
Prediction of the Potential Distribution of Vaccinium uliginosum in China Based on the Maxent Niche Model

Abstract
1. Introduction
2. Materials and Methods
2.1. Species Distribution
2.2. Environmental Data
2.3. Model Building and Data Processing
3. Results
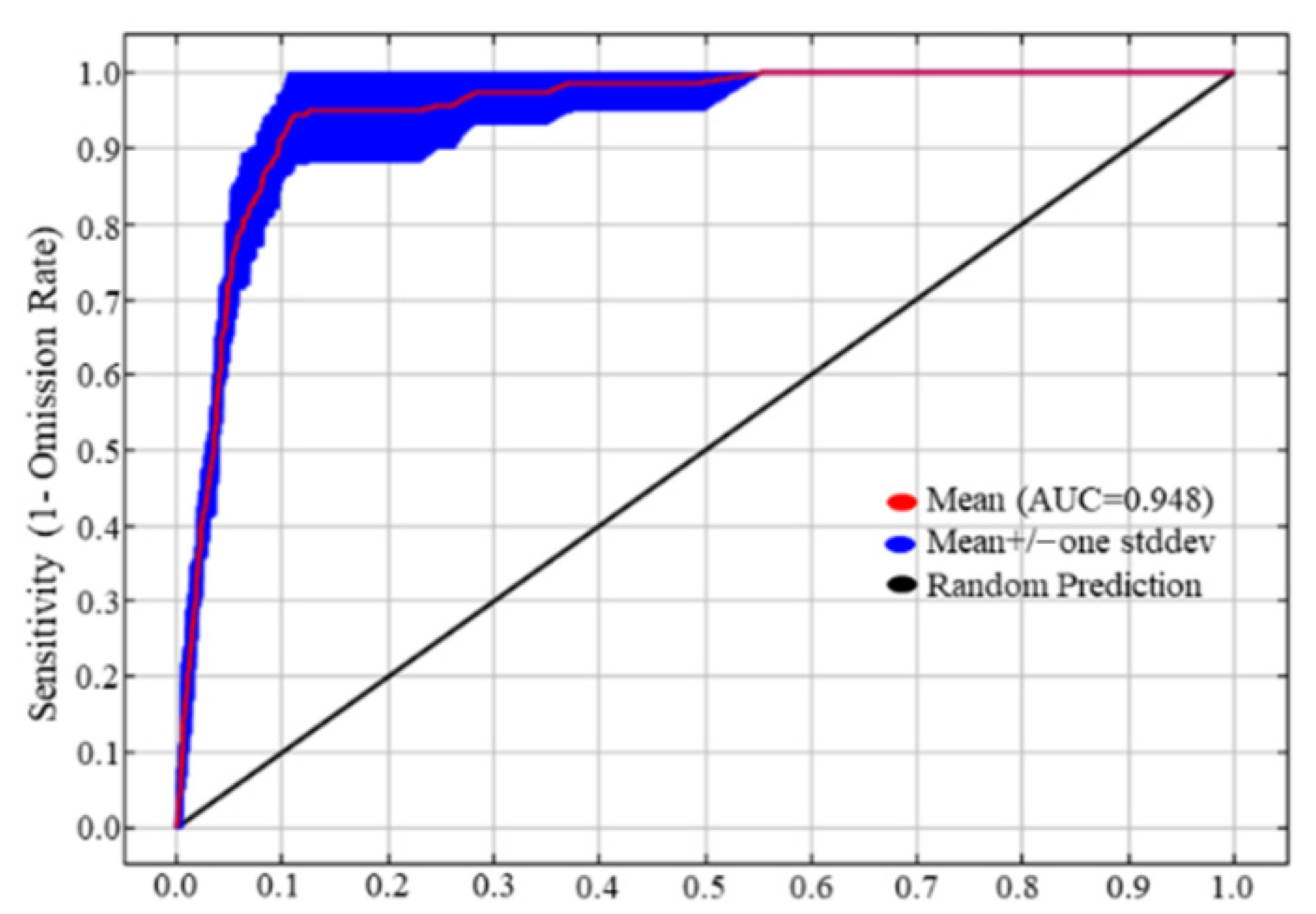
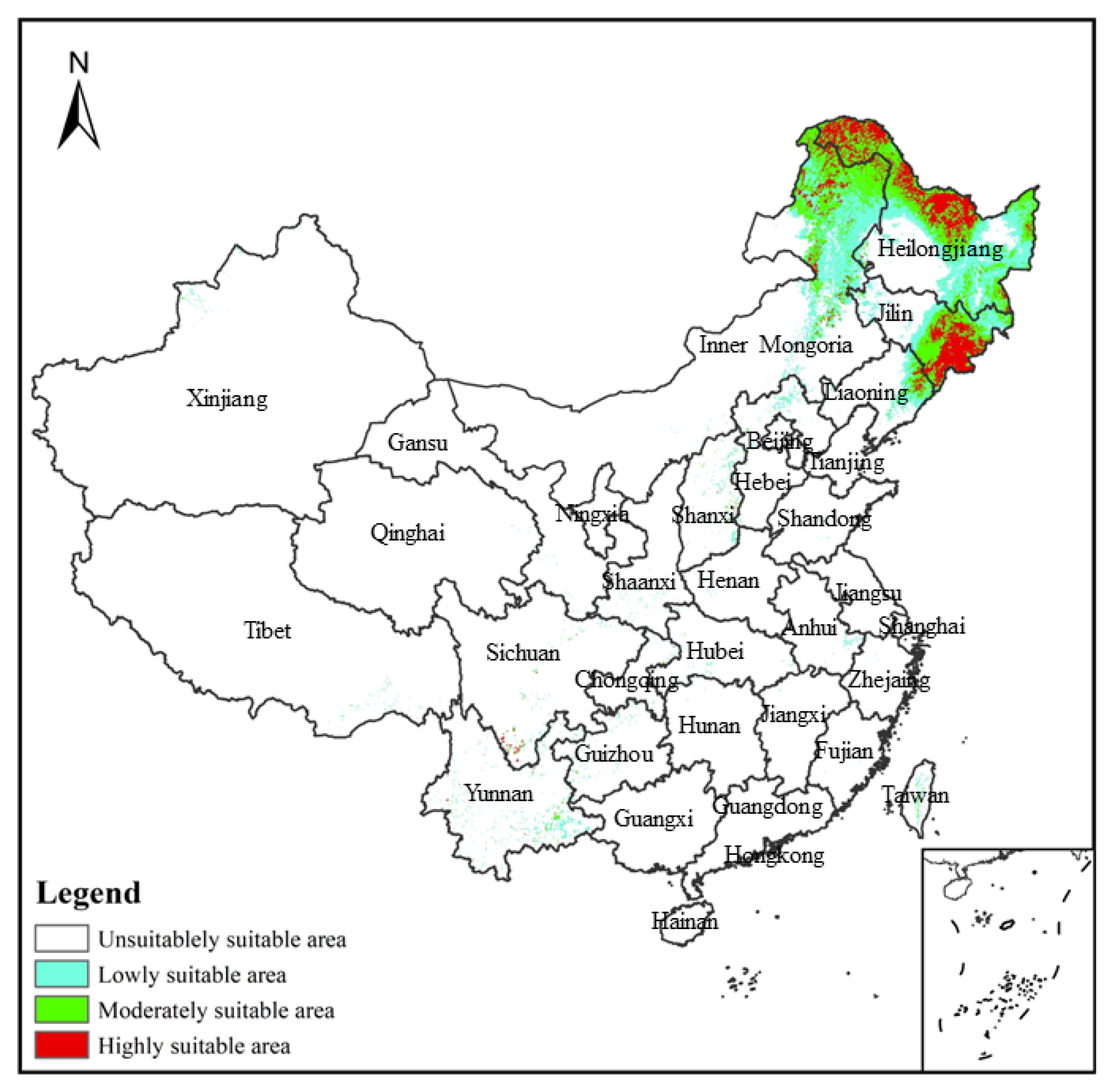
3.1. Model Prediction Accuracy Test and Its Prediction of the Potential Distribution Region of V. uliginosum in the Contemporary Region
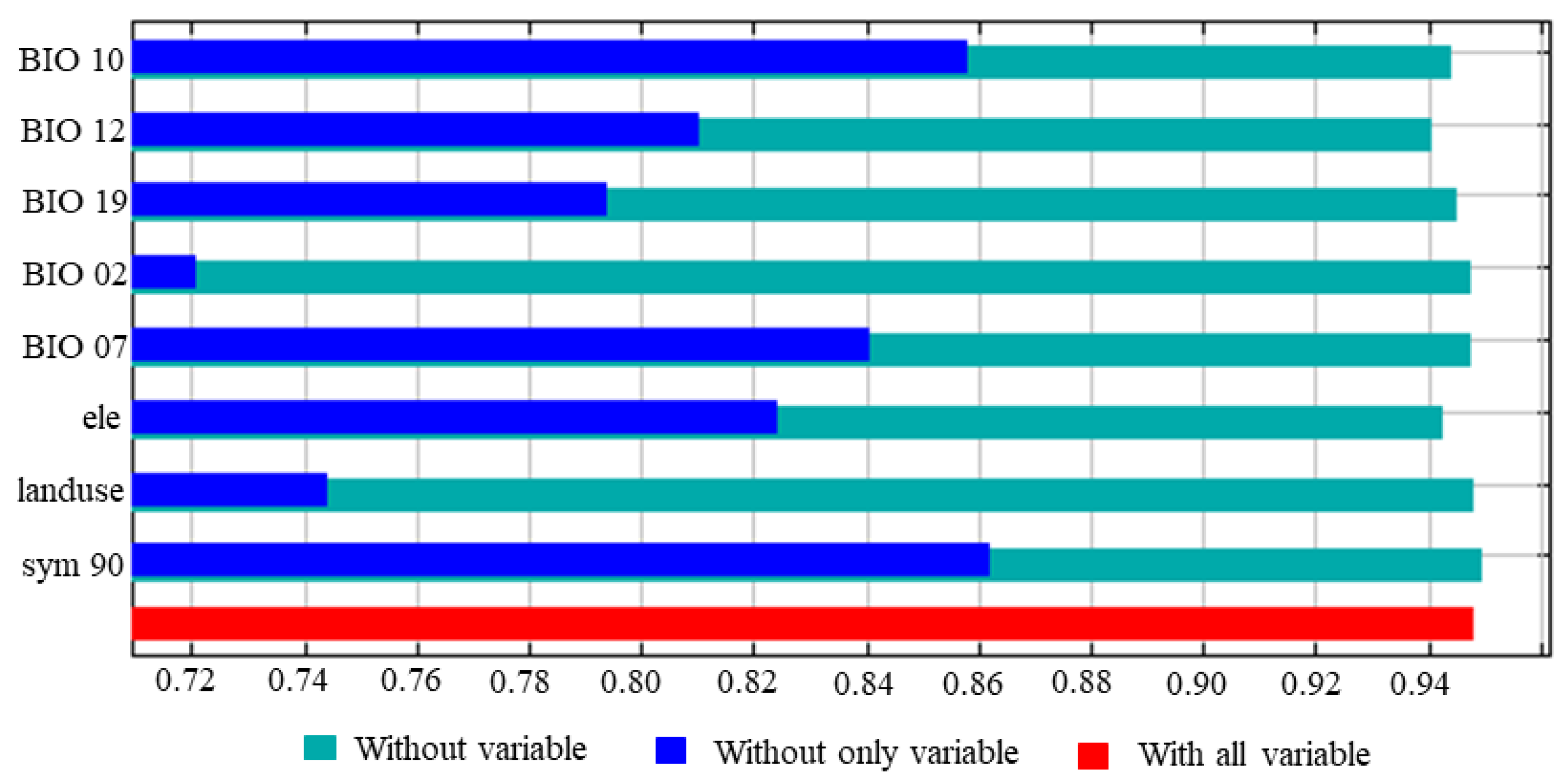
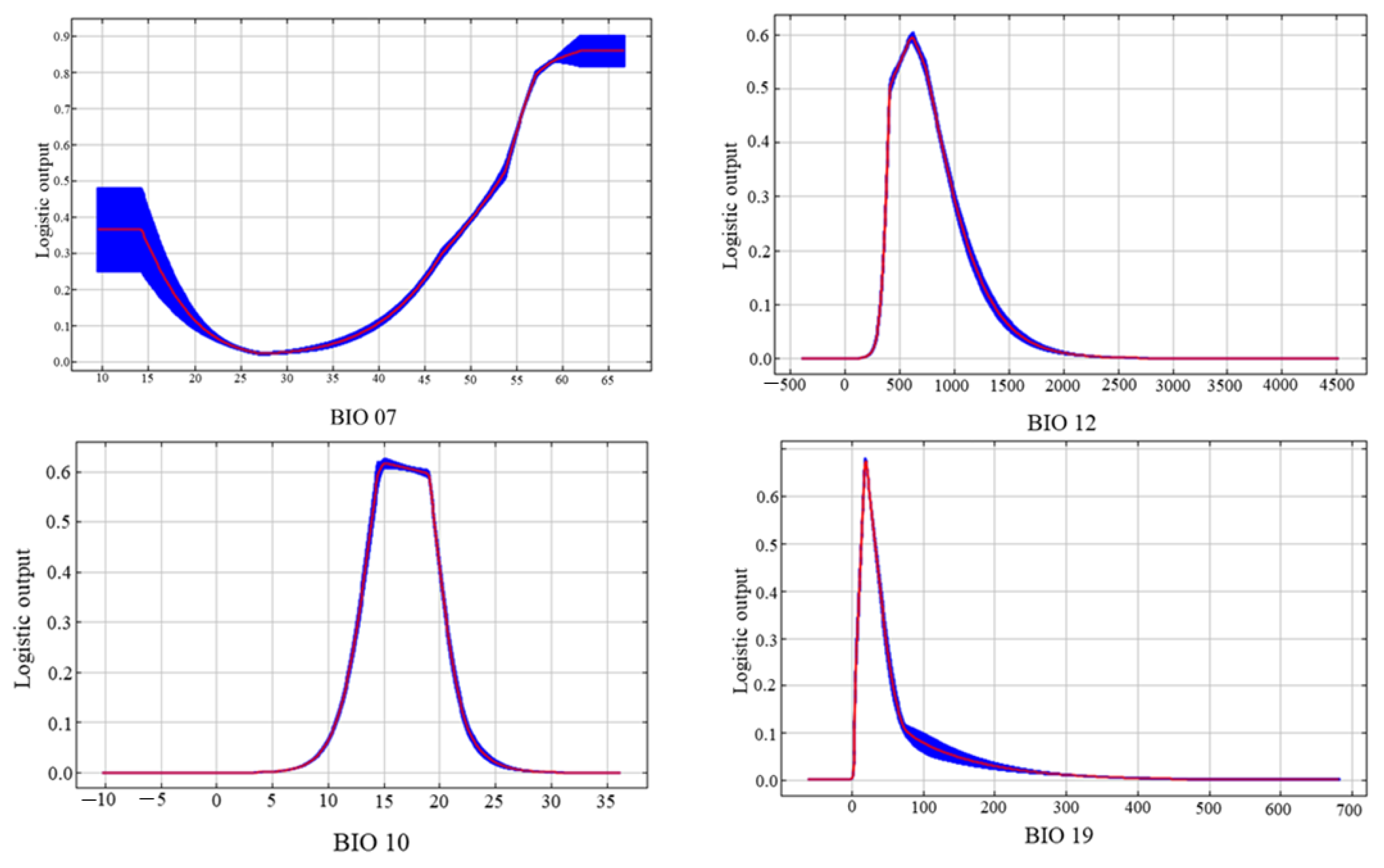
3.2. Analysis of the Influence of Environmental Variables on the Prediction of V. uliginosum Distribution
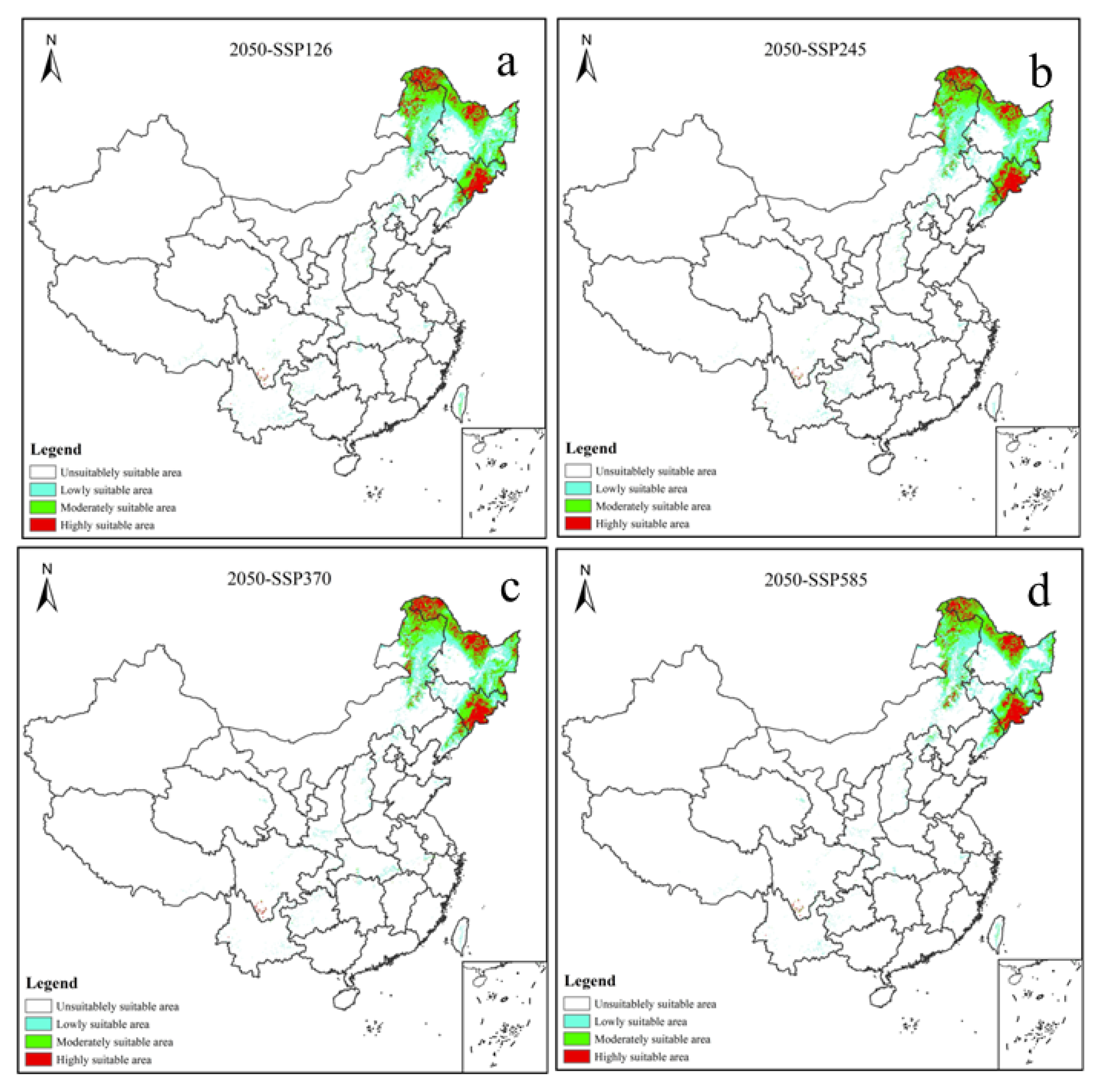
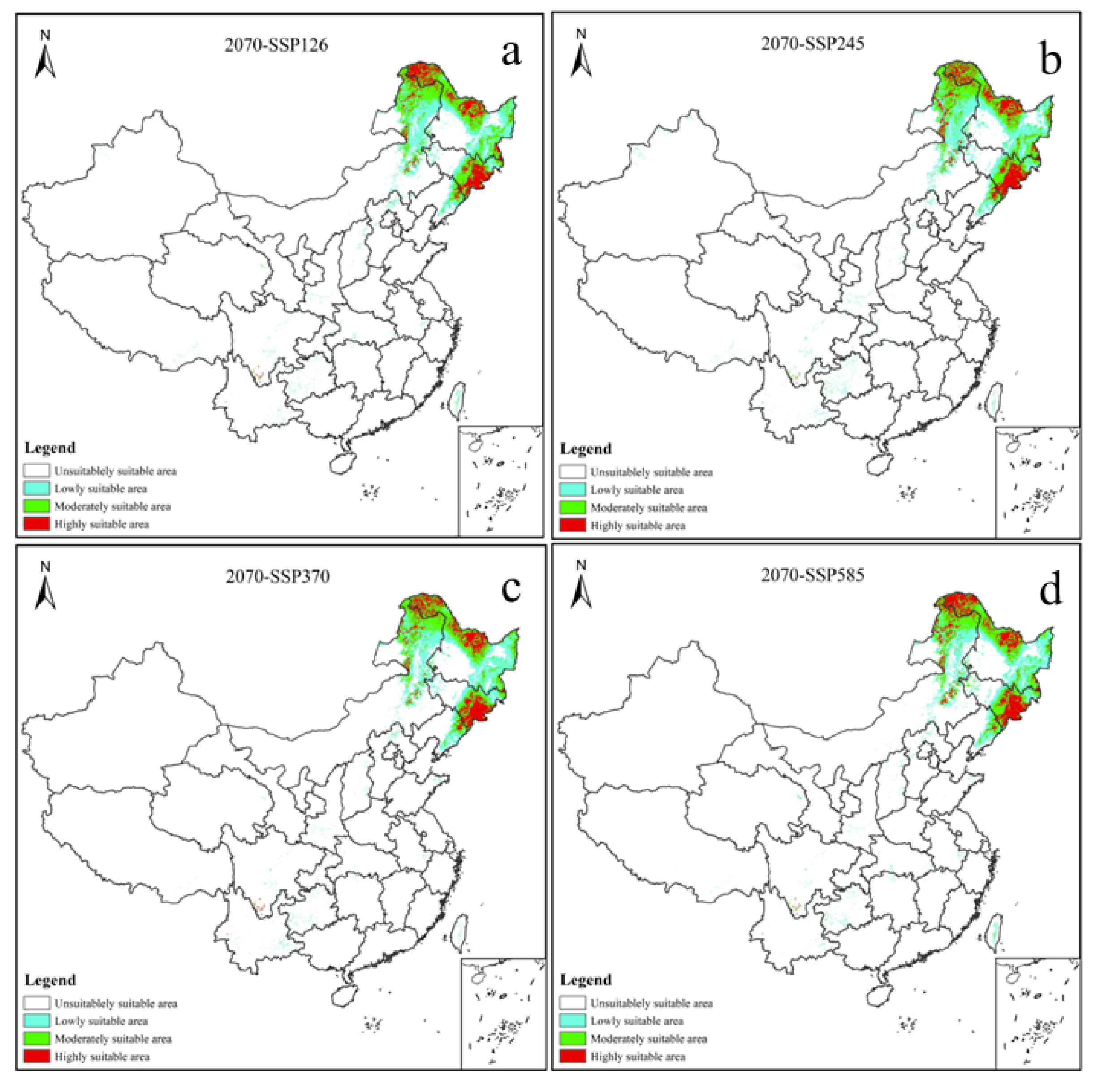
3.3. Potential Distribution of V. uliginosum under Future Climatic Scenario
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kozak, K.H.; Graham, C.H.; Wiens, J.J. Integrating GIS-based environmental data into evolutionary biology. Trends Ecol. Evol. 2008, 23, 141–148. [Google Scholar] [CrossRef]
- Chen, J.; Brissette, F.P.; Lucas-Picher, P. Assessing the limits of bias-correcting climate model outputs for climate change impact studies. J. Geophys. Res. Atmos. 2015, 120, 1123–1136. [Google Scholar] [CrossRef]
- Jewitt, D.; Erasmus, B.F.N.; Goodman, P.S.; O’Connor, T.G.; Hargrove, W.W.; Maddalena, D.M.; Witkowski, E.T.F. Climate-induced change of environmentally defined floristic domains: A conservation-based vulnerability framework. Appl. Geogr. 2015, 63, 33–42. [Google Scholar] [CrossRef]
