
Mineral Monitorization in Different Tissues of Solanum tuberosum L. during Calcium Biofortification Process

 ,
, 
 ,
,  , ,
, ,  ,
,  ,
,  ,
,  ,
,  , ,
, ,  ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Biofortification Itinerary
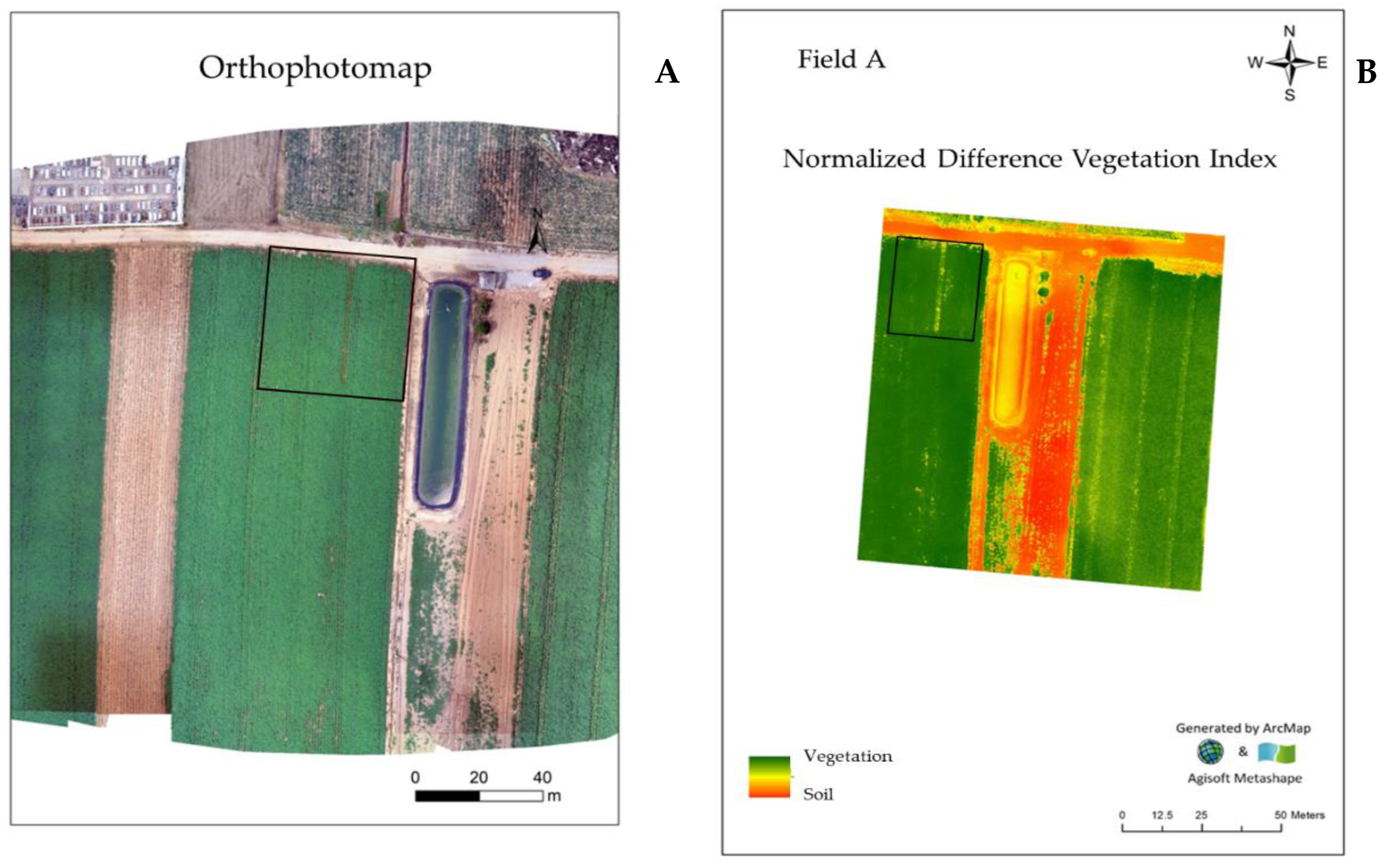
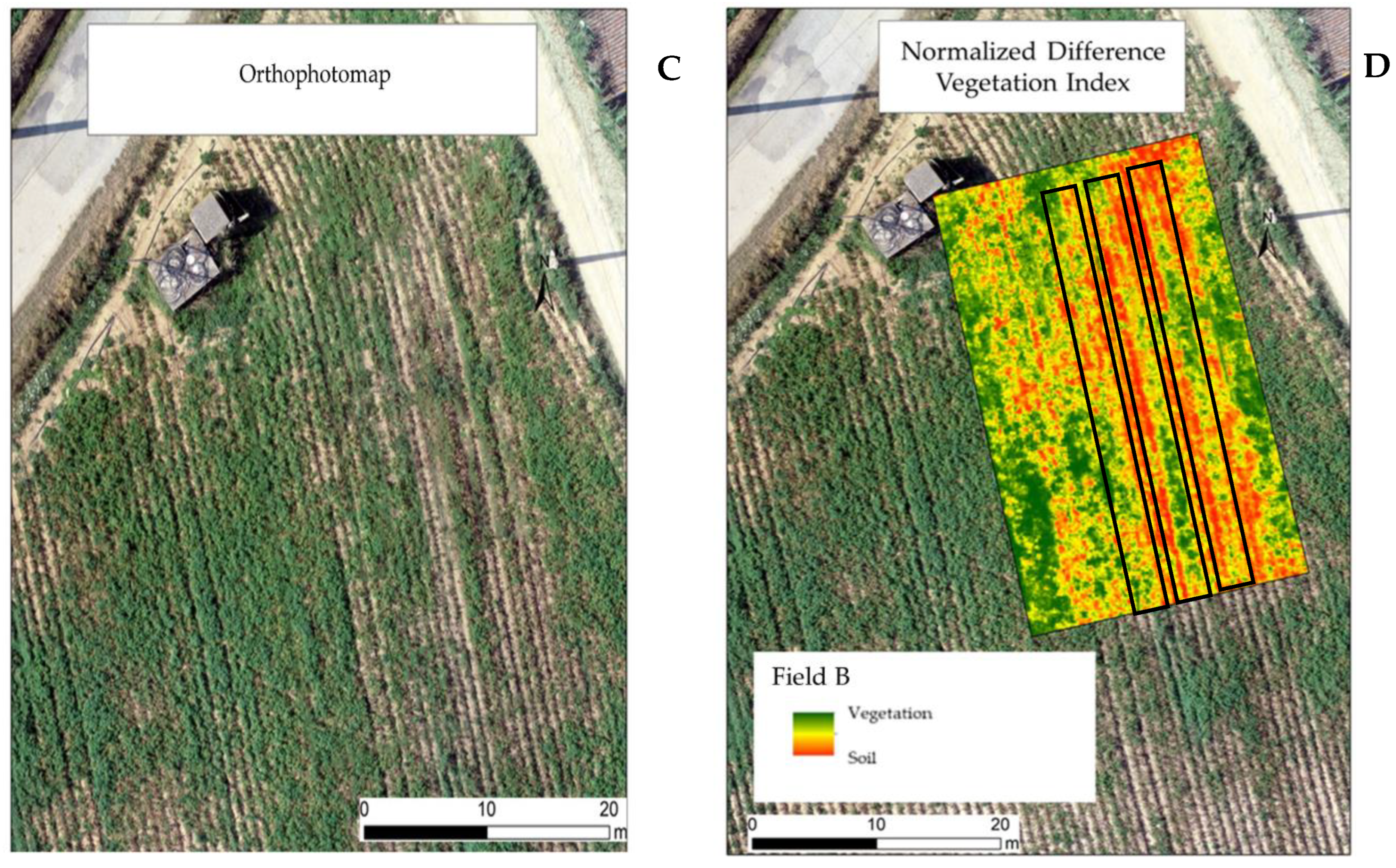
