
Impact of Bee and Fly Pollination on Physical and Biochemical Properties of Strawberry Fruit

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Pollinator Abundance
2.3. Foraging Behavior
2.4. Single-Visit Effectiveness
2.5. Physical Parameters
2.6. Biochemical Parameters
2.7. Data Analysis
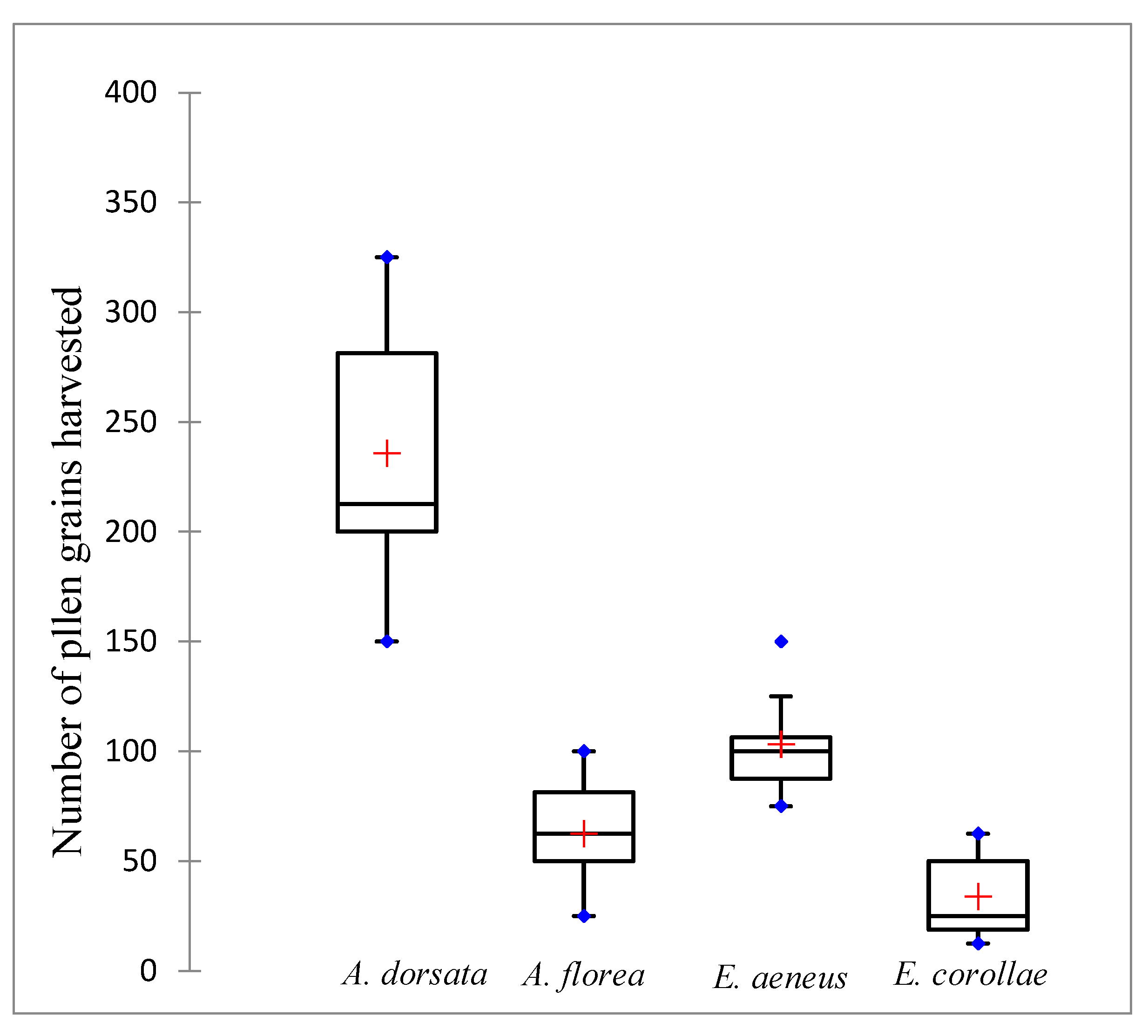
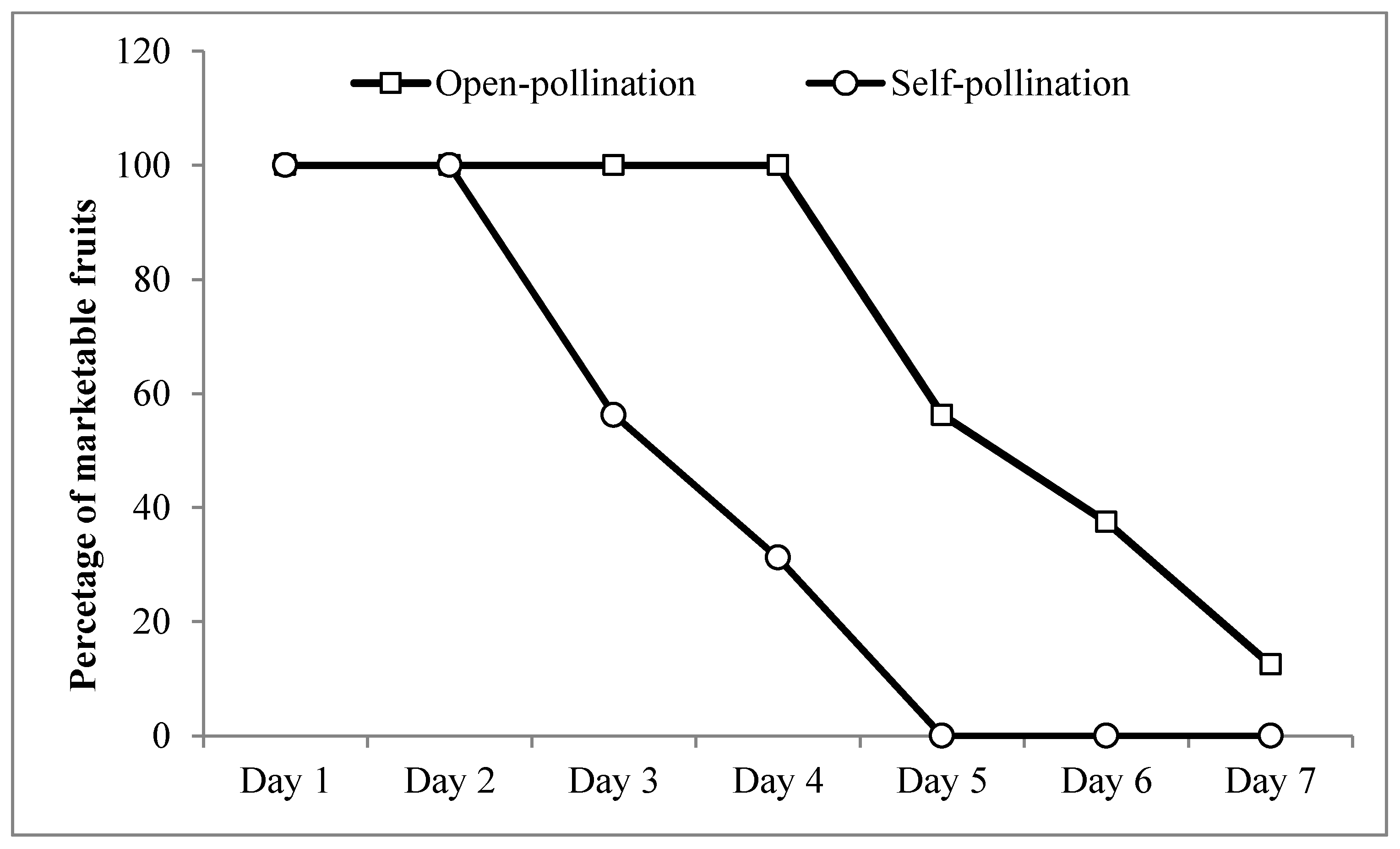
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Free, J.B. Hormonal regulation of ripening in the strawberry, a non-climacteric fruit. J. Appl. Ecol. 1993, 174, 402–406. [Google Scholar]
- Klatt, B.K.; Holzschuh, A.; Westphal, C.; Clough, Y.; Smit, I.; Pawelzik, E.; Tscharntke, T. Bee pollination improves crop quality, shelf life and commercial value. Proc. R. Soc. B 2013, 281, 2013–2440. [Google Scholar] [CrossRef]
- Carew, J.G.; Morretini, M.; Battey, N.H. Misshapen fruits in strawberry. Small Fruits Rev. 2003, 2, 37–50. [Google Scholar] [CrossRef]
- Vincent, I.C.; Hilla, R.; Campling, R.C. A note on the use of rapeseed, sunflower and soyabean meals as protein sources in compound foods for milking cattle. Ani. Prod. Sci. 1990, 50, 541–543. [Google Scholar] [CrossRef]
- Horth, L.; Campbell, L.A. Supplementing small farms with native mason bees increases strawberry size and growth rate. J. Appl. Ecol. 2018, 55, 591–599. [Google Scholar] [CrossRef]
- Albano, S.; Salvado, E.; Duarte, S.; Mexia, A.; Borges, P.A. Pollination effectiveness of different strawberry floral visitors in Ribatejo, Portugal: Selection of potential pollinators. Adv. Horti. Sci. 2009, 24, 246–253. [Google Scholar]
- Çolak, A.M.M.; Sahinler, N.; Islamoglu, M. The Effect of Honeybee Pollination on Productivity and Quality of Strawberry. Alınteri J. Agric. Sci. 2017, 32, 87–90. [Google Scholar]
- MacInnis, G.; Forrest, J.R. Pollination by wild bees yields larger strawberries than pollination by honey bees. J. Appl. Ecol. 2019, 56, 824–832. [Google Scholar] [CrossRef]
- Masyitah, S.; Rauf, A.; Maryana, N.; Kahono, S. Pollination services of flower-visiting insects in strawberry fields in Ciwidey, Bandung. Indones. J. Entomol. 2019, 16, 115–127. [Google Scholar] [CrossRef]
- Hodgkiss, D.; Brown, M.J.; Fountain, M.T. Syrphine hoverflies are effective pollinators of commercial strawberry. J. Pollinat. Ecol. 2018, 22, 208–220. [Google Scholar] [CrossRef]