- Mantyka-Pringle, C.S.; Visconti, P.; Di Marco, M.; Martin, T.G.; Rondinini, C.; Rhodes, J.R. Climate change modifies risk of global biodiversity loss due to land-cover change. Biol. Conserv. 2015, 187, 103–111. [Google Scholar] [CrossRef]
- Trumbore, S.; Brando, P.; Hartmann, H. Forest health and global change. Science 2015, 349, 814–818. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.E.; Stouffer, R.J.; Meehl, G.A. An overview of CMIP5 and the experiment design. Am. Meteorol. Soc. 2012, 93, 485–498. [Google Scholar] [CrossRef]
- Thuiller, W.; Vayreda, J.; Pino, J. Large-scale environmental correlates of forest tree distributions in Catalonia (NE Spain). Glob. Ecol. Biogeogr. 2003, 12, 313–325. [Google Scholar] [CrossRef]
- Jeffree, E.P.; Jeffree, C.E. Temperature and the bio-geographical distribution of species. Funct. Ecol. 1994, 8, 640–650. [Google Scholar] [CrossRef]
- Ohsawa, M. The montane cloud forest and its gradational changes in southeast Asia. In Tropical Montane Cloud Forests; Springer: New York, NY, USA, 1993; pp. 163–170. [Google Scholar]
- Chen, I.C.; Hill, J.K.; Ohlemuller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent Plant Diversity Changes on Europe’s Mountain Summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef]
- CMA Climate Change Centre. Blue Book on Climate Change in China (2022); Science Press: Beijing, China, 2022. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Duan, Y.Z.; Wang, H.T.; Wang, C.; Du, Z.Y. Potential distribution of endangered plant Helianthemum songaricum in China under climate change. J. Plant. Resour. Environ. 2020, 29, 55–68. [Google Scholar]
