
Genetic Architecture of Chile Pepper (Capsicum spp.) QTLome Revealed Using Meta-QTL Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Characterization of QTL for Different Traits in Chile Peppers
2.2. Projection to a Consensus Map and Analysis of Meta-QTL
2.3. Candidate Gene Identification for the Meta-QTL
3. Results
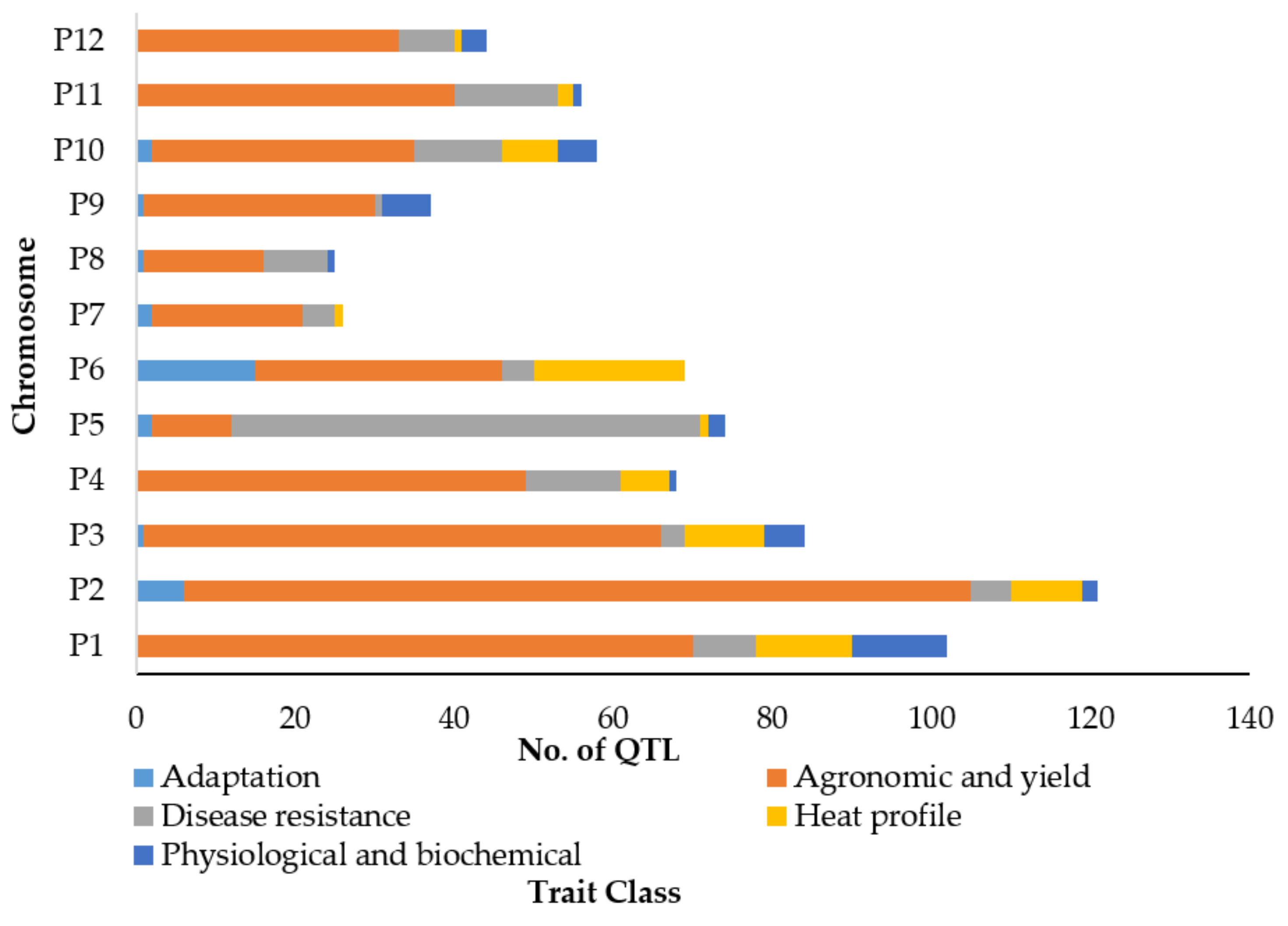
3.1. Classification of QTL for Diverse Traits in Capsicum spp.
3.2. Meta-QTL Analysis
3.3. Candidate Gene Mining
4. Discussion
4.1. Meta-QTL Regions Were Identified for Capsicum QTLome for Diverse Sets of Traits
4.2. Two Meta-QTL Regions Related with P. capsici Resistance Were Delimited to <1.0 cM
4.3. Epigenetic Mechanisms Could Play a Role in Conferring Resistance to Chile Pepper Blight
4.4. Colocalization of QTL Related with Different Traits Indicates Potential of Multi-Trait Genetic Improvement in Capsicum
4.5. From Meta-QTL to Marker-Assisted Selection for Disease Resistance in Chile Peppers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kantar, M.B.; Anderson, J.E.; Lucht, S.A.; Mercer, K.; Bernau, V.; Case, K.A.; Le, N.C.; Frederiksen, M.K.; DeKeyser, H.C.; Wong, Z.-Z. Vitamin variation in Capsicum spp. provides opportunities to improve nutritional value of human diets. PLoS ONE 2016, 11, e0161464. [Google Scholar] [CrossRef]
- Guzmán, I.; Bosland, P.W. Sensory properties of chile pepper heat–and its importance to food quality and cultural preference. Appetite 2017, 117, 186–190. [Google Scholar] [CrossRef]
- Lozada, D.N.; Coon, D.L.; Guzmán, I.; Bosland, P.W. Heat profiles of ‘superhot’and New Mexican type chile peppers (Capsicum spp.). Sci. Hortic. 2021, 283, 110088. [Google Scholar] [CrossRef]
- Palevitch, D.; Craker, L.E. Nutritional and medical importance of red pepper (Capsicum spp.). J. Herbs Spices Med. Plants 1996, 3, 55–83. [Google Scholar] [CrossRef]
- Bosland, P.W.; Walker, S.J. Growing chiles in New Mexico. New Mexico State Univ. Coop. Ext. Serv. Guid. H-230 2004, 1–8. [Google Scholar]
- Kenyon, L.; Kumar, S.; Tsai, W.S.; Hughes, J.d.A. Virus Diseases of peppers (Capsicum spp.) and their control. Adv. Virus Res. 2014, 90, 297–354. [Google Scholar]
- Moulin, M.M.; Rodrigues, R.; Bento, C.S.; Gonçalves, L.; Santos, J.O.; Sudré, C.P.; Viana, A.P. Genetic dissection of agronomic traits in capsicum baccatum var. Pendulum. Genet. Mol. Res. 2015, 14, 2122–2132. [Google Scholar] [CrossRef]