- Csukasi, F.; Osorio, S.; Gutierrez, J.R.; Kitamura, J.; Giavalisco, P.; Nakajima, M.; Fernie, A.R.; Rathjen, J.P.; Botella, M.A.; Valpuesta, V.; et al. Gibberellin biosynthesis and signalling during development of the strawberry receptacle. New Phytol. 2011, 191, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Muñoz, P.; Almenar, E.; Ocio, M.J.; Gavara, R. Effect of calcium dips and chitosan coatings on postharvest life of strawberries (Fragaria x ananassa). Postharvest. Biol. Technol. 2006, 39, 247–253. [Google Scholar] [CrossRef]
- Klatt, B.K.; Klaus, F.; Westphal, C.; Tscharntke, T. Enhancing crop shelf life with pollination. Agr. Food Sec. 2014, 3, 14. [Google Scholar] [CrossRef]
- Fait, A.; Hanhineva, K.; Beleggia, R.; Dai, N.; Rogachev, I.; Nikiforova, V.J.; Fernie, A.R.; Aharoni, A. Reconfiguration of the achene and receptacle metabolic networks during strawberry fruit development. Plant Physiol. 2008, 148, 730–750. [Google Scholar] [CrossRef] [PubMed]
- Abrol, D.P.; Gorka, A.K.; Ansari, M.J.; Al-Ghamdi, A.; Al-Kahtani, S. Impact of insect pollinators on yield and fruit quality of strawberry. Saudi J. Biol. Sci. 2019, 26, 524–530. [Google Scholar] [CrossRef]
- Wietzke, A.; Westphal, C.; Gras, P.; Kraft, M.; Pfohl, K.; Karlovsky, P.; Pawelzik, E.; Tscharntke, T.; Smit, I. Insect pollination as a key factor for strawberry physiology and marketable fruit quality. Agric. Ecosyst. Environ. 2018, 258, 197–204. [Google Scholar] [CrossRef]
- Dung, C.D.; Wallace, H.M.; Bai, S.H.; Ogbourne, S.M.; Trueman, S.J. Cross-pollination affects fruit colour, acidity, firmness and shelf life of self-compatible strawberry. PLoS ONE 2021, 16, 0256964. [Google Scholar] [CrossRef]
- Ali, M.; Saeed, S.; Sajjad, A.; Whittington, A. In search of the best pollinators for canola (Brassica napus L.) production in Pakistan. Pak. J. Zool. 2011, 46, 53–361. [Google Scholar] [CrossRef]
- Ali, M.; Saeed, S.; Sajjad, A.; Bashir, M.A. Exploring the Best Native Pollinators for Pumpkin (Cucurbitapepo) Production in Punjab, Pakistan. Pak. J. Zool. 2014, 46, 557–561. [Google Scholar]
- Artz, D.R.; Hsu, C.L.; Nault, B.A. Influence of honey bee, Apis mellifera, hives and field size on foraging activity of native bee species in pumpkin fields. Environ. Entomol. 2011, 40, 1144–1158. [Google Scholar] [CrossRef]