- Hirzel, A.H.; Hausser, J.; Chessel, D.; Perrin, N. Ecological-niche factor analysis: How to compute habitat-suitability maps without absence data? Ecology 2002, 83, 2027–2036. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. BIOCLIM: The first species distribution modelling package, its early applications and relevance to most current MaxEnt studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Papes, M.; Gaubert, P. Modelling ecological niches from low number of occurrences: Assessment of the conservation status of poorly known viverrids (Mammalia, Carnivora) across two continents. Divers. Distrib. 2007, 13, 890–902. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P. Novel methods improve prediction of species’distributions from occurrence data. Ecography 2010, 29, 129–151. [Google Scholar] [CrossRef]
- Cabra-Rivas, I.; Saldaña, A.; Castro-Díez, P.; Gallien, L. A multi-scale approach to identify invasion drivers and invader future dynamics. Biol. Invasions 2016, 18, 411–426. [Google Scholar] [CrossRef]
- Liu, H.; Gong, H.; Qi, X.; Li, Y.; Lin, Z. Relative importance of environmental variables for the distribution of the invasive marsh species Spartina altenrniflora across different spatial scales. Mar. Freshw. Res. 2018, 69, 790–801. [Google Scholar] [CrossRef]
- Vu, T.T.; Tran, D.V.; Tran, H.T.P.; Nguyen, M.D.; Do, T.A.; Ta, N.T.; Cao, H.T.; Pham, N.T.; Phan, D.V. An assessment of the impact of climate change on the distribution of the grey-shanked douc Pygathrix cinerea using an ecological niche model. Primates 2020, 61, 267–275. [Google Scholar] [CrossRef]
- Pan, J.; Fan, X.; Luo, S.; Zhang, Y.; Yao, S.; Guo, Q.; Qian, Z. Predicting the Potential Distribution of Two Varieties of Litsea coreana (Leopard-Skin Camphor) in China under Climate Change. Forests 2020, 11, 1159. [Google Scholar] [CrossRef]
