

Efficiency of Sodium and Calcium Chloride in Conferring Cross-Tolerance to Water Deficit in Periwinkle

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Flower Bud Initiation, Opening and Longevity
2.3. Plant Growth, Morphology and Biomass Allocation
2.4. Gas Exchange Traits
2.5. Chlorophyll and Carotenoid Contents
2.6. Stomatal and Epidermal Cell Anatomical Characteristics
2.7. Water Status
2.8. Proline Content
2.9. Electrolyte Leakage
2.10. Lipid Peroxidation
2.11. Enzymatic Activity
2.12. Statistical Analysis
3. Results
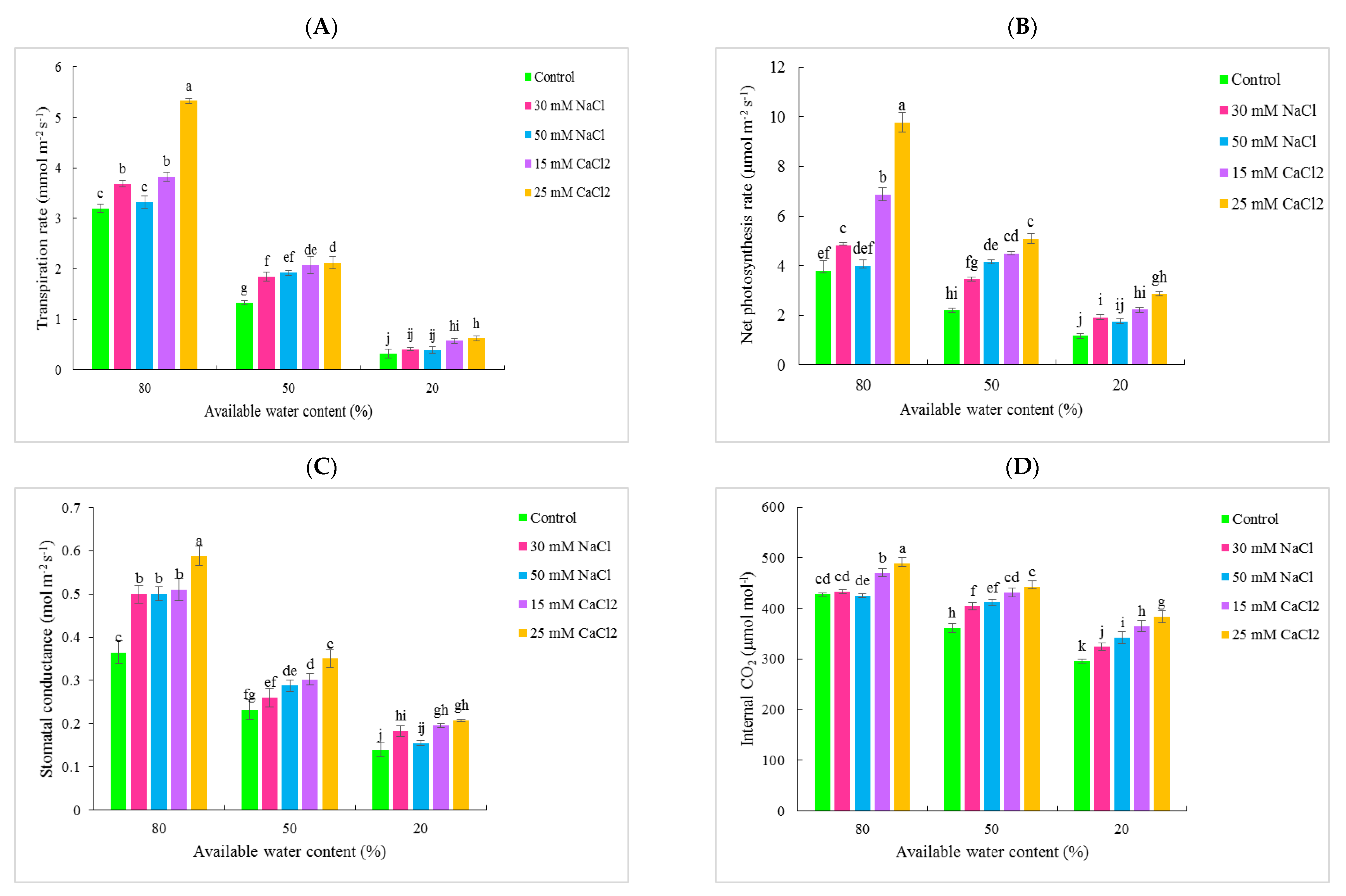
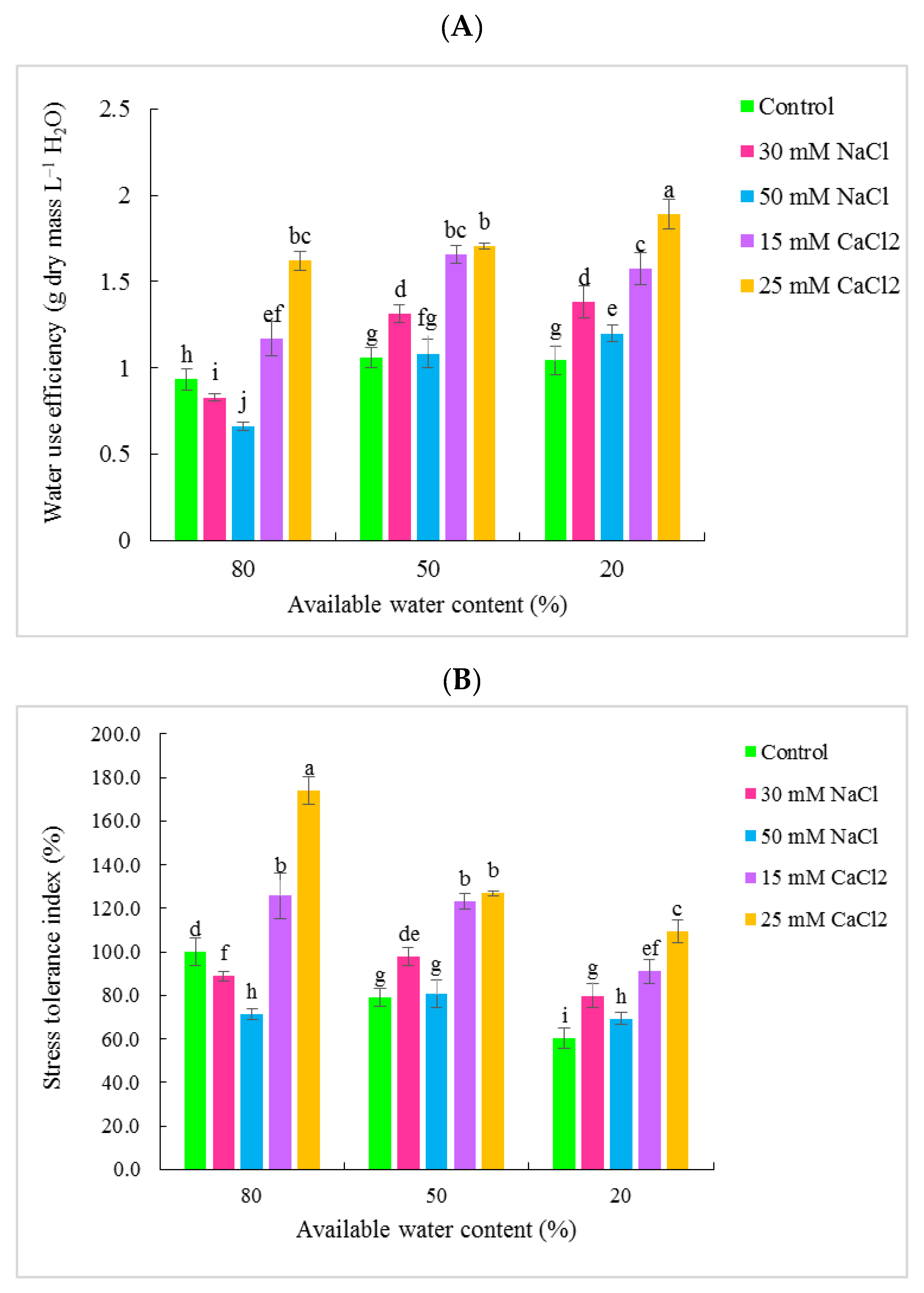
3.1. Gas Exchange Traits
3.2. Flower Bud Initiation, Opening and Longevity
3.3. Plant Growth, Morphology, and Biomass Allocation
3.4. Stomatal and Epidermal Cell Anatomical Characteristics
3.5. Water Status, as Well as Chlorophyll and Carotenoid Contents
3.6. Leaf Proline Content
3.7. Electrolyte Leakage and Lipid Peroxidation
3.8. Enzymatic Activity
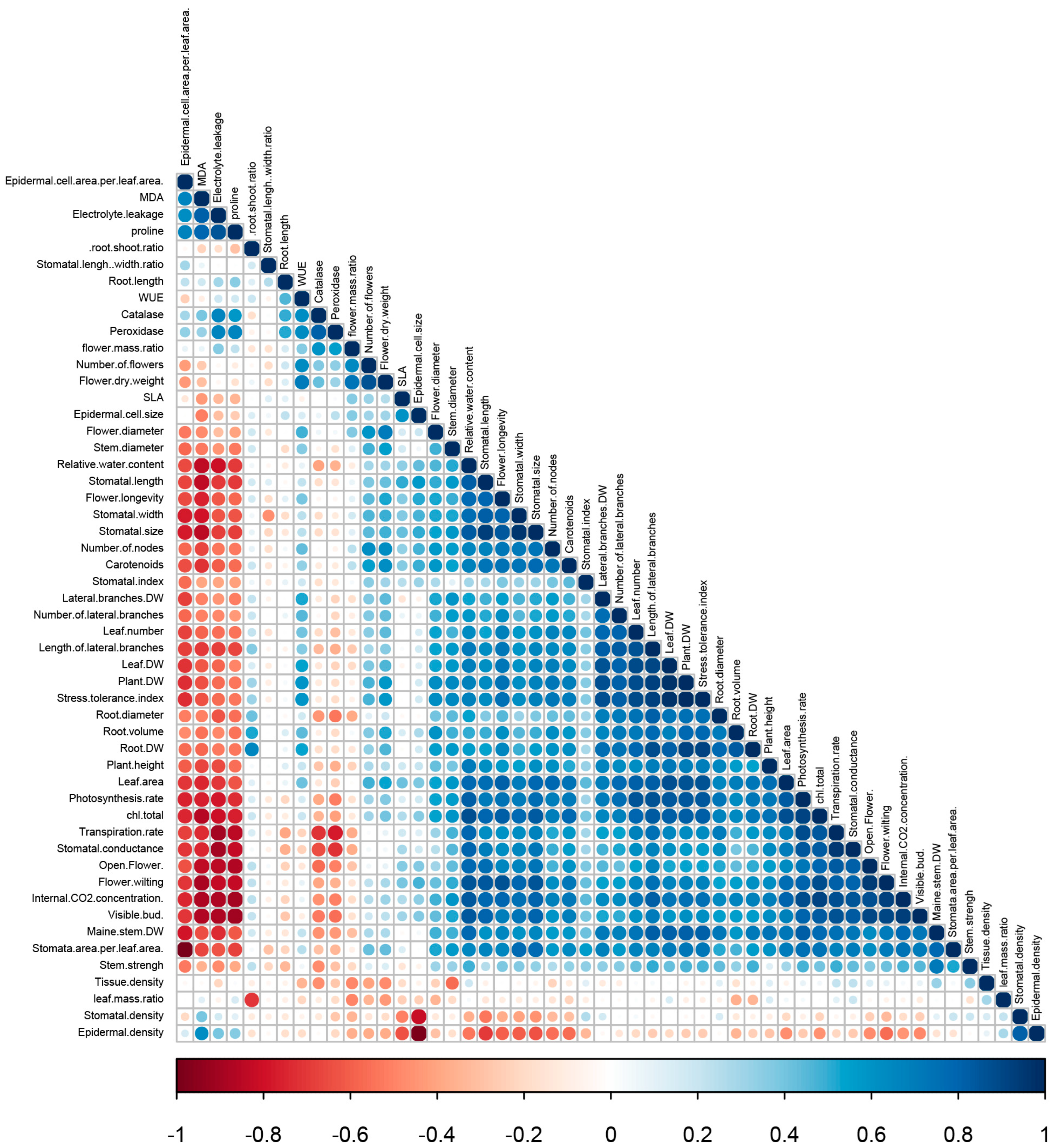
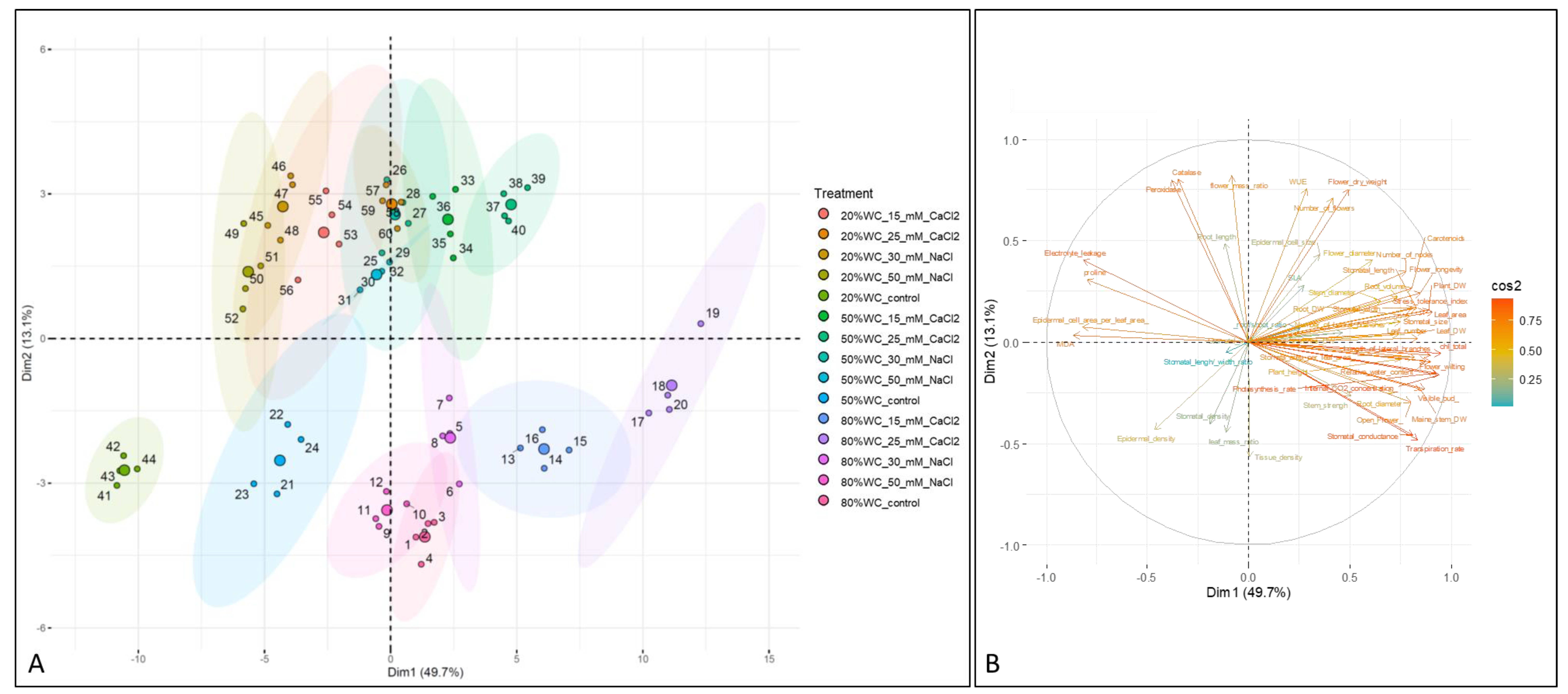
3.9. Principal Component Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CAT | Catalase |
| MDA | Malondialdehyde |
| PCA | Principal component analysis |
| POD | Peroxidase |
| ROS | Reactive oxygen species |
| RWC | Relative water content |
References
- Zomorrodi, N.; Rezaei Nejad, A.; Mousavi-Fard, S.; Feizi, H.; Tsaniklidis, G.; Fanourakis, D. Potency of titanium dioxide nanoparticles, sodium hydrogen sulfide and salicylic acid in ameliorating the depressive effects of water deficit on periwinkle ornamental quality. Horticulturae 2022, 8, 675. [Google Scholar] [CrossRef]
- Yousefzadeh, K.; Houshmand, S.; Shiran, B.; Mousavi-Fard, S.; Zeinali, H.; Nikoloudakis, N.; Gheisari, M.M.; Fanourakis, D. Joint effects of developmental stage and water deficit on essential oil traits (content, yield, composition) and related gene expression: A case study in two Thymus species. Agronomy 2022, 12, 1008. [Google Scholar] [CrossRef]
- Seifikalhor, M.; Niknam, V.; Aliniaeifard, S.; Didaran, F.; Tsaniklidis, G.; Fanourakis, D.; Teymoorzadeh, M.; Mousavi, S.H.; Bustachi, M.; Li, T. The regulatory role of γ-Aminobutyric acid in chickpea plants depends on drought tolerance and water scarcity level. Sci. Rep. 2022, 12, 7034. [Google Scholar] [CrossRef]
- Li, Z.G.; Gong, M. Mechanical stimulation-induced cross-adaptation in plants: An overview. J. Plant Biol. 2011, 54, 358–364. [Google Scholar] [CrossRef]
- Hossain, M.A.; Li, Z.G.; Hoque, T.S.; Burritt, D.J.; Fujita, M.; Munné-Bosch, S. Heat or cold priming-induced cross-tolerance to abiotic stresses in plants: Key regulators and possible mechanisms. Protoplasma 2018, 255, 399–412. [Google Scholar] [CrossRef]