- Pinto, C.M.F.; dos Santos, I.C.; de Araujo, F.F.; da Silva, T.P. Pepper importance and growth (Capsicum spp.). In Production and Breeding of Chilli Peppers (Capsicum Spp.); Springer: Cham, Switzerland, 2016; ISBN 978-3-319-06532-8. [Google Scholar]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Salvi, S.; Tuberosa, R. The crop QTLome comes of age. Curr. Opin. Biotechnol. 2015, 32, 179–185. [Google Scholar] [CrossRef]
- Martinez, A.K.; Soriano, J.M.; Tuberosa, R.; Koumproglou, R.; Jahrmann, T.; Salvi, S. Yield QTLome distribution correlates with gene density in maize. Plant Sci. 2016, 242, 300–309. [Google Scholar] [CrossRef]
- Goffinet, B.; Gerber, S. Quantitative trait loci: A meta-analysis. Genetics 2000, 155, 463–473. [Google Scholar] [CrossRef]
- Acuña-Galindo, M.A.; Mason, R.E.; Subramanian, N.K.; Hays, D.B. Meta-analysis of wheat QTL regions associated with adaptation to drought and heat stress. Crop Sci. 2015, 55, 477–492. [Google Scholar] [CrossRef]
- Löffler, M.; Schön, C.-C.; Miedaner, T. Revealing the genetic architecture of FHB resistance in hexaploid wheat (Triticum aestivum L.) by QTL meta-analysis. Mol. Breed. 2009, 23, 473–488. [Google Scholar] [CrossRef]
- Danan, S.; Veyrieras, J.-B.; Lefebvre, V. Construction of a potato consensus map and QTL meta-analysis offer new insights into the genetic architecture of late blight resistance and plant maturity traits. BMC Plant Biol. 2011, 11, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.; Ontoy, J.; Subudhi, P.K. Meta-analysis of quantitative trait loci associated with seedling-stage salt tolerance in rice (Oryza sativa L.). Plants 2019, 8, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, I.S.; Nadarajah, K. A Meta-Analysis of Quantitative Trait Loci Associated with Multiple Disease Resistance in Rice (Oryza sativa L.). Plants 2020, 9, 1491. [Google Scholar] [CrossRef]
- Emebiri, L.; Singh, S.; Tan, M.-K.; Singh, P.K.; Fuentes-Dávila, G.; Ogbonnaya, F. Unravelling the Complex Genetics of Karnal Bunt (Tilletia indica) Resistance in Common Wheat (Triticum aestivum) by Genetic Linkage and Genome-Wide Association Analyses. G3 Genes Genomes Genet. 2019, 9, 1437–1447. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Saripalli, G.; Jan, I.; Kumar, K.; Sharma, P.K.; Balyan, H.S.; Gupta, P.K. Meta-QTL analysis and identification of candidate genes for drought tolerance in bread wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2020, 26, 1713–1725. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shabala, S.; Koutoulis, A.; Shabala, L.; Zhou, M. Meta-analysis of major QTL for abiotic stress tolerance in barley and implications for barley breeding. Planta 2017, 245, 283–295. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Z.; Deng, D.; Ding, H.; Zhang, R.; Wang, S.; Bian, Y.; Yin, Z.; Xu, X. Meta-analysis combined with syntenic metaQTL mining dissects candidate loci for maize yield. Mol. Breed. 2013, 31, 601–614. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, G.; Tan, S.; Li, D.; Weiß, T.M.; Wang, X.; Chen, S.; Würschum, T.; Liu, W. A QTL atlas for grain yield and its component traits in maize (Zea mays). Plant Breed. 2020, 139, 562–574. [Google Scholar] [CrossRef]
- Said, J.I.; Song, M.; Wang, H.; Lin, Z.; Zhang, X.; Fang, D.D.; Zhang, J. A comparative meta-analysis of QTL between intraspecific Gossypium hirsutum and interspecific G. hirsutum × G. barbadense populations. Mol. Genet. Genom. 2015. [Google Scholar] [CrossRef] [PubMed]
- Van, K.; McHale, L.K. Meta-analyses of QTLs associated with protein and oil contents and compositions in soybean [Glycine max (L.) Merr.] seed. Int. J. Mol. Sci. 2017, 18, 1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallard, S.; Cantet, M.; Massire, A.; Bachellez, A.; Ewert, S.; Lefebvre, V. A key QTL cluster is conserved among accessions and exhibits broad-spectrum resistance to Phytophthora capsici: A valuable locus for pepper breeding. Mol. Breed. 2013, 32, 349–364. [Google Scholar] [CrossRef]
- Cheng, J.; Qin, C.; Tang, X.; Zhou, H.; Hu, Y.; Zhao, Z.; Cui, J.; Li, B.; Wu, Z.; Yu, J. Development of a SNP array and its application to genetic mapping and diversity assessment in pepper (Capsicum spp.). Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Zhu, Z.; Sun, B.; Wei, J.; Cai, W.; Huang, Z.; Chen, C.; Cao, B.; Chen, G.; Lei, J. Construction of a high density genetic map of an interspecific cross of Capsicum chinense and Capsicum annuum and QTL analysis of floral traits. Sci. Rep. 2019, 9, 1054. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.R.; Yoon, J.B.; Lee, J. A SNP-based genetic linkage map of Capsicum baccatum and its comparison to the Capsicum annuum reference physical map. Mol. Breed. 2016, 36, 61. [Google Scholar] [CrossRef]
- Endelman, J.B.; Plomion, C. LPmerge: An R package for merging genetic maps by linear programming. Bioinformatics 2014, 30, 1623–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 17 November 2020).