2.2. Ortophotomaps and NDVI (Normalized Difference Vegetation Index) Models in the Experimental Fields
2.3. Mineral Content in Roots, Stems, Leaves, and Tubers
2.4. Quality Parameters
2.5. Statistical Analysis
3. Results
3.1. Monitorization of Solanum tuberosum L. Plants
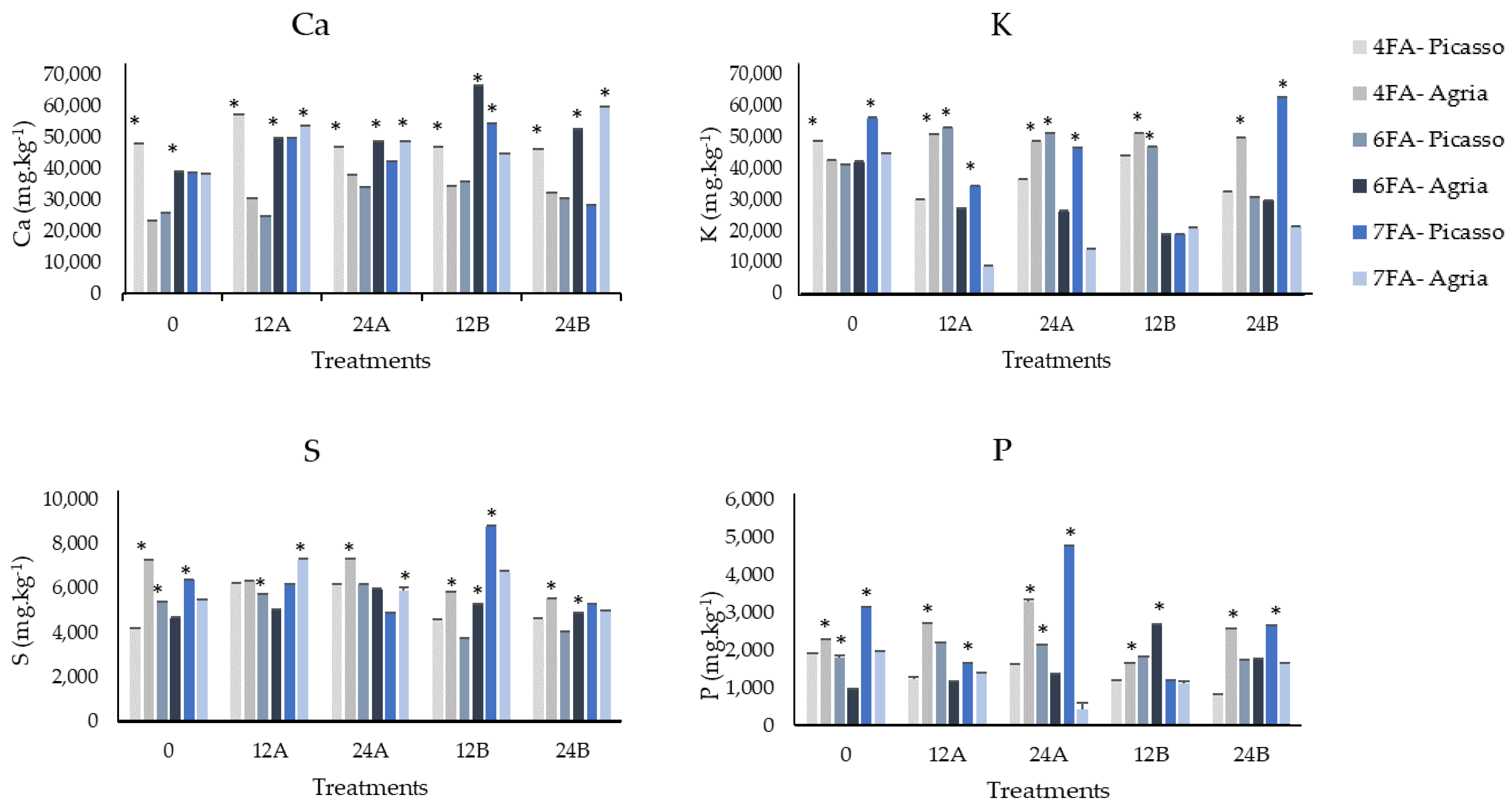
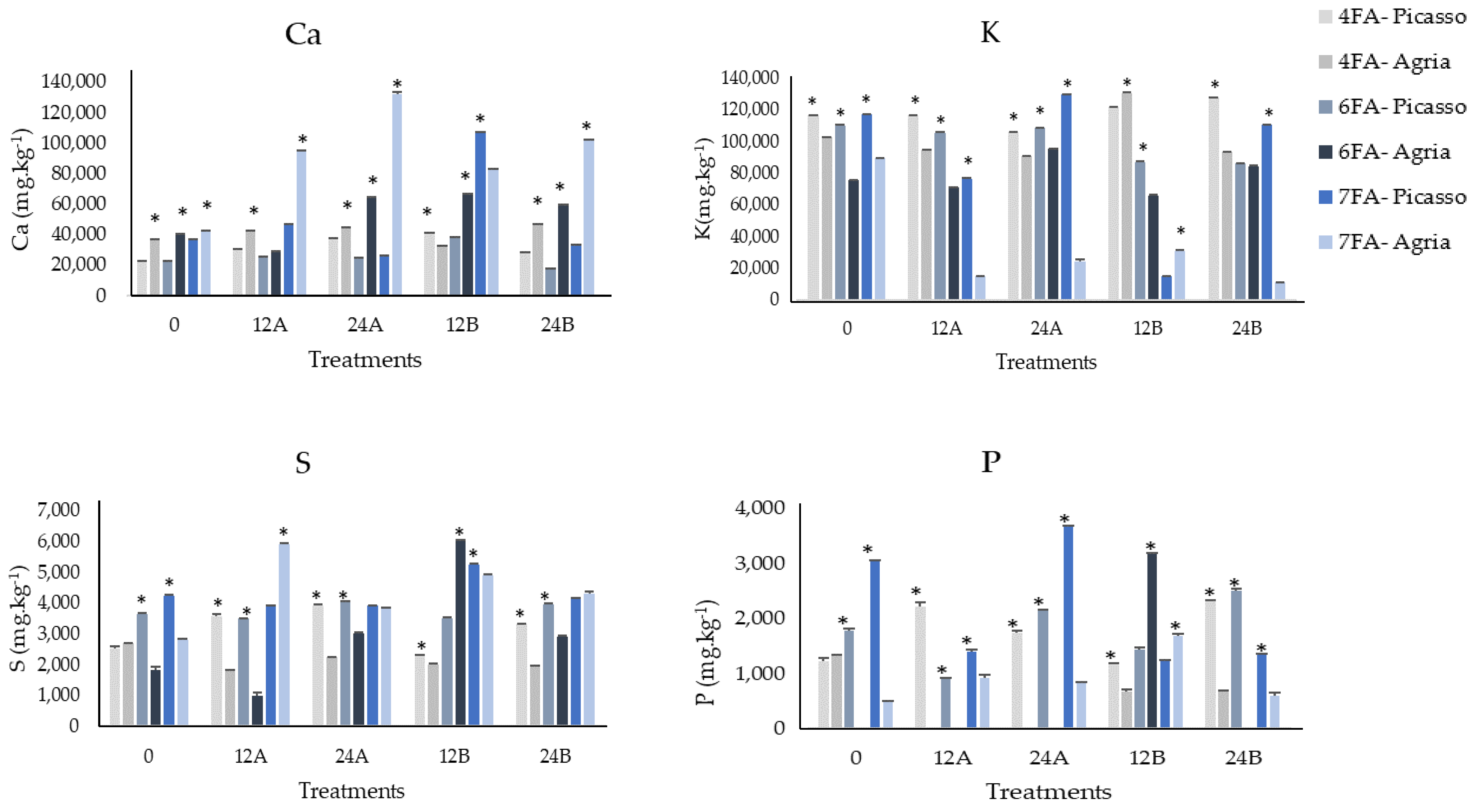
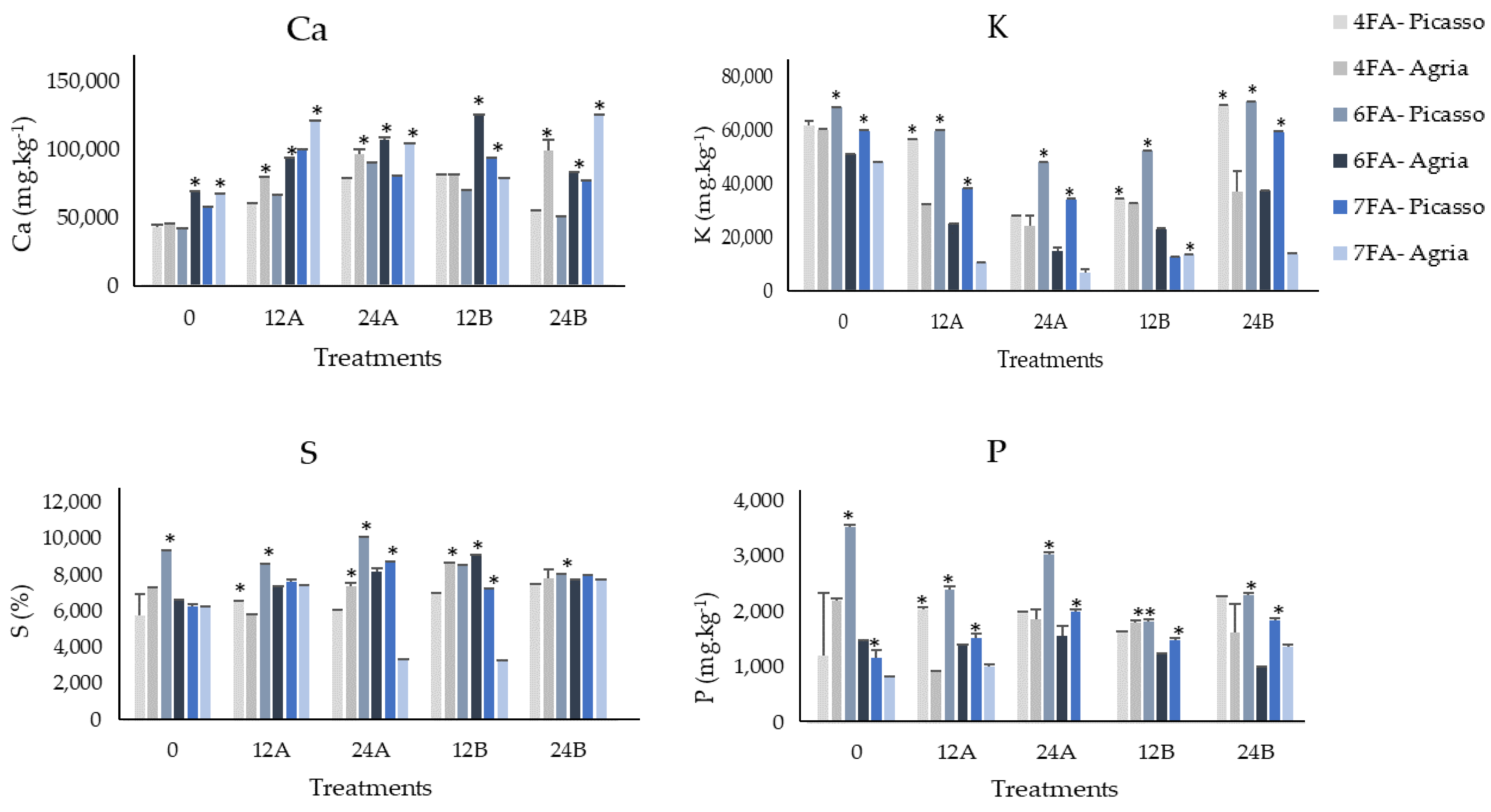
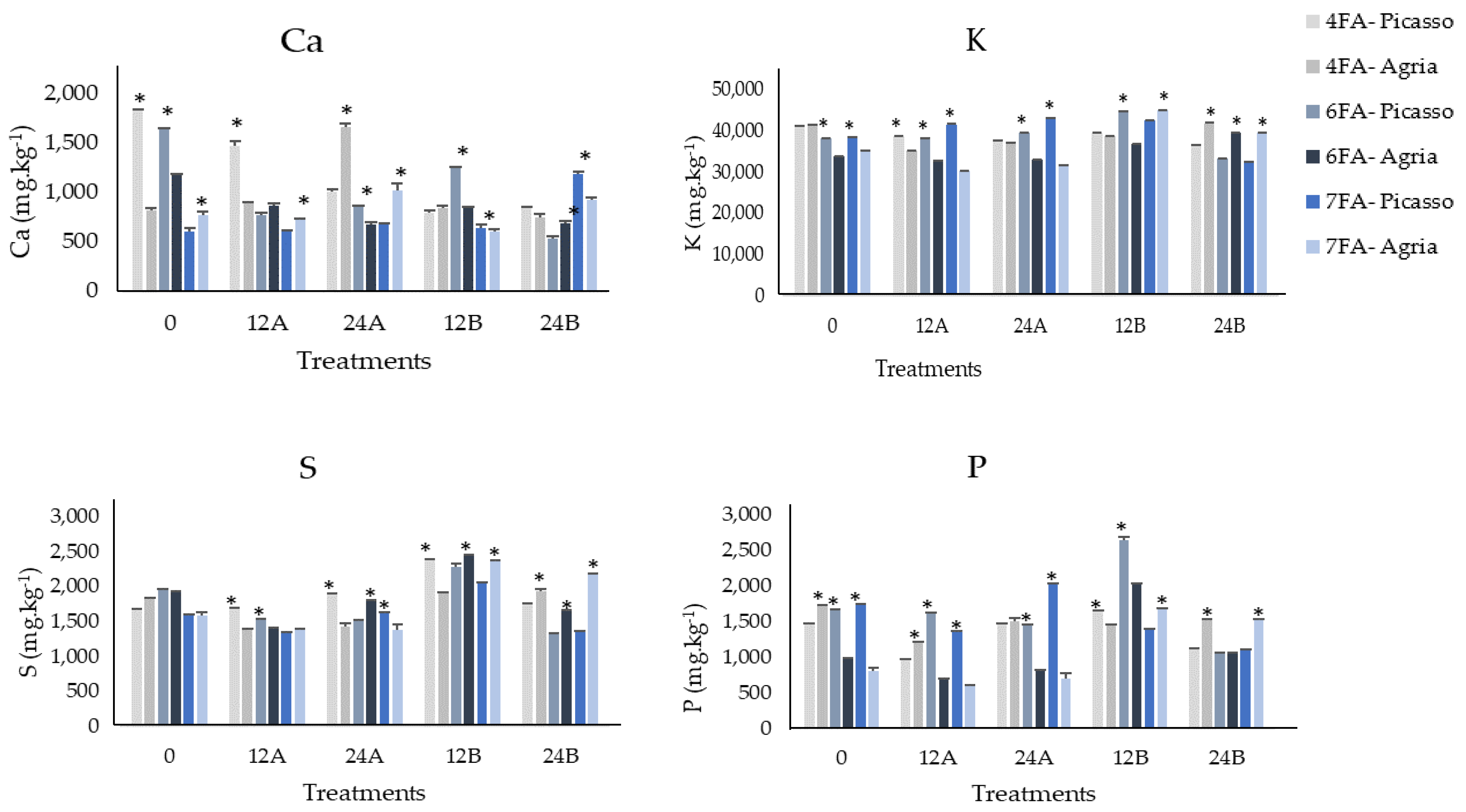
3.2. Mineral Content of Ca, K, S, and P after the Fourth, Sixth, and Seventh Foliar Applications
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Camire, M.E.; Kubow, S.; Donnelly, D.J. Potatoes and human health. Crit. Rev. Food Sci. Nutr. 2009, 49, 823–840. [Google Scholar] [CrossRef] [PubMed]
- FAO. Potatoes: So Familiar, So Much More to Learn. 2022. Available online: https://www.fao.org/publications/highlights-detail/en/c/1529406/ (accessed on 1 August 2022).