- Ramegowda, V.; Da Costa, M.V.J.; Harihar, S.; Karaba, N.N.; Sreeman, S.M. Abiotic and Biotic Stress Interactions in Plants: A Cross-Tolerance Perspective. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Academic Press: Cambridge, MA, USA, 2020; pp. 267–302. [Google Scholar]
- Yang, X.; Li, Y.; Chen, H.; Huang, J.; Zhang, Y.; Qi, M.; Liu, Y.; Li, T. Photosynthetic response mechanism of soil salinity-induced cross-tolerance to subsequent drought stress in tomato plants. Plants 2020, 9, 363. [Google Scholar] [CrossRef]
- Pushpavalli, R.; Berger, J.D.; Turner, N.C.; Siddique, K.H.; Colmer, T.D.; Vadez, V. Cross-tolerance for drought, heat and salinity stresses in chickpea (Cicer arietinum L.). J. Agron. Crop Sci. 2020, 206, 405–419. [Google Scholar] [CrossRef]
- Farooq, S.; Hussain, M.; Jabran, K.; Hassan, W.; Rizwan, M.S.; Yasir, T.A. Osmopriming with CaCl2 improves wheat (Triticum aestivum L.) production under water-limited environments. Environ. Sci. Pollut. Res. 2017, 24, 13638–13649. [Google Scholar] [CrossRef]
- Das, A.; Ray, R.; Mandal, N.; Chakrabarti, K. An analysis of transcripts and enzyme profiles in drought stressed jute (Corchorus capsularis) and rice (Oryza sativa) seedlings treated with CaCl2, hydroxyapatite nano-particle and β-amino butyric acid. Plant Growth Regul. 2016, 79, 401–412. [Google Scholar] [CrossRef]
- Xu, C.; Li, X.; Zhang, L. The effect of calcium chloride on growth, photosynthesis, and antioxidant responses of Zoysia japonica under drought conditions. PLoS ONE 2013, 8, e68214. [Google Scholar] [CrossRef]
- Marković, M.; Šoštarić, J.; Kojić, A.; Popović, B.; Bubalo, A.; Bošnjak, D.; Stanisavljević, A. Zinnia (Zinnia elegans L.) and periwinkle (Catharanthus roseus (L.) G. Don) responses to salinity stress. Water 2022, 14, 1066. [Google Scholar] [CrossRef]
- Javadi Asayesh, E.; Aliniaeifard, S.; Askari, N.; Roozban, M.R.; Sobhani, M.; Tsaniklidis, G.; Woltering, E.J.; Fanourakis, D. Supplementary light with increased blue fraction accelerates emergence and improves development of the inflorescence in Aechmea, Guzmania and Vriesea. Horticulturae 2021, 7, 485. [Google Scholar] [CrossRef]
- Hassanvand, F.; Rezaei Nejad, A.; Fanourakis, D. Morphological and physiological components mediating the silicon-induced enhancement of geranium essential oil yield under saline conditions. Ind. Crops Prod. 2019, 134, 19–25. [Google Scholar] [CrossRef]
- Giday, H.; Fanourakis, D.; Kjaer, K.H.; Fomsgaard, I.S.; Ottosen, C.O. Threshold response of stomatal closing ability to leaf abscisic acid concentration during growth. J. Exp. Bot. 2014, 65, 4361–4370. [Google Scholar] [CrossRef]
- Fanourakis, D.; Giday, H.; Li, T.; Kambourakis, E.; Ligoxigakis, E.K.; Papadimitriou, M.; Strataridaki, A.; Bouranis, D.; Fiorani, F.; Heuvelink, E.; et al. Antitranspirant compounds alleviate the mild-desiccation-induced reduction of vase life in cut roses. Postharvest Biol. Technol. 2016, 117, 110–117. [Google Scholar] [CrossRef]