- Brittain, C.; Williams, N.; Kremen, C.; Klein, A.M. Synergistic effects of non-Apis bees and honey bees for pollination services. Proc. R. Soc. B. 2013, 280, 20122767. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Greenleaf, S.S.; Kremen, C. Wild bees enhance honey bees’ pollination of hybrid sunflower. Proc. Natl. Acad. Sci. USA 2006, 103, 13890–13895. [Google Scholar] [CrossRef] [PubMed]
- Rader, R.; Howlett, B.G.; Cunningham, S.A.; Westcott, D.A.; Newstrom, L.E.; Loyd, L.; Walker, M.K.; Teulon, D.A.; Edwards, W. Alternative pollinator taxa are equally efficient but not as effective as the honeybee in a mass flowering crop. J. Appl. Ecol. 2009, 46, 1080–1087. [Google Scholar] [CrossRef]
- Saeed, S.; Malik, S.A.; Dad, K.; Sajjad, A.; Ali, M. In search of the best native pollinators for bitter gourd (Momordica charantia L.) pollination in Multan, Pakistan. Pak. J. Zool. 2012, 102, 467–473. [Google Scholar]
- Sajjad, A.; Saeed, S.; Masood, A. Pollinator community of onion (Allium cepa L.) and its role in crop reproductive success. Pak. J. Zool. 2008, 40, 451–456. [Google Scholar]
- Khan, S. Climate classification of Pakistan. Int. J. Econ. Environ. Geol. 2019, 10, 60–71. [Google Scholar]
- Canto-Aguilar, M.A.; Parra-Tabla, V. Importance of conserving alternative pollinators: Assessing the pollination efficiency of the squash bee, Peponapis limitaris in Cucurbita moschata (Cucurbitaceae). J. Insect. Conserv. 2000, 4, 201–208. [Google Scholar] [CrossRef]
- Lynch, J.M. Use of AOAC International method performance statistics in the laboratory. J. AOAC Int. 1998, 81, 679–684. [Google Scholar] [CrossRef][Green Version]
- Horowitz, R.M.; Gentili, B. Flavonoid compounds of citrus. III. Isolation and structure of eriodictyol glycoside. J. Am. Chem. Soc. 1960, 82, 2803–2806. [Google Scholar] [CrossRef]
- Ruck, J.A. Chemical Method for Fruit and Vegetable Products; Research Station, Summerland, Department of Agriculture: Ottawa, ON, Canada, 1961. [Google Scholar]
- Roussos, P.A.; Denaxa, N.; Damvakaris, T. Strawberry fruit quality attributes after application of plant growth stimulating compounds. Sci. Hortic. 2009, 119, 138–146. [Google Scholar] [CrossRef]
- Sahli, H.F.; Conner, J.K. Visitation, effectiveness, and efficiency of 15 genera of visitors to wild radish, Raphanus raphanistrum (Brassicaceae). Am. J. Bot. 2007, 94, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Agarwal, O.P.; Mishra, R.C. Foraging rates of different Apis species visiting parental lines of Brassica napus L. Zoos’ Print J. 2006, 21, 2226–2227. [Google Scholar] [CrossRef]