- Ledwożyw-Smoleń, I.; Smoleń, S.; Rożek, S.; Sady, W.; Strzetelski, P. Iodine biofortification of potato (Solanum tuberosum L.) grown in field. Agronomy 2020, 10, 1916. [Google Scholar] [CrossRef]
- Kromann, P.; Valverde, F.; Alvarado, S.; Vélez, R.; Pisuña, J.; Potosí, B.; Taipe, A.; Caballero, D.; Cabezas, A.; Devaux, A. Can Andean potatoes be agronomically biofortified with iron and zinc fertilizers? Plant Soil 2017, 411, 121–138. [Google Scholar] [CrossRef]
- Reboredo, F. Copper and zinc uptake by Halimione portulacoides (L.) Aellen. A long-term accumulation experiment. Bull. Environ. Contam. Toxicol. 1991, 46, 442–449. [Google Scholar] [CrossRef]
- Reboredo, F. The interaction between copper and zinc and their uptake by Halimione portulacoides (L.) Aellen. Bull. Environ. Contam. Toxicol. 1994, 52, 598–605. [Google Scholar] [CrossRef]
- Fageria, V.D. Nutrient interactions in crop plants. J. Plant Nutr. 2001, 24, 1269–1290. [Google Scholar] [CrossRef]
- Romera, F.J.; Lan, P.; Rodríguez-Celma, J.; Pérez-Vicente, R. Editorial: Nutrient Interactions in Plants. Front. Plant Sci. 2021, 12, 782505. [Google Scholar] [CrossRef]
- Smolen, S.; Rozek, S.; Ledwozyw-Smolen, I.; Strzetelski, P. Preliminary evaluation of the influence of soil fertilization and foliar nutrition with iodine on the efficiency of iodine biofortification and chemical composition of lettuce. J. Elem. 2011, 16, 613–622. [Google Scholar] [CrossRef]
- Sahin, O. Combined biofortification of soilless grown lettuce with iodine, selenium and zinc and its effect on essential and non-essential elemental composition. J. Plant Nutr. 2021, 44, 673–678. [Google Scholar] [CrossRef]
- Koç, E.; Karayiğit, B. Assessment of biofortification approaches used to improve micronutrient dense plants that are a sustainable solution to combat hidden hunger. J. Soil Sci. Plant Nutr. 2022, 22, 475–500. [Google Scholar] [CrossRef]
- Gunes, A.; Inal, A.; Kadioglu, Y.K. Determination of mineral element concentrations in wheat, sunflower, chickpea and lentil cultivars in response to P fertilization by polarized energy dispersive X-ray fluorescence. X-ray Spectrom. 2009, 38, 451–462. [Google Scholar] [CrossRef]
- Reboredo, F.; Lidon, F.J.C. UV-B radiation effects on terrestrial plants—A perspective. Emir. J. Food Agric. 2012, 24, 502–509. [Google Scholar] [CrossRef]
- Chen, Z.; Gao, W.; Reddy, K.R.; Chen, M.; Taduri, S.; Meyers, S.L.; Shankle, M.W. Ultraviolet (UV) B effects on growth and yield of three contrasting sweet potato cultivars. Photosynthetica 2020, 58, 37–44. [Google Scholar] [CrossRef]
- Ibañez, V.N.; Berli, F.J.; Masuelli, R.W.; Bottini, R.A.; Marfil, C.F. Influence of altitude and enhanced ultraviolet-B radiation on tuber production, seed viability, leaf pigments and morphology in the wild potato species Solanum kurtzianum Bitter & Wittm collected from an elevational gradient. Plant Sci. 2017, 261, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Semedo, J.N.; Rodrigues, A.P.; Lidon, F.C.; Pais, I.P.; Marques, I.; Gouveia, D.; Armengaud, J.; Silva, M.J.; Martins, S.; Semedo, M.C.; et al. Intrinsic non-stomatal resilience to drought of the photosynthetic apparatus in Coffea spp. is strengthened by elevated air [CO2]. Tree Physiol. 2020, 41, 708–727. [Google Scholar] [CrossRef]
- Davenport, J.R.; Milburn, P.H.; Rosen, C.J.; Thornton, R.E. Environmental impacts of potato nutrient management. Am. J. Potato Res. 2005, 82, 321–328. [Google Scholar] [CrossRef]
- Oliveira, R.C.; da Silva, J.R.R.; Lana, R.M.Q.; de Azevedo Pereira, A.I.; Castoldi, R.; de Camargo, R.; Luz, J.M.Q. Fertilizer appli-cation levels in potato crops and the Diagnosis and Recommendation Integrated System (DRIS). Agronomy 2021, 11, 51. [Google Scholar] [CrossRef]
- Westermann, D.T. Nutritional requirements of potatoes. Am. J. Potato Res. 2005, 82, 301–307. [Google Scholar] [CrossRef]
- Myvan, F.F.; Al-Ahmadi, M.J.; Eslami, S.V.; Noferest, K.S. Role of Potassium in modifying the potato physiological responses to irrigation regimes under different planting patterns. Potato Res. 2022, 65, 581–600. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, S.; Mohapatra, T. Interaction between macro-and micro-nutrients in plants. Front. Plant Sci. 2021, 12, 665583. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, B.; Horneck, D.; MacGuidwin, E. Improving phosphorus use efficiency through potato rhizosphere modification and extension. Am. J. Potato Res. 2014, 91, 161–174. [Google Scholar] [CrossRef]