- Fanourakis, D.; Papadakis, V.M.; Psyllakis, E.; Tzanakakis, V.A.; Nektarios, P.A. The role of water relations and oxidative stress in the vase life response to prolonged storage: A case study in chrysanthemum. Agriculture 2022, 12, 185. [Google Scholar] [CrossRef]
- Jafari, S.; Mousavi-Fard, S.; Rezaei Nejad, A.; Moumivand, H.; Nikoloudakis, N.; Sorkheh, K.; Fanourakis, D. Chitosan and titanium dioxide in the nanoparticle form are more effective in improving seed yield and quality: A case study in five milk thistle ecotypes. Agronomy 2022, 12, 1827. [Google Scholar] [CrossRef]
- Taheri-Garavand, A.; Rezaei Nejad, A.; Fanourakis, D.; Fatahi, S.; Ahmadi-Majd, M. Employment of artificial neural networks for non-invasive estimation of leaf water status using color features: A case study in Spathiphyllum wallisii. Acta Physiol. Plant. 2021, 43, 78. [Google Scholar] [CrossRef]
- Chen, Y.; Fanourakis, D.; Tsaniklidis, G.; Aliniaeifard, S.; Yang, Q.; Li, T. Low UVA intensity during cultivation improves the lettuce shelf-life, an effect that is not sustained at higher intensity. Postharvest Biol. Technol. 2021, 172, 111376. [Google Scholar] [CrossRef]
- Paschalidis, K.; Fanourakis, D.; Tsaniklidis, G.; Tzanakakis, V.A.; Bilias, F.; Samara, E.; Kalogiannakis, K.; Debouba, F.J.; Ipsilantis, I.; Tsoktouridis, G.; et al. Pilot cultivation of the vulnerable cretan endemic Verbascum arcturus L.(scrophulariaceae): Effect of fertilization on growth and quality features. Sustainability 2021, 13, 14030. [Google Scholar] [CrossRef]
- Ahmadi-Majd, M.; Mousavi-Fard, S.; Rezaei Nejad, A.; Fanourakis, D. Carbon nanotubes in the holding solution stimulate flower opening and prolong vase life in carnation. Chem. Biol. Technol. Agric. 2022, 9, 15. [Google Scholar] [CrossRef]
- Ahmadi-Majd, M.; Rezaei Nejad, A.; Mousavi-Fard, S.; Fanourakis, D. Postharvest application of single, multi-walled carbon nanotubes and nanographene oxide improves rose keeping quality. J. Hortic. Sci. Biotechnol. 2022, 97, 346–360. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Sørensen, H.K.; Fanourakis, D.; Tsaniklidis, G.; Bouranis, D.; Rezaei Nejad, A.; Ottosen, C.O. Using artificial lighting based on electricity price without a negative impact on growth, visual quality or stomatal closing response in Passiflora. Sci. Hortic. 2020, 267, 109354. [Google Scholar] [CrossRef]
- Seif, M.; Aliniaeifard, S.; Arab, M.; Mehrjerdi, M.Z.; Shomali, A.; Fanourakis, D.; Li, T.; Woltering, E. Monochromatic red light during plant growth decreases the size and improves the functionality of stomata in chrysanthemum. Funct. Plant Biol. 2021, 48, 515–528. [Google Scholar] [CrossRef]
- Fanourakis, D.; Heuvelink, E.; Carvalho, S.M.P. Spatial heterogeneity in stomatal features during leaf elongation: An analysis using Rosa hybrida. Funct. Plant Biol. 2015, 42, 737–745. [Google Scholar] [CrossRef]
