0.0066 ± 0.000 | 0.0034 ± 0.000 | 0.0062 ± 0.000 | 0.07 |
| Alkaloid biosynthesis from L-lysine | 0.0366 ± 0.004 | 0.0357 ± 0.001 | 0.0299 ± 0.002 | 0.07 |
| Tannin biosynthesis | 0.0068 ± 0.000 | 0.0067 ± 0.001 | 0.0049 ± 0.000 | 0.06 |
| Heme and Siroheme Biosynthesis | 0.4208 ± 0.01 | 0.3947 ± 0.01 | 03775 ± 0.01 | 0.04 |
| Pyridoxin (Vitamin B6) Biosynthesis | 0.3935 ± 0.004 | 0.3815 ± 0.003 | 0.3791 ± 0.003 | 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adedayo, A.A.; Fadiji, A.E.; Babalola, O.O. The Effects of Plant Health Status on the Community Structure and Metabolic Pathways of Rhizosphere Microbial Communities Associated with Solanum lycopersicum. Horticulturae 2022, 8, 404. https://doi.org/10.3390/horticulturae8050404
Adedayo AA, Fadiji AE, Babalola OO. The Effects of Plant Health Status on the Community Structure and Metabolic Pathways of Rhizosphere Microbial Communities Associated with Solanum lycopersicum. Horticulturae. 2022; 8(5):404. https://doi.org/10.3390/horticulturae8050404
Chicago/Turabian StyleAdedayo, Afeez Adesina, Ayomide Emmanuel Fadiji, and Olubukola Oluranti Babalola. 2022. "The Effects of Plant Health Status on the Community Structure and Metabolic Pathways of Rhizosphere Microbial Communities Associated with Solanum lycopersicum" Horticulturae 8, no. 5: 404. https://doi.org/10.3390/horticulturae8050404
APA StyleAdedayo, A. A., Fadiji, A. E., & Babalola, O. O. (2022). The Effects of Plant Health Status on the Community Structure and Metabolic Pathways of Rhizosphere Microbial Communities Associated with Solanum lycopersicum. Horticulturae, 8(5), 404. https://doi.org/10.3390/horticulturae8050404