- Leonel, M.; do Carmo, E.L.; Fernandes, A.M.; Soratto, R.P.; Ebúrneo, J.A.M.; Garcia, É.L.; Dos Santos, T.P.R. Chemical composition of potato tubers: The effect of cultivars and growth conditions. J. Food Sci. Technol. 2017, 54, 2372–2378. [Google Scholar] [CrossRef] [PubMed]
- Narayan, O.P.; Kumar, P.; Yadav, B.; Dua, M.; Johri, A.K. Sulfur nutrition and its role in plant growth and development. Plant Signal. Behav. 2022, 2030082. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.L.; Kushwah, S.S.; Nema, P.K.; Rathore, S.S. Effect of sulphur on yield and quality of potato (Solanum tuberosum L.). Int. J. Agric. Res. 2011, 6, 143–148. [Google Scholar] [CrossRef]
- Barczak, B.; Nowak, K.; Knapowski, T. Potato yield is affected by sulphur form and rate. Agrochimica 2013, 57, 363–372. [Google Scholar]
- Thor, K. Calcium—Nutrient and Messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef]
- Pilbeam, D.J.; Morley, P.S. Calcium. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; CRC Press: Boca Raton, FL, USA, 2, 2016; pp. 137–160. [Google Scholar]
- Abbasian, A.; Ahmadi, A.; Abbasi, A.-R.; Darvishi, B. Effect of various phosphorus and calcium concentrations on potato seed tuber production. J. Plant Nutr. 2018, 41, 1765–1777. [Google Scholar] [CrossRef]
- Luís, I.C.; Lidon, F.C.; Pessoa, C.C.; Marques, A.C.; Coelho, A.R.F.; Simões, M.; Patanita, M.; Dôres, J.; Ramalho, J.C.; Silva, M.M.; et al. Zinc Enrichment in Two Contrasting Genotypes of Triticum aestivum L. Grains: Interactions between Edaphic Conditions and Foliar Fertilizers. Plants 2021, 10, 204. [Google Scholar] [CrossRef]
- Coelho, A.R.F.; Lidon, F.C.; Pessoa, C.C.; Marques, A.C.; Luís, I.C.; Caleiro, J.; Simões, M.; Kullberg, J.; Legoinha, P.; Brito, M.; et al. Can foliar pulverization with CaCl2 and Ca(NO3)2 trigger ca enrichment in Solanum tuberosum L. tubers? Plants 2021, 10, 245. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, N.K.; White, P.J.; Broadley, M.R.; Ramsay, G. The three-dimensional distribution of minerals in potato tubers. Ann. Bot. 2011, 107, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Winkelmann, M.K.; Hasler, M.; Pawelzik, E.; Naumann, M. Root growth in light of changing magnesium distribution and transport between source and sink tissues in potato (Solanum tuberosum L.). Sci. Rep. 2020, 10, 8796. [Google Scholar] [CrossRef] [PubMed]
- El-Zohiri, S.S.M.; Asfour, H.E. Effects of foliar sprays of potassium, magnesium and calcium on yield, quality and storageability of potato. In Proceedings of the Fifth International Congress of Sustainable Agriculture Development, Fayoum, Egypt, 21–23 December 2009; Faculty of Agriculture, Fayoum University: Fayoum, Egypt, 2009; pp. 57–71. [Google Scholar]
- El-Hadidi, E.M.; El-Dissoky, R.A.; Abdelhafez, A.A.H. Foliar calcium and magnesium application effect on potato crop grown in clay loam soils. J. Soil Sci. Agric. Eng. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Seifu, Y.W.; Deneke, S. Effect of calcium chloride and calcium nitrate on potato (Solanum tuberosum L.) growth and yield. J. Hortic. 2017, 4, 1000207. [Google Scholar]
- Mousavi, S.; Galavi, M.; Ahmadvand, G. Effect of zinc and manganese foliar application on yield, quality and enrichment on potato (Solanum tuberosum L.). Asian J. Plant Sci. 2007, 6, 1256–1260. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R.; Hammond, J.P.; Ramsay, G.; Subramanian, N.K.; Thompson, J.; Wright, G. Biofortification of potato tubers using foliar zinc-fertilisers. J. Hortic. Sci. Biotechnol. 2012, 87, 123–129. [Google Scholar] [CrossRef]
- White, P.J.; Thompson, J.A.; Wright, G.; Rasmussen, S.K. Biofortifying Scottish potatoes with zinc. Plant Soil 2017, 411, 151–165. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Z.; Zhang, X.; Zhang, W.; Huang, L.; Zhang, Z.; Yuan, L.; Liu, X. Effects of foliar application of selenate and selenite at different growth stages on Selenium accumulation and speciation in potato (Solanum tuberosum L.). Food Chem. 2019, 286, 550–556. [Google Scholar] [CrossRef]
- Poggi, V.; Arcioni, A.; Filippini, P.; Pifferi, P. Foliar application of selenite and selenate to potato (Solanum tuberosum): Effect of a ligand agent on selenium content of tubers. J. Agric. Food Chem. 2000, 48, 4749–4751. [Google Scholar] [CrossRef]