- Gao, Z.Y. Distribution of wild Dusky lingonberry resources in Wenshan region, Yunnan. Pract. Rural. Technol. 2010, 4, 39–41. [Google Scholar]
- Lu, L. Wild Dusky orange in the Daxingan Mountains. North. Fruits 2004, 5, 42. [Google Scholar] [CrossRef]
- Fang, J.Y.; Song, Y.C.; Liu, H.Y.; Piao, S.L. Vegetation-climate relationship and its application in the division of vegetation zone in China. Acta Bot. Sin. 2002, 44, 1105–1122. [Google Scholar]
- Chakraborty, A.; Joshi, P.K.; Sachdeva, K. Predicting distribution of major forest tree species to potential impacts of climate change in the central Himalayan region. Ecol. Eng. 2016, 97, 593–609. [Google Scholar] [CrossRef]
- Zuo, J.; Tang, X.; Zhang, H.; Zu, M.; Zhang, X.; Yuan, Y. Analysis of niche shift and potential suitable distributions of Dendrobium under the impact of global climate change. Environ. Sci. Pollut. Res. 2022. [Google Scholar] [CrossRef]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecograph 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Cobos MEPeterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef]
- Kong, F.; Tang, L.; He, H.; Yang, F.; Tao, J.; Wang, W. Assessing the impact of climate change on the distribution of Osmanthus fragrans using Maxent. Environ. Sci. Pollut. 2021, 28, 34655–34663. [Google Scholar] [CrossRef]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Wu, D.; Zhu, C. Prediction of the spatial distribution of Alternanthera philoxeroides in China based on ArcGIS and MaxEnt. Glob. Ecol. Conserv. 2020, 21, e00856. [Google Scholar] [CrossRef]