- Ahmadi-Majd, M.; Mousavi-Fard, S.; Rezaei Nejad, A.; Fanourakis, D. Nano-Selenium in the holding solution promotes rose and carnation vase life by improving both water relations and antioxidant status. J. Hortic. Sci. Biotechnol. 2022, 1–16. [Google Scholar] [CrossRef]
- Tabassum, T.; Farooq, M.; Ahmad, R.; Zohaib, A.; Wahid, A. Seed priming and transgenerational drought memory improves tolerance against salt stress in bread wheat. Plant Physiol. Biochem. 2017, 118, 362–369. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Panda, S.K.; Dutta, B.K. CaCl2 improves post-drought recovery potential in Camellia sinensis (L.) O. Kuntze. Plant Cell Rep. 2011, 30, 495–503. [Google Scholar] [CrossRef]
- Farooq, M.; Aziz, T.; Basra, S.M.A.; Wahid, A.; Khaliq, A.; Cheema, M.A. Exploring the role of calcium to improve chilling tolerance in hybrid maize. J. Agron. Crop Sci. 2008, 194, 350–359. [Google Scholar] [CrossRef]
- Moosavi-Nezhad, M.; Salehi, R.; Aliniaeifard, S.; Tsaniklidis, G.; Woltering, E.J.; Fanourakis, D.; Żuk-Gołaszewska, K.; Kalaji, H.M. Blue light improves photosynthetic performance during healing and acclimatization of grafted watermelon seedlings. Int. J. Mol. Sci. 2021, 22, 8043. [Google Scholar] [CrossRef]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef]
- Tanveer, K.; Gilani, S.; Hussain, Z.; Ishaq, R.; Adeel, M.; Ilyas, N. Effect of salt stress on tomato plant and the role of calcium. J. Plant Nutr. 2020, 43, 28–35. [Google Scholar] [CrossRef]
- Karabaliev, M. Effects of divalent cations on the formation and structure of solid supported lipid films. Bioelectrochemistry 2007, 71, 54–59. [Google Scholar] [CrossRef]
- Balantič, K.; Weiss, V.U.; Allmaier, G.; Kramar, P. Calcium ion effect on phospholipid bilayers as cell membrane analogues. Bioelectrochemistry 2022, 143, 107988. [Google Scholar] [CrossRef]
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.P.; Thuleau, P.; Mazars, C. Calcium sensors as key hubs in plant responses to biotic and abiotic stresses. Front. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef]
- Kurusu, T.; Kuchitsu, K.; Tada, Y. Plant signaling networks involving Ca2+ and Rboh/Nox-mediated ROS production under salinity stress. Front. Plant Sci. 2015, 6, 427. [Google Scholar] [CrossRef]
- Ali, L.G.; Nulit, R.; Ibrahim, M.H.; Yien, C.Y.S. Efficacy of KNO3, SiO2 and SA priming for improving emergence, seedling growth and antioxidant enzymes of rice (Oryza sativa), under drought. Sci. Rep. 2021, 11, 3864. [Google Scholar] [CrossRef]
- Saed-Moocheshi, A.; Shekoofa, A.; Sadeghi, H.; Pessarakli, M. Drought and salt stress mitigation by seed priming with KNO3 and urea in various maize hybrids: An experimental approach based on enhancing antioxidant responses. J. Plant Nutr. 2014, 37, 674–689. [Google Scholar] [CrossRef]