- Oliveira, V.; Faquin, V.; Andrade, F.; Carneiro, J.; Júnior, E.; Souza, K.; Pereira, J.; Guilherme, L. Physiological and physicochemical responses of potato to selenium biofortification in tropical soil. Potato Res. 2019, 62, 315–331. [Google Scholar] [CrossRef]
- Almekinders, C.J.M.; Struik, P.C. Shoot development and flowering in potato (Solanum tuberosum L.). Potato Res. 1996, 39, 581–607. [Google Scholar] [CrossRef]
- de Souza Alonso, T.A.; Ferreira Barreto, R.; de Mello Prado, R.; Pereira de Souza, J.; Falleiros Carvalho, R. Silicon spraying alleviates calcium deficiency in tomato plants, but Ca-EDTA is toxic. J. Plant Nutr. Soil Sci. 2020, 183, 659–664. [Google Scholar] [CrossRef]
- Hütsch, B.W.; Keipp, K.; Glaser, A.K.; Schubert, S. Potato plants (Solanum tuberosum L.) are chloride-sensitive: Is this dogma valid? J. Sci. Food Agric. 2018, 98, 3161–3168. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Naumann, M.; Pawelzik, E.; Gransee, A.; Thiel, H. The importance of nutrient management for potato production Part I: Plant nutrition and yield. Potato Res. 2020, 63, 97–119. [Google Scholar] [CrossRef]
- Abdel-Salam, M.A.; Shams, A.S. Feldspar-K fertilization of potato (Solanum tuberosum L.) augmented by biofertilizer. J. Agric. Environ. Sci. 2012, 12, 694–699. [Google Scholar] [CrossRef]
- Ewais, M.A.; Abd El-Rahman, L.A.; Sayed, D.A. Effect of foliar application of boron and potassium sources on yield and quality of potato (Solanum tuberosum L.). Middle East J. Appl. Sci. 2020, 10, 120–137. [Google Scholar]
- Sharma, U.C.; Arora, B.R. Critical nutrient ranges for potassium in potato leaves and petioles. J. Hortic. Sci. 1989, 64, 47–51. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, H.; Fan, J.; Zhang, F.; Cheng, M.; Yang, L.; Ji, Q.; Li, Z. Quantifying source-sink relationships of drip-fertigated potato under various water and potassium supplies. Field Crops Res. 2022, 285, 108604. [Google Scholar] [CrossRef]
- Karley, A.J.; White, P.J. Moving cationic minerals to edible tissues: Potassium, magnesium, calcium. Curr. Opin. Plant Biol. 2009, 12, 291–298. [Google Scholar] [CrossRef]
- Raghothama, K.G.; Karthikeyan, A.S. Phosphate acquisition. Plant Soil 2005, 274, 37–49. [Google Scholar] [CrossRef]
- Potarzycki, J.; Grzebisz, W. Trends in phosphorus concentrations in potato organs during the growing season. J. Elem. 2019, 24, 935–952. [Google Scholar] [CrossRef]
- White, P.J.; Bradshaw, J.E.; Brown, L.K.; Dale, M.F.B.; Dupuy, L.X.; George, T.S.; Hammond, J.P.; Subramanian, N.K.; Thompson, J.A.; Wishart, J.; et al. Juvenile root vigour improves phosphorus use efficiency of potato. Plant Soil 2018, 432, 45–63. [Google Scholar] [CrossRef]
- Grzebisz, W.; Frąckowiak, K.; Spiżewski, T.; Przygocka-Cyna, K. Does Elemental Sulfur Act as an Effective Measure to Control the Seasonal Growth Dynamics of Potato Tubers (Solanum tuberosum L.)? Plants 2022, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Mineral Nutrition and Yield Response. In Mineral Nutrition of Higher Plants; Academic Press: London, UK, 1995; pp. 184–200. [Google Scholar]
- Navarre, D.A.; Goyer, A.; Shakya, R. Nutritional value of potatoes: Vitamin, phytonutrient, and mineral content. In Advances in Potato Chemistry and Technology; Academic Press: Cambridge, MA, USA, 2009; pp. 395–424. [Google Scholar] [CrossRef]
- Braun, H.; Fontes, P.; Finger, F.; Busato, C.; Cecon, P. Carboidratos e matéria seca de tubérculos de cultivares de batata influenciados por doses de nitrogênio. Cienc. Agrotec. 2010, 34, 285–293. [Google Scholar] [CrossRef]
- Portaria n.° 587/87 de 9 de Julho. Diário da República n.o 155/1987, Série, I. Ao Abrigo do Disposto no n.o1 do Artigo 4.o do Decreto-Lei n.o 512/85, de 31 de Dezembro; Ministérios da Agricultura, Pescas e Alimentação e da Indústria e Comércio: Lisboa, Portugal, 2021.
- Cohen, J.; Cohen, P.; West, S.G.; Aiken, L.S. Applied Multiple Correlation/Regression Analysis for the Behavioral Sciences; Lawrence Erlbaum Associates: Mahwah, NJ, USA, 2003. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Foliar Application | Treatments | Picasso | Agria | ||||
|---|---|---|---|---|---|---|---|
| Height (cm) | Diameter (cm) | Dry Weight (%) | Height (cm) | Diameter (cm) | Dry Weight (%) | ||