- Jackson, C.R.; Robertson, M.P. Predicting the potential distribution of an endangered cryptic subterranean mammal from few occurrence records. J. Nat. Conserv. 2011, 19, 87–94. [Google Scholar] [CrossRef]
- Clark, J.T.; Fei, S.; Liang, L.; Rieske, L.K. Mapping eastern hemlock: Comparing classification techniques to evaluate susceptibility of a fragmented and valued resource to an exotic invader, the hemlock woolly adelgid. For. Ecol. Manag. 2012, 266, 216–222. [Google Scholar] [CrossRef]
- Slater, H.; Michael, E. Predicting the Current and Future Potential Distributions of Lymphatic Filariasis in Africa Using Maximum Entropy Ecological Niche Modelling. PLoS ONE 2012, 7, e32202. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Yu, W.; Zhao, H.; Xian, X.; Jing, K.; Yang, N.; Lu, X.; Liu, W. Potential Global Distribution of Invasive Alien Species, Anthonomus grandis Boheman, under Current and Future Climate Using Optimal MaxEnt Model. Agriculture 2022, 12, 1759. [Google Scholar] [CrossRef]
- Gong, L.; Li, X.; Wu, S.; Jiang, L. Prediction of potential distribution of soybean in the frigid region in China with MaxEnt modeling. Ecol. Inform. 2022, 72, 101834. [Google Scholar] [CrossRef]
- Wang, J.; Qian, Q.; Zhang, F.; Jia, X.; He, J. The possible future changes in potential suitable habitats of Tetrastigma hemsleyanum (Vitaceae) in China predicted by an ensemble mode. Glob. Ecol. Conserv. 2022, 35, e02083. [Google Scholar] [CrossRef]
- Vega, R.; Fløjgaard, C.; Lira-Noriega, A.; Nakazawa, Y.; Svenning, J.C.; Searle, J.B. Northern glacial refugia for the pygmy shrew Sorex minutus in Europe revealed by phylogeographic analyses and species distribution modelling. Ecography 2010, 33, 260–271. [Google Scholar] [CrossRef]