- Darvasi, A.; Soller, M. A simple method to calculate resolving power and confidence interval of QTL map location. Behav. Genet. 1997, 27, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Sleper, D.A.; Lu, P.; Shannon, J.G.; Nguyen, H.T.; Arelli, P.R. QTLs associated with resistance to soybean cyst nematode in soybean: Meta-analysis of QTL locations. Crop Sci. 2006, 46, 595–602. [Google Scholar] [CrossRef]
- Arcade, A.; Labourdette, A.; Falque, M.; Mangin, B.; Chardon, F.; Charcosset, A.; Joets, J. BioMercator: Integrating genetic maps and QTL towards discovery of candidate genes. Bioinformatics 2004, 20, 2324–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venske, E.; Dos Santos, R.S.; Farias, D.d.R.; Rother, V.; da Maia, L.C.; Pegoraro, C.; Costa de Oliveira, A. Meta-analysis of the QTLome of Fusarium head blight resistance in bread wheat: Refining the current puzzle. Front. Plant Sci. 2019, 10, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouellette, L.A.; Reid, R.W.; Blanchard, S.G.; Brouwer, C.R. LinkageMapView—Rendering high-resolution linkage and QTL maps. Bioinformatics 2018, 34, 306–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolser, D.; Staines, D.M.; Pritchard, E.; Kersey, P. Ensembl plants: Integrating tools for visualizing, mining, and analyzing plant genomics data. In Plant Bioinformatics; Springer: Berlin/Heidelberg, Germany, 2016; pp. 115–140. [Google Scholar]
- Alimi, N.A.; Bink, M.; Dieleman, J.A.; Nicolaï, M.; Wubs, M.; Heuvelink, E.; Magan, J.; Voorrips, R.E.; Jansen, J.; Rodrigues, P.C. Genetic and QTL analyses of yield and a set of physiological traits in pepper. Euphytica 2013, 190, 181–201. [Google Scholar] [CrossRef]
- Arjun, K.; Dhaliwal, M.S.; Jindal, S.K.; Fakrudin, B. Mapping of fruit length related QTLs in interspecific cross (Capsicum annuum L. × Capsicum galapagoense Hunz.) of chilli. Breed. Sci. 2018, 68, 17073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chunthawodtiporn, J.; Hill, T.; Stoffel, K.; Van Deynze, A. Quantitative trait loci controlling fruit size and other horticultural traits in bell pepper (Capsicum annuum). Plant Genome 2018, 11, 160125. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Wen, C.; Zhang, X.; Xu, X.; Yang, J.; Chen, B.; Geng, S. Identification of a major QTL (qRRs-10.1) that confers resistance to Ralstonia solanacearum in pepper (Capsicum annuum) using SLAF-BSA and QTL mapping. Int. J. Mol. Sci. 2019, 20, 5887. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, N.; Kumar, R.; Paliwal, R.; Kumar, U.; Kumar, S.; Singh, M.; Singh, R.K. QTL mapping for important horticultural traits in pepper (Capsicum annuum L.). J. Plant Biochem. Biotechnol. 2015, 24, 154–160. [Google Scholar] [CrossRef]
- Dwivedi, N.; Kumar, R.; Singh, R.K.; Singh, M. QTL analysis associated with oleoresin content in intraspecific RIL population of chilli (Capsicum annuum L.). J. Appl. Hortic. 2014, 16, 107–111. [Google Scholar] [CrossRef]
- Eggink, P.M.; Tikunov, Y.; Maliepaard, C.; Haanstra, J.P.W.; De Rooij, H.; Vogelaar, A.; Gutteling, E.W.; Freymark, G.; Bovy, A.G.; Visser, R.G.F. Capturing flavors from Capsicum baccatum by introgression in sweet pepper. Theor. Appl. Genet. 2014, 127, 373–390. [Google Scholar] [CrossRef]