- Singhal, R.K.; Pandey, S.; Bose, B. Seed priming with Mg(NO3)2 and ZnSO4 salts triggers physio-biochemical and antioxidant defense to induce water stress adaptation in wheat (Triticum aestivum L.). Plant Stress 2021, 2, 100037. [Google Scholar] [CrossRef]
- Farman, M.; Nawaz, F.; Majeed, S.; Javeed, H.M.R.; Ahsan, M.; Ahmad, K.S.; Aurangzaib, M.; Bukhari, M.A.; Shehzad, M.A.; Hussain, M.B. Silicon seed priming combined with foliar spray of sulfur regulates photosynthetic and antioxidant systems to confer drought tolerance in maize (Zea mays L.). Silicon 2022, 14, 7901–7917. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Time to | Intact Flower Bud Longevity (d) | ||||
|---|---|---|---|---|---|---|
| Available Water Content (%) | Compound | Concentration (mM) | Visible Bud (d) | Open Flower (d) | Flower Wilting (d) | |
| 80 | - | 0 | 74.2 bc | 90.2 b | 97.5 de | 7.2 f |
| NaCl | 30 | 74.7 b | 92.5 ab | 103.2 c | 10.7 de | |
| 50 | 74.2 bc | 93.0 a | 102.5 c | 9.5 e | ||
| CaCl2 | 15 | 77.2 a | 92.2 ab | 106.2 b | 14.0 ab | |
| 25 | 78.2 a | 93.7 a | 109.5 a | 15.7 a | ||
| 50 | - | 0 | 69.0 fg | 84.5 d | 88.0 g | 3.5 g |
| NaCl | 30 | 71.2 de | 86.2 cd | 97.4 de | 11.1 cde | |
| 50 | 72.7 cd | 87.5 c | 98.0 de | 10.5 de | ||
| CaCl2 | 15 | 70.7 ef | 86.2 cd | 96.5 ef | 10.2 de | |
| 25 | 74.5 bc | 86.5 cd | 99.2 d | 12.7 bc | ||
| 20 | - | 0 | 58.2 j | 72.7 g | 74.7 i | 2.0 g |
| NaCl | 30 | 64.2 i | 76.7 f | 82.7 h | 6.0 f | |
| 50 | 67.5 gh | 81.0 e | 88.2 g | 7.2 f | ||
| CaCl2 | 15 | 66.7 h | 84.7 d | 94.7 f | 10.0 de | |
| 25 | 67.0 h | 86.0 cd | 97.7 de | 11.7 cd | ||
| Treatment | Lateral Branches | Main Stem | Number of Nodes | Leaf | Flower | Root | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Available Water Content (%) | Compound | Concentration (mM) | Number | Length (cm) | Length (cm) | Diameter (mm) | Strength (g cm–1) | Tissue Density (g cm–3) | Number | Area (cm2) | Number | Diameter (mm) | Length (cm) | Diameter (mm) | Volume (cm3) | |
| 80 | - | 0 | 8.0 de | 9.2 cd | 9.7 bcd | 3.40 cde | 18.9 a | 2.09 a | 9.0 de | 67.5 ef | 157 h | 3.5 fg | 33.4 c–f | 20.1 ef | 3.24 bc | 4.25 bc |
| NaCl | 30 | 8.3 d | 7.6 efg | 10.1 bc | 3.21 efg | 14.3 bcd | 1.77 abc | 10.0 bc | 75.5 cd | 234 d | 5.5 b–e | 29.9 fgh | 22.0 cde | 2.84 c–f | 3.50 cd | |
| 50 | 8.0 de | 6.1 h | 9.5 cd | 3.63 bc | 16.0 b | 1.56 bcd | 8.8 e | 53.5 hi | 180 g | 3.2 fg | 28.7 gh | 18.7 f | 2.50 fgh | 2.00 f | ||
| CaCl2 | 15 | 12.8 a | 11.2 b | 12.7 a | 3.34 def | 18.7 a | 2.16 a | 10.5 ab | 87.3 b | 273 b | 4.0 efg | 32.7 c–g | 24.6 ab | 3.62 ab | 5.25 a | |
| 25 | 13.8 a | 14.6 a | 12.0 a | 3.91 a | 19.7 a | 1.67 bcd | 11.3 a | 106.3 a | 307 a | 7.0 b | 39.5 a | 20.2 ef | 3.95 a | 5.50 a | ||
| 50 | - | 0 | 8.5 cd | 6.1 h | 9.2 cde | 3.29 def | 12.0 e | 1.41 cd | 7.8 f | 63.8 fg | 131 i | 3.0 g | 30.2 e–h | 22.0 cde | 2.89 c–f | 3.50 cd |