- Kantar, M.B.; Sosa, C.C.; Khoury, C.K.; Castañeda-Álvarez, N.P.; Achicanoy, H.A.; Bernau, V.; Kane, N.C.; Marek, L.; Seiler, G.; Rieseberg, L.H. Ecogeography and utility to plant breeding of the crop wild relatives of sunflower (Helianthus annuus L.). Front. Plant Sci. 2015, 6, 841. [Google Scholar] [CrossRef]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Chardon, N.I.; Cornwell, W.K.; Flint, L.E.; Flint, A.L.; Ackerly, D.D. Topographic, latitudinal and climatic distribution of Pinus coulteri:geographic range limits are not at the edge of the climate envelope. Ecography 2015, 38, 590–601. [Google Scholar] [CrossRef]
- Runting, R.K.; Bryan, B.A.; Dee, L.E.; Maseyk, F.J.F.; Mandle, L.; Hamel, P.; Wilson, K.A.; Yetka, K.; Possingham, H.P.; Rhodes, J.R. Incorporating climate change into ecosystem service assessments and decisions: A review. Glob. Chang. Biol. 2016, 23, 28–29. [Google Scholar] [CrossRef]
- Ning, X.Z.; Geng, Q.C.; Zhong, G.Y. Investigation on Wild Resources of Vaccinium uliginosum and V. vitisidaea in The Greater Xing’an Mountains, Inner Mongolia. J. Inn. Mong. For. Sci. Technol. 2016, 42, 17–20. [Google Scholar]
- Bian, J.J.; Hao, Z.X.; Zheng, J.Y.; Ge, Q.S.; Yi, Y.H. The shift on boundary of climate regionalization in China from 1951 to 2010. Geogr. Res. 2013, 32, 1179–1187. [Google Scholar]