- Ariza, M.T.; Soria, C.; Medina, J.J.; Martinez-Ferri, E. Fruit misshapen in strawberry cultivars (Fragaria x ananassa) is related to achenes functionality. Ann. Appl. Biol. 2010, 158, 130–138. [Google Scholar] [CrossRef]
- Grace, A.; Jane, O.; Jared, M.; Christine, K.; Dinah, M.; Patricia, N.; David, P.; Blaise, O.; Kamunyu, K. Productivity of Tomato in the Greenhouse Using Bee Pollination. Inter. J. Appl. Agric. Sci. 2017, 3, 161–435. [Google Scholar] [CrossRef]
- Caner, C.; Aday, M.S.; Demir, M. Extending the quality of fresh strawberries by equilibrium modified atmosphere packaging. Eur. Food Res. Technol. 2008, 227, 1575–1583. [Google Scholar] [CrossRef]
- Shin, Y.S.; Park, S.D.; Kim, H.S. Influence of pollination methods on fruit development and sugar contents of oriental melon (Cucumis melo L. cv. Sagyejeol-Ggul). Sci. Hortic. 2007, 112, 388–392. [Google Scholar] [CrossRef]
- Bashir, M.A.; Alvi, A.M.; Khan, K.A.; Rehmani, M.I.A.; Ansari, M.J.; Atta, S.; Ghramh, H.A.; Batool, T.; Tariq, M. Role of pollination in yield and physicochemical properties of tomatoes (Lycopersicon esculentum). Saud. J. Biol. Sci. 2018, 25, 91–1297. [Google Scholar] [CrossRef]
- Iftikhar, F.; Masood, M.A.; Waghchoure, S.E. Comparison of Apis cerana Apis dorsata Apis florea Apis mellifera honey from different areas of Pakistan. Asian J. Exp. Biol. Sci. 2011, 2, 399–403. [Google Scholar]
- Guérin, G.; Renaud, C.; Maucourt, M.; Rolin, D.; Roudeillac, P.; Denoyes-Rothan, B.; Lerceteau-Kohler, E.; Moing, A. QTL Analysis for Sugars and Organic Acids in Strawberry Fruits. Int. Strawb. Symp. 2004, 708, 573–578. [Google Scholar]
- Watts, S.; Ovalle, D.H.; Herrera, M.M.; Ollerton, J. Pollinator effectiveness of native and non-native flower visitors to an apparently generalist Andean shrub, Duranta mandonii (Verbenaceae). Plant Sp. Biol. 2012, 27, 147–158. [Google Scholar] [CrossRef]
- Malagodi-Braga, K.S.; Kleinert, A.M. Como o comportamento das abelhas na flor do morangueiro (Fragaria ananassa Duchesne) influencia a formação dos frutos? Biosci. J. 2007, 23, 76–81. [Google Scholar]
- Chagnon, M.; Gingras, J.; De-Oliveira, D. Complementary Aspects of Strawberry Pollination by Honey and Indigenous Bees (Hymenoptera). J. Ecol. Entomol. 1993, 86, 416–420. [Google Scholar] [CrossRef]