| NaCl | 30 | 9.0 cd | 7.2 fgh | 9.5 cd | 3.54 bcd | 12.4 de | 1.27 de | 9.5 cde | 65.5 fg | 203 f | 6.2 bcd | 35.6 abc | 23.8 abc | 2.82 d–g | 5.00 ab | |
| 50 | 8.3 cd | 6.3 gh | 8.1 ef | 3.44 cde | 12.3 de | 1.34 d | 10.0 bc | 72.5 de | 148 h | 6.0 bcd | 34.9 bcd | 20.7 de | 2.25 hi | 3.50 cd | ||
| CaCl2 | 15 | 11.0 b | 10.1 bc | 10.0 bc | 3.66 abc | 9.5 f | 0.91 e | 10.5 ab | 81.3 bc | 250 c | 6.2 bcd | 34.7 b–e | 23.0 bc | 3.00 cde | 5.75 a | |
| 25 | 11.0 b | 8.8 cde | 10.7 b | 3.75 ab | 15.0 bc | 1.37 d | 10.5 ab | 75.5 cd | 237 d | 10.2 a | 38.6 ab | 22.0 cde | 3.02 cd | 5.00 ab | ||
| 20 | - | 0 | 7.5 de | 3.6 i | 7.7 f | 3.02 g | 13.2 cde | 1.85 ab | 7.3 f | 57.5 hi | 90 k | 2.7 g | 25.9 h | 22.2 cd | 2.52 fgh | 1.75 f |
| NaCl | 30 | 8.3 d | 4.3 i | 9.2 cde | 3.33 def | 12.7 de | 1.47 bcd | 9.8 bcd | 59.5 gh | 183 g | 6.7 bc | 32.3 c–g | 22.5 cd | 2.44 gh | 3.00 de | |
| 50 | 6.5 e | 4.2 i | 8.6 def | 3.10 fg | 12.4 de | 1.65 bcd | 8.8 e | 53.0 i | 116 j | 4.7 def | 30.5 d–g | 23.1 bc | 1.92 i | 2.13 ef | ||
| CaCl2 | 15 | 8.5 cd | 6.3 gh | 9.0 cde | 3.19 efg | 13.5 cde | 1.78 abc | 8.8 e | 64.0 fg | 157 h | 5.2 cde | 30.4 d–g | 24.5 ab | 2.62 e–h | 3.63 cd | |
| 25 | 10.0 bc | 8.5 def | 9.7 bcd | 3.52 bcd | 14.0 b–e | 1.44 cd | 9.8 bcd | 71.8 de | 219 e | 7.0 b | 34.7 bcd | 25.1 a | 2.56 fgh | 3.25 d | ||
| Treatment | Main Stem | Lateral Branches | Leaf | Flower | Root | Plant | Root to Shoot Ratio | Specific Leaf Area (cm2 g−1) | Leaf Mass Ratio | Flower Mass Ratio | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Available Water Content (%) | Compound | Concentration (mM) | Dry Weight (g) | |||||||||
| 80 | - | 0 | 0.17 c | 0.03 cd | 0.59 ef | 0.06 hi | 0.34 bc | 1.21 d | 39.3 a | 264 c | 0.49 ef | 0.06 g |
| NaCl | 30 | 0.13 de | 0.01 ef | 0.60 e | 0.10 fg | 0.21 ef | 1.07 f | 24.6 efg | 385 a | 0.56 ab | 0.09 cd | |
| 50 | 0.13 def | 0.01 ef | 0.48 hi | 0.06 hi | 0.16 fg | 0.86 h | 24.2 fg | 373 a | 0.56 ab | 0.07 ef | ||
| CaCl2 | 15 | 0.18 b | 0.04 bc | 0.85 b | 0.08 gh | 0.35 b | 1.52 b | 30.6 c–f | 322 b | 0.56 ab | 0.06 g | |
| 25 | 0.20 a | 0.07 a | 1.13 a | 0.18 b | 0.50 a | 2.10 a | 31.6 b–e | 270 c | 0.54 bc | 0.09 de | ||
| 50 | - | 0 | 0.11 gh | 0.03 bc | 0.50 hi | 0.06 hi | 0.23 de | 0.95 g | 32.6 a–d | 262 c | 0.52 cde | 0.07 fg |
| NaCl | 30 | 0.12 efg | 0.02 def | 0.53 gh | 0.17 bc | 0.33 bc | 1.18 de | 38.8 ab | 317 a | 0.45 g | 0.15 a | |
| 50 | 0.11 gh | 0.02 def | 0.47 ij | 0.14 de | 0.22 ef | 0.97 g | 29.3 c–g | 317 b | 0.48 f | 0.15 a | ||
| CaCl2 | 15 | 0.12 fg | 0.04 b | 0.79 c | 0.16 cd | 0.36 b | 1.49 b | 32.4 a–d | 314 b | 0.53 bcd | 0.11 bc | |
| 25 | 0.17 bc | 0.04 b | 0.75 cd | 0.22 a | 0.33 bc | 1.53 b | 27.6 d–g | 312 b | 0.49 ef | 0.15 a | ||