- Wang, J.; Wang, Y.; Feng, J.; Chen, C.; Chen, J.; Long, T.; Li, J.; Zang, R.; Li, J. Differential responses to climate and land-use changes in threatened chinese taxus species. Forests 2019, 10, 766. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Kumar, S.; Stohlgren, T. Maxent modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 1, 94–98. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Environmental Variable | Unit | V. uliginosum |
|---|---|---|---|
| BIO01 | Annual mean temperature | °C | |
| BIO02 | Mean diurnal range | °C | 0.1 |
| BIO03 | Isothermality (BIO02/BIO07) | % | |
| BIO04 | Temperature seasonality | - | |
| BIO05 | Max. temperature of warmest month | °C | |
| BIO06 | Min. temperature of coldest month | °C | |
| BIO07 | Annual temperature range | °C | 35.1 |
| BIO08 | Mean temperature of wettest quarter | °C | |
| BIO09 | Mean temperature of driest quarter | °C | |
| BIO10 | Mean temperature of warmest quarter | °C | 14.5 |
| BIO11 | Mean temperature of coldest quarter | °C | |
| BIO12 | Annual precipitation | mm | 24.5 |
| BIO13 | Precipitation of wettest month | mm | |
| BIO14 | Precipitation of driest month | mm | |
| BIO15 | Precipitation seasonality | mm | |
| BIO16 | Precipitation of wettest quarter | mm | |
| BIO17 | Precipitation of driest quarter | mm | |
| BIO18 | Precipitation of warmest quarter | mm | |
| BIO19 | Precipitation of coldest quarter | mm | 7.6 |
| Ele | Elevation | m | 7.3 |
| Land use | Land use | 6 | |
| Sym90 | Soil names in the FAO90 soil classification system | 4.9 |
| Decades | Total Suitable Habitats | Minimally Suitable Habitats | Moderately Suitable Habitats | Highly Suitable Habitats | |||||
|---|---|---|---|---|---|---|---|---|---|
| Area | Area | Area | Area | Area | Area | Area | Area | ||
| (104 km2) | (%) | (104 km2) | (%) | (104 km2) | (%) | (104 km2) | (%) | ||