| 4° | Control | 8.30 ± 2.08a * | 3.53 ± 0.78a | 18.40 ± 1.11a | 2.33 ± 0.88b | 3.00 ± 1.15a | 16.75 ± 2.40a |
| CaCl2 12 kg∙ha−1 | 9.27 ± 1.49a | 6.17 ± 1.17a | 15.90 ± 0.70a | 8.73 ± 0.22a | 6.03 ± 0.37a | 21.24 ± 0.76a * | |
| CaCl2 24 kg∙ha−1 | 7.00 ± 0.65a * | 7.30 ± 1.15a * | 15.08 ± 0.98a | 4.80 ± 1.75ab | 3.70 ± 1.26a | 16.90 ± 3.10a | |
| Ca-EDTA 12 kg∙ha−1 | 5.20 ± 1.30a | 5.40 ± 0.57a * | 18.04 ± 2.45a | 4.17 ± 0.46ab | 3.47 ± 0.29a | 18.86 ± 0.34a | |
| Ca-EDTA 24 kg∙ha−1 | 6.80 ± 0.40a * | 4.53 ± 1.12a | 14.98 ± 1.57a | 4.63 ± 0.88ab | 3.53 ± 0.78a | 26.46 ± 10.5a | |
| 6° | Control | 6.63 ± 2.19a | 4.67 ± 1.20a | 14.86 ± 1.08a | 4.77 ± 0.39a | 3.67 ± 0.18a | 21.47 ± 0.63a * |
| CaCl2 12 kg∙ha−1 | 9.20 ± 1.68a | 7.03 ± 1.21a | 16.61 ± 0.67a | 7.50 ± 1.00a | 6.00 ± 0.40a | 23.13 ± 0.49a * | |
| CaCl2 24 kg∙ha−1 | 6.57 ± 2.13a | 4.73 ± 1.28a | 14.47 ± 1.62a | 7.77 ± 0.38a | 5.73 ± 0.20a | 28.99 ± 5.58a * | |
| Ca-EDTA 12 kg∙ha−1 | 6.90 ± 0.21a | 5.73 ± 0.52a | 16.02 ± 0.20a | 7.00 ± 2.17a | 5.53 ± 0.69a | 19.32 ± 0.34a * | |
| Ca-EDTA 24 kg∙ha−1 | 9.07 ± 2.17a | 5.40 ± 1.07a | 15.38 ± 0.30a | 6.37 ± 0.71a | 4.87 ± 0.69a | 18.08 ± 3.10a | |
| 7° | Control | 13.6 ± 0.60a * | 6.30 ± 0.80a | 17.13 ± 0.93a | 7.43 ± 0.55a | 5.40 ± 0.53a | 23.67 ± 0.43a * |
| CaCl2 12 kg∙ha−1 | 8.70 ± 1.11a | 4.40 ± 0.31a | 16.57 ± 0.78a | 10.43 ± 0.09a * | 7.67 ± 0.12a | 23.13 ± 0.78a * | |
| CaCl2 24 kg∙ha−1 | 9.27 ± 0.37a * | 5.50 ± 0.52a | 16.21 ± 1.04a | 7.53 ± 1.06a | 5.83 ± 0.77a | 20.83 ± 0.56a * | |
| Ca-EDTA 12 kg∙ha−1 | 7.90 ± 0.61a | 5.10 ± 0.21a | 15.84 ± 0.36a | 7.20 ± 1.90a | 6.40 ± 1.40a | 23.46 ± 0.19a * | |
| Ca-EDTA 24 kg∙ha−1 | 8.20 ± 1.52a * | 4.97 ± 0.71a | 15.22 ± 0.43a | 5.20 ± 1.30a | 3.80 ± 0.90a | 22.40 ± 0.99a * | |
| Ca | K | S | P | H | D | Dw | |
|---|---|---|---|---|---|---|---|
| Ca | 1 | −0.866 | −0.398 | −0.579 | 0.045 | 0.012 | 0.131 |
| K | −0.866 | 1 | 0.515 | 0.603 | −0.040 | 0.022 | 0.045 |
| S | −0.398 | 0.515 | 1 | 0.226 | 0.269 | −0.087 | 0.081 |
| P | −0.579 | 0.603 | 0.226 | 1 | 0.245 | 0.125 | 0.059 |
| H | 0.045 | −0.040 | 0.269 | 0.245 | 1 | −0.056 | −0.039 |
| D | 0.012 | 0.022 | −0.087 | 0.125 | −0.056 | 1 | 0.817 |
| Dw | 0.131 | 0.045 | 0.081 | 0.059 | −0.039 | 0.817 | 1 |
| Ca | K | S | P | H | D | Dw | |
|---|---|---|---|---|---|---|---|
| Ca | 1 | −0.360 | −0.316 | −0.251 | −0.651 | −0.123 | −0.129 |
| K | −0.360 | 1 | 0.970 | 0.955 | 0.273 | −0.637 | −0.532 |
| S | −0.316 | 0.970 | 1 | 0.994 | 0.227 | −0.662 | −0.561 |
| P | −0.251 | 0.955 | 0.994 | 1 | 0.162 | −0.685 | −0.585 |
| H | −0.651 | 0.273 | 0.227 | 0.162 | 1 | −0.144 | −0.156 |
| D | −0.123 | −0.637 | −0.662 | −0.685 | −0.144 | 1 | 0.974 |
| Dw | −0.129 | −0.521 | −0.561 | −0.585 | −0.156 | 0.974 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coelho, A.R.F.; Lidon, F.C.; Pessoa, C.C.; Daccak, D.; Luís, I.C.; Marques, A.C.; Ramalho, J.C.; Semedo, J.M.N.; Silva, M.M.; Pais, I.P.; et al. Mineral Monitorization in Different Tissues of Solanum tuberosum L. during Calcium Biofortification Process. Horticulturae 2022, 8, 1020. https://doi.org/10.3390/horticulturae8111020
Coelho ARF, Lidon FC, Pessoa CC, Daccak D, Luís IC, Marques AC, Ramalho JC, Semedo JMN, Silva MM, Pais IP, et al. Mineral Monitorization in Different Tissues of Solanum tuberosum L. during Calcium Biofortification Process. Horticulturae. 2022; 8(11):1020. https://doi.org/10.3390/horticulturae8111020
Chicago/Turabian StyleCoelho, Ana Rita F., Fernando Cebola Lidon, Cláudia Campos Pessoa, Diana Daccak, Inês Carmo Luís, Ana Coelho Marques, José Cochicho Ramalho, José Manuel N. Semedo, Maria Manuela Silva, Isabel P. Pais, and et al. 2022. "Mineral Monitorization in Different Tissues of Solanum tuberosum L. during Calcium Biofortification Process" Horticulturae 8, no. 11: 1020. https://doi.org/10.3390/horticulturae8111020
APA StyleCoelho, A. R. F., Lidon, F. C., Pessoa, C. C., Daccak, D., Luís, I. C., Marques, A. C., Ramalho, J. C., Semedo, J. M. N., Silva, M. M., Pais, I. P., Brito, M. G., Kullberg, J. C., Legoinha, P., Simões, M., Scotti-Campos, P., Pessoa, M. F., & Reboredo, F. H. (2022). Mineral Monitorization in Different Tissues of Solanum tuberosum L. during Calcium Biofortification Process. Horticulturae, 8(11), 1020. https://doi.org/10.3390/horticulturae8111020