| 20 | - | 0 | 0.10 i | 0.01 ef | 0.42 j | 0.05 i | 0.13 g | 0.73 i | 22.9 g | 205 d | 0.58 a | 0.08 ef |
| NaCl | 30 | 0.11 gh | 0.01 ef | 0.49 hi | 0.14 de | 0.19 efg | 0.96 g | 25.3 d–g | 367 a | 0.52 c–f | 0.15 a | |
| 50 | 0.10 hi | 0.01 f | 0.45 ij | 0.10 f | 0.16 fg | 0.84 h | 24.3 efg | 255 c | 0.54 bc | 0.12 b | ||
| CaCl2 | 15 | 0.12 fg | 0.02 de | 0.55 fg | 0.11 f | 0.28 cd | 1.10 ef | 35.1 abc | 310 b | 0.51 def | 0.10 cd | |
| 25 | 0.13 d | 0.04 bc | 0.72 d | 0.13 e | 0.28 cd | 1.32 c | 27.8 c–g | 326 b | 0.55 bc | 0.10 cd | ||
| Treatment | Epidermal Cell | Stomatal | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Available Water Content (%) | Compound | Concentration (mM) | Density (mm–2) | Size (µm2) | Area per Leaf Area (%) | Density (mm2) | Index (%) | Length (µm) | Width (µm) | Size (µm2) | Length to Width Ratio | Area per Leaf Area (%) |
| 80 | - | 0 | 575 cd | 1666 ef | 95.8 fg | 155 d | 21.2 cd | 26.5 gh | 10.1 fgh | 536 gh | 2.65 a–d | 4.16 cd |
| NaCl | 30 | 500 h | 1911 a | 95.4 h | 145 e | 22.5 ab | 28.3 b–e | 11.1 bc | 632 b | 2.54 bcd | 4.59 b | |
| 50 | 572 cd | 1677 ef | 95.9 f | 135 f | 19.1 fg | 28.7 bc | 10.6 c–f | 609 bcd | 2.71 ab | 4.12 d | ||
| CaCl2 | 15 | 510 gh | 1878 ab | 95.7 fgh | 137 f | 21.2 cd | 29.0 b | 10.7 cde | 621 bc | 2.71 ab | 4.27 bcd | |
| 25 | 600 c | 1575 f | 94.5 i | 150 de | 20.0 ef | 30.4 a | 12.0 a | 731 a | 2.54 bcd | 5.48 a | ||
| 50 | - | 0 | 695 b | 1381 g | 96.0 def | 175 b | 20.1 ef | 25.0 i | 9.0 i | 455 i | 2.76 a | 3.99 def |
| NaCl | 30 | 527 fgh | 1826 ab | 96.3 cd | 132 f | 20.0 ef | 27.6 c–g | 10.0 gh | 553 fgh | 2.77 a | 3.67 fg | |
| 50 | 535 efg | 1798 bcd | 95.9 ef | 137 f | 20.4 de | 27.1 fgh | 10.9 bcd | 594 cde | 2.48 d | 4.08 de | ||
| CaCl2 | 15 | 517 gh | 1863 ab | 96.2 cde | 132 f | 20.4 de | 27.8 c–f | 10.2 e–h | 568 efg | 2.74 a | 3.77 efg | |
| 25 | 557 def | 1699 cde | 94.7 i | 165 c | 22.8 a | 28.3 bcd | 11.3 b | 641 b | 2.51 cd | 5.29 a | ||
| 20 | - | 0 | 800 a | 1211 h | 96.9 a | 190 a | 19.1 fg | 20.5 j | 7.9 j | 325 j | 2.59 a–d | 3.09 i |
| NaCl | 30 | 530 gh | 1821 ab | 96.5 bc | 132 f | 19.9 ef | 26.3 h | 9.8 h | 520 h | 2.66 abc | 3.44 gh | |
| 50 | 535 efg | 1808 abc | 96.7 ab | 122 g | 18.6 g | 26.6 gh | 10.1 e–h | 541 fgh | 2.62 a–d | 3.32 hi | ||
| CaCl2 | 15 | 517 gh | 1881 ab | 96.5 bc | 125 g | 19.5 efg | 27.2 e–h | 10.2 e–h | 556 fgh | 2.66 a–d | 3.48 gh | |
| 25 | 562 de | 1698 de | 95.5 gh | 155 d | 21.6 bc | 27.5 d–g | 10.4 d–g | 575 def | 2.63 a–d | 4.46 bc | ||
| Treatment | Relative Water Content (%) | Chlorophyll a | Chlorophyll b | Chlorophyll (a + b) | Carotenoid | Electrolyte Leakage (%) | MDA | Proline | Catalase | Peroxidase | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Available Water Content (%) | Compound | Concentration (mM) | Content (mg g−1 FW) | Content (µmol g−1 FW) | Activity (µmol min−1 g−1 FW) | |||||||