| 1960–1990 | 83.8 | - | 36.48 | - | 31.43 | - | 15.89 | - | |
| SSP1-2.6 | 2050 | 81.95 | 97.79 | 34.95 | 95.81 | 31.53 | 100.32 | 15.47 | 97.36 |
| 2070 | 83.77 | 99.96 | 35.22 | 96.55 | 32.73 | 104.14 | 15.81 | 99.50 | |
| SSP2-4.5 | 2050 | 84.97 | 101.40 | 36.47 | 99.97 | 32.69 | 104.01 | 15.61 | 98.24 |
| 2070 | 85.61 | 102.16 | 36.19 | 99.21 | 32.59 | 103.69 | 16.82 | 105.85 | |
| SSP3-7.0 | 2050 | 82.40 | 98.33 | 34.59 | 94.82 | 31.33 | 99.68 | 16.48 | 103.71 |
| 2070 | 81.99 | 97.84 | 36.04 | 98.97 | 30.00 | 95.45 | 15.94 | 100.31 | |
| SSP5-8.5 | 2050 | 79.08 | 94.37 | 34.98 | 95.89 | 29.30 | 93.22 | 14.80 | 93.14 |
| 2070 | 81.37 | 97.10 | 33.95 | 93.06 | 30.01 | 95.48 | 17.40 | 109.50 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Qi, Y.; Wang, Q.; Wang, D. Prediction of the Potential Distribution of Vaccinium uliginosum in China Based on the Maxent Niche Model. Horticulturae 2022, 8, 1202. https://doi.org/10.3390/horticulturae8121202
Li Q, Qi Y, Wang Q, Wang D. Prediction of the Potential Distribution of Vaccinium uliginosum in China Based on the Maxent Niche Model. Horticulturae. 2022; 8(12):1202. https://doi.org/10.3390/horticulturae8121202
Chicago/Turabian StyleLi, Qian, Ying Qi, Qi Wang, and Delu Wang. 2022. "Prediction of the Potential Distribution of Vaccinium uliginosum in China Based on the Maxent Niche Model" Horticulturae 8, no. 12: 1202. https://doi.org/10.3390/horticulturae8121202
APA StyleLi, Q., Qi, Y., Wang, Q., & Wang, D. (2022). Prediction of the Potential Distribution of Vaccinium uliginosum in China Based on the Maxent Niche Model. Horticulturae, 8(12), 1202. https://doi.org/10.3390/horticulturae8121202