- Han, K.; Lee, H.; Ro, N.; Hur, O.; Lee, J.; Kwon, J.; Kang, B. QTL mapping and GWAS reveal candidate genes controlling capsaicinoid content in Capsicum. Plant Biotechnol. J. 2018, 16, 1546–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kim, K.-T.; Kim, D.-H.; Yang, E.-Y.; Cho, M.-C.; Jamal, A.; Chae, Y.; Pae, D.-H.; Oh, D.-G.; Hwang, J.-K. Identification of quantitative trait loci associated with anthracnose resistance in chili pepper (Capsicum spp.). Hortic. Sci. Technol. 2010, 28, 1014–1024. [Google Scholar]
- Lee, J.; Park, S.J.; Hong, S.C.; Han, J.; Choi, D.; Yoon, J.B. QTL mapping for capsaicin and dihydrocapsaicin content in a population of Capsicum annuum ‘NB 1’ × Capsicum chinense ‘Bhut Jolokia’. Plant Breed. 2016, 135, 376–383. [Google Scholar] [CrossRef]
- Lee, J.-H.; An, J.-T.; Siddique, M.I.; Han, K.; Choi, S.; Kwon, J.-K.; Kang, B.-C. Identification and molecular genetic mapping of Chili veinal mottle virus (ChiVMV) resistance genes in pepper (Capsicum annuum). Mol. Breed. 2017, 37, 121. [Google Scholar] [CrossRef]
- Lee, H.Y.; Ro, N.Y.; Patil, A.; Lee, J.H.; Kwon, J.K.; Kang, B.C. Uncovering Candidate Genes Controlling Major Fruit-Related Traits in Pepper via Genotype-by-Sequencing Based QTL Mapping and Genome-Wide Association Study. Front. Plant Sci. 2020, 11, 1100. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yin, Y.; Wang, F.; Yao, M. Construction of a high-density genetic map and identification of QTLs for cucumber mosaic virus resistance in pepper (Capsicum annuum L.) using specific length amplified fragment sequencing (SLAF-seq). Breed. Sci. 2018, 68, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-Y.; Kang, J.-H.; Jeong, H.-S.; Choi, H.-J.; Yang, H.-B.; Kim, K.-T.; Choi, D.; Choi, G.J.; Jahn, M.; Kang, B.-C. Combined use of bulked segregant analysis and microarrays reveals SNP markers pinpointing a major QTL for resistance to Phytophthora capsici in pepper. Theor. Appl. Genet. 2014, 127, 2503–2513. [Google Scholar] [CrossRef]
- Lu, F.-H.; Kwon, S.-W.; Yoon, M.-Y.; Kim, K.-T.; Cho, M.-C.; Yoon, M.-K.; Park, Y.-J. SNP marker integration and QTL analysis of 12 agronomic and morphological traits in F 8 RILs of pepper (Capsicum annuum L.). Mol. Cells 2012, 34, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Mahasuk, P.; Struss, D.; Mongkolporn, O. QTLs for resistance to anthracnose identified in two Capsicum sources. Mol. Breed. 2016, 36, 10. [Google Scholar] [CrossRef]
- Naegele, R.P.; Ashrafi, H.; Hill, T.A.; Chin-Wo, S.R.; Van Deynze, A.E.; Hausbeck, M.K. QTL mapping of fruit rot resistance to the plant pathogen Phytophthora capsici in a recombinant inbred line Capsicum annuum population. Phytopathology 2014, 104, 479–483. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Lee, J.-H.; Han, K.; Jang, S.; Han, J.; Lim, J.-H.; Jung, J.-W.; Kang, B.-C. A major QTL and candidate genes for capsaicinoid biosynthesis in the pericarp of Capsicum chinense revealed using QTL-seq and RNA-seq. Theor. Appl. Genet. 2019, 132, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Barka, G.D.; Yang, E.-Y.; Cho, M.-C.; Yoon, J.B.; Lee, J. Identification of QTLs Controlling α-Glucosidase Inhibitory Activity in Pepper (Capsicum annuum L.) Leaf and Fruit Using Genotyping-by-Sequencing Analysis. Genes 2020, 11, 1116. [Google Scholar] [CrossRef] [PubMed]