- Alonso, C.; Herrera, C.M.; Ashman, T.L. A piece of the puzzle: A method for comparing pollination quality and quantity across multiple species and reproductive events. New Phytol. 2012, 193, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Sajjad, A.; Farooqi, M.A.; Bashir, M.A.; Aslam, M.N.; Nafees, M.; Aslam, M.N.; Adnan, M.; Khan, K.A. Assessing indigenous and local knowledge of farmers about pollination services in cucurbit agro-ecosystem of Punjab, Pakistan. Saudi J. Biol. Sci. 2020, 27, 189–194. [Google Scholar] [CrossRef]
- Aigner, P.A. Optimality Modeling and Fitness Trade-Offs: When Should Plants Become Pollinator Specialists? Oikos 2001, 95, 177–184. [Google Scholar] [CrossRef]
- Majewska, A.A.; Altizer, S. Planting gardens to support insect pollinators. Conserv. Biol. 2020, 34, 15–25. [Google Scholar] [CrossRef]
- Kremen, C.; Williams, N.M.; Bugg, R.L.; Fay, J.P.; Thorp, R.W. The area requirements of an ecosystem service: Crop pollination by native bee communities in California. Ecol. Lett. 2004, 7, 1109–1119. [Google Scholar] [CrossRef]
- Kleijn, D.; Van Langevelde, F. Interacting effects of landscape context and habitat quality on flower visiting insects in agricultural landscapes. Basic Appl. Ecol. 2006, 7, 201–214. [Google Scholar] [CrossRef]
- Klein, A.M.; Steffan–Dewenter, I.; Tscharntke, T. Fruit set of highland coffee increases with the diversity of pollinating bees. Proc. R. Soc. B. 2003, 270, 955–961. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Order | Family | Genus/Species | Total Abundance | Relative Proportion % | Foraging Task (N/P) |
|---|---|---|---|---|---|
| Hymenoptera | Apidae | Apis dorsata | 81 | 30.22 | N |
| Apis florea | 35 | 13.06 | N | ||
| Diptera | Syrphidae | Eristalinus aeneus Eupeodes corollae Episyrphus balteatus Eristalinus megacephalus Ischiodon scutellaris | 67 39 19 15 12 | 25.0 14.55 7.09 5.59 4.48 | N/P P P N/P N/P |
| Total syrphid flies | 152 | 56.71 | |||
| Total honey bees | 116 | 43.28 |
| Pollinator Species | Stay Time/Flower/Visit (N = 100) | Visitation Rate (No. of Flowers Visited/Min) (N = 100) |
|---|---|---|
| A. dorsata | 3.70 ± 0.19b | 9.69 ± 0.27a |
| A. florea | 14.46 ± 1.08a | 2.60 ± 0.22b |
| E. aeneus | 6.90 ± 1.02b | 7.66 ± 0.34a |
| E. corollae | 15.08 ± 9.83a | 2.50 ± 0.22b |
| Pollinator Species | Length (cm) | Pole (cm) | Equator (cm) | Weight (g) | Fruit Set (%) |