| 80 | - | 0 | 80.9 bcd | 6.0 cd | 3.2 ef | 9.2 ef | 2.2 cde | 23.7 f | 0.280 efg | 4.19 e | 0.001 j | 0.010 k |
| NaCl | 30 | 81.7 bc | 6.4 bcd | 3.8 c | 10.3 cd | 2.5 bc | 23.1 f | 0.269 fg | 4.19 e | 0.007 i | 0.016 jk | |
| 50 | 82.5 b | 6.4 bcd | 3.4 de | 9.8 cde | 2.2 cde | 23.6 f | 0.274 efg | 4.12 e | 0.008 h | 0.018 j | ||
| CaCl2 | 15 | 90.9 a | 8.6 a | 4.3 b | 13.0 b | 2.4 cd | 20.1 g | 0.238 gh | 4.09 e | 0.010 g | 0.014 jk | |
| 25 | 93.8 a | 9.4 a | 5.0 a | 14.5 a | 3.1 a | 22.3 fg | 0.215 h | 3.34 e | 0.008 h | 0.015 jk | ||
| 50 | - | 0 | 66.6 f | 4.3 fg | 2.1 ij | 6.4 h | 1.5 f | 36.0 bcd | 0.395 b | 6.74 d | 0.012 f | 0.027 i |
| NaCl | 30 | 77.6 cde | 6.9 bc | 2.4 hi | 9.3 def | 2.4 cd | 33.0 d | 0.311 def | 6.57 d | 0.013 e | 0.075 g | |
| 50 | 77.2 cde | 6.3 bcd | 2.9 fg | 9.3 def | 2.4 cd | 34.2 cd | 0.312 def | 5.91 d | 0.014 d | 0.085 ef | ||
| CaCl2 | 15 | 76.2 de | 6.9 bc | 3.5 cde | 10.5 c | 2.5 c | 29.3 e | 0.299 def | 5.88 d | 0.016 b | 0.083 f | |
| 25 | 81.6 bc | 7.0 b | 3.6 cd | 10.7 c | 2.9 ab | 28.2 e | 0.290 def | 5.83 d | 0.016 b | 0.092 e | ||
| 20 | - | 0 | 59.3 g | 3.5 g | 1.2 l | 4.7 i | 1.4 f | 43.0 a | 0.533 a | 16.85 a | 0.014 de | 0.065 h |
| NaCl | 30 | 73.8 e | 4.9 ef | 1.8 jk | 6.7 h | 2.1 de | 36.5 bc | 0.380 bc | 12.95 b | 0.015 c | 0.114 d | |
| 50 | 73.2 e | 4.8 ef | 1.6 k | 6.5 h | 2.0 e | 37.7 b | 0.386 b | 13.49 b | 0.015 b | 0.129 c | ||
| CaCl2 | 15 | 74.7 e | 5.6 de | 2.1 ij | 7.8 g | 2.1 de | 37.5 b | 0.335 cd | 12.57 b | 0.015 b | 0.160 b | |
| 25 | 76.5 de | 6.0 cd | 2.8 gh | 8.8 f | 2.2 cde | 35.1 bcd | 0.322 de | 10.63 c | 0.017 a | 0.168 a | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zomorrodi, N.; Rezaei Nejad, A.; Mousavi-Fard, S.; Feizi, H.; Nikoloudakis, N.; Fanourakis, D. Efficiency of Sodium and Calcium Chloride in Conferring Cross-Tolerance to Water Deficit in Periwinkle. Horticulturae 2022, 8, 1091. https://doi.org/10.3390/horticulturae8111091
Zomorrodi N, Rezaei Nejad A, Mousavi-Fard S, Feizi H, Nikoloudakis N, Fanourakis D. Efficiency of Sodium and Calcium Chloride in Conferring Cross-Tolerance to Water Deficit in Periwinkle. Horticulturae. 2022; 8(11):1091. https://doi.org/10.3390/horticulturae8111091
Chicago/Turabian StyleZomorrodi, Nahid, Abdolhossein Rezaei Nejad, Sadegh Mousavi-Fard, Hassan Feizi, Nikolaos Nikoloudakis, and Dimitrios Fanourakis. 2022. "Efficiency of Sodium and Calcium Chloride in Conferring Cross-Tolerance to Water Deficit in Periwinkle" Horticulturae 8, no. 11: 1091. https://doi.org/10.3390/horticulturae8111091
APA StyleZomorrodi, N., Rezaei Nejad, A., Mousavi-Fard, S., Feizi, H., Nikoloudakis, N., & Fanourakis, D. (2022). Efficiency of Sodium and Calcium Chloride in Conferring Cross-Tolerance to Water Deficit in Periwinkle. Horticulturae, 8(11), 1091. https://doi.org/10.3390/horticulturae8111091