|---|---|---|---|---|---|
| A. dorsata | 22.23 ± 1.01a | 9.24 ± 2.02a | 7.11 ± 0.7a | 19.34 ± 1.67a | 80a |
| A. florea | 6.06 ± 0.78c | 7.88 ± 2.83a | 4.60 ± 0.92b | 8.37 ± 2.4b | 40c |
| E. aeneus | 12.57 ± 0.26b | 6.39 ± 0.84b | 4.06 ± 2.11b | 7.37 ± 1.28b | 60b |
| E. corollae | 4.02 ± 0.67c | 4.17 ± 2.94b | 1.89 ± 0.76c | 6.01 ± 0.43b | 30d |
| Results of Mann–Whitney U Test | Length (cm) | Pole (cm) | Equator (cm) | Weight (g) | Days to Reach Market Maturity | Fruit Set (%) |
|---|---|---|---|---|---|---|
| Mean of Ranks (open-pollination) | 32.745 | 28.745 | 28.138 | 31.809 | 8.871 | 22.25 |
| Mean of Ranks (self-pollination) | 14.755 | 18.755 | 19.362 | 15.691 | 22.629 | 18.25 |
| Mann–Whitney U | 306 | 682 | 739 | 394 | 54 | 640 |
| p-value | 0.0001 | 0.0011 | 0.0048 | 0.0001 | 0.0001 | 0.0253 |
| Pollination | TSS (Degree Brix) | TA (%) | Vitamin C | pH |
|---|---|---|---|---|
| Open-pollination | 7.14 ± 0.07 | 0.39 ± 0.03 | 220.6 ± 10.16 | 2.82 ± 0.11 |
| Self-pollination | 5.64 ± 0.25 | 0.29 ± 0.04 | 208.3 ± 13.17 | 3.79 ± 0.16 |
| Results of t-test | ||||
| t-observed | 5.622 | 2.023 | 0.740 | −4.718 |
| t-critical | 2.074 | 2.074 | 2.074 | 2.074 |
| p-value | <0.0001 | 0.055 | 0.467 | 0.000 |
| Df | 22 | 22 | 22 | 22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anees, M.; Ali, M.; Ghramh, H.A.; Sajjad, A.; Ali Khan, K.; Saeed, S.; Razzaq, K. Impact of Bee and Fly Pollination on Physical and Biochemical Properties of Strawberry Fruit. Horticulturae 2022, 8, 1072. https://doi.org/10.3390/horticulturae8111072
Anees M, Ali M, Ghramh HA, Sajjad A, Ali Khan K, Saeed S, Razzaq K. Impact of Bee and Fly Pollination on Physical and Biochemical Properties of Strawberry Fruit. Horticulturae. 2022; 8(11):1072. https://doi.org/10.3390/horticulturae8111072
Chicago/Turabian StyleAnees, Muhammad, Mudssar Ali, Hamed A. Ghramh, Asif Sajjad, Khalid Ali Khan, Shafqat Saeed, and Kashif Razzaq. 2022. "Impact of Bee and Fly Pollination on Physical and Biochemical Properties of Strawberry Fruit" Horticulturae 8, no. 11: 1072. https://doi.org/10.3390/horticulturae8111072
APA StyleAnees, M., Ali, M., Ghramh, H. A., Sajjad, A., Ali Khan, K., Saeed, S., & Razzaq, K. (2022). Impact of Bee and Fly Pollination on Physical and Biochemical Properties of Strawberry Fruit. Horticulturae, 8(11), 1072. https://doi.org/10.3390/horticulturae8111072