- Rehrig, W.Z.; Ashrafi, H.; Hill, T.; Prince, J.; Van Deynze, A. CaDMR1 cosegregates with QTL Pc5. 1 for resistance to Phytophthora capsici in pepper (Capsicum annuum). Plant Genome 2014, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Siddique, M.I.; Lee, H.-Y.; Ro, N.-Y.; Han, K.; Venkatesh, J.; Solomon, A.M.; Patil, A.S.; Changkwian, A.; Kwon, J.-K.; Kang, B.-C. Identifying candidate genes for Phytophthora capsici resistance in pepper (Capsicum annuum) via genotyping-by-sequencing-based QTL mapping and genome-wide association study. Sci. Rep. 2019, 9, 9962. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Mao, S.L.; Zhang, Z.H.; Palloix, A.; Wang, L.H.; Zhang, B.X. Resistances to anthracnose (Colletotrichum acutatum) of Capsicum mature green and ripe fruit are controlled by a major dominant cluster of QTLs on chromosome P5. Sci. Hortic. 2015, 181, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Cheng, J.W.; Zhang, L.; Qin, C.; Nong, D.G.; Li, W.P.; Tang, X.; Wu, Z.M.; Hu, K.L. Construction of an interspecific genetic map based on InDel and SSR for mapping the QTLs affecting the initiation of flower primordia in pepper (Capsicum spp.). PLoS ONE 2015, 10, e0119389. [Google Scholar] [CrossRef]
- Wei, J.; Li, J.; Yu, J.; Cheng, Y.; Ruan, M.; Ye, Q.; Yao, Z.; Wang, R.; Zhou, G.; Deng, M. Construction of high-density bin map and QTL mapping of horticultural traits from an interspecific cross between Capsicum annuum and Chinese wild Capsicum frutescens. Biotechnol. Biotechnol. Equip. 2020, 34, 549–561. [Google Scholar] [CrossRef]
- Yao, M.; Li, N.; Wang, F.; Ye, Z. Genetic analysis and identification of QTLs for resistance to cucumber mosaic virus in chili pepper (Capsicum annuum L.). Euphytica 2013, 193, 135–145. [Google Scholar] [CrossRef]
- Yarnes, S.C.; Ashrafi, H.; Reyes-Chin-Wo, S.; Hill, T.A.; Stoffel, K.M.; Van Deynze, A. Identification of QTLs for capsaicinoids, fruit quality, and plant architecture-related traits in an interspecific Capsicum RIL population. Genome 2013, 56, 61–74. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, G.; Dong, T.; Chen, B.; Du, H.; Li, C.; Zhang, F.; Zhang, H.; Xu, Y.; Wang, Q. High-density genetic map construction and QTL mapping of first flower node in pepper (Capsicum annuum L.). BMC Plant Biol. 2019, 19, 167. [Google Scholar] [CrossRef]
- Klein, A.; Houtin, H.; Rond-Coissieux, C.; Naudet-Huart, M.; Touratier, M.; Marget, P.; Burstin, J. Meta-analysis of QTL reveals the genetic control of yield-related traits and seed protein content in pea. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Poland, J.A.; Nelson, R.J. In the eye of the beholder: The effect of rater variability and different rating scales on QTL mapping. Phytopathology 2011, 101, 290–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vales, M.I.; Schön, C.C.; Capettini, F.; Chen, X.M.; Corey, A.E.; Mather, D.E.; Mundt, C.C.; Richardson, K.L.; Sandoval-Islas, J.S.; Utz, H.F. Effect of population size on the estimation of QTL: A test using resistance to barley stripe rust. Theor. Appl. Genet. 2005, 111, 1260–1270. [Google Scholar] [CrossRef]
- Dhingani, R.M.; Umrania, V.V.; Tomar, R.S.; Parakhia, M.V.; Golakiya, B. Introduction to QTL mapping in plants. Ann. Plant Sci. 2015, 4, 1072–1079. [Google Scholar]
- Truong, H.T.H.; Kim, K.T.; Kim, D.W.; Kim, S.; Chae, Y.; Park, J.H.; Oh, D.G.; Cho, M.C. Identification of isolate-specific resistance QTLs to phytophthora root rot using an intraspecific recombinant inbred line population of pepper (Capsicum annuum). Plant Pathol. 2012, 61, 48–56. [Google Scholar] [CrossRef]
- Xu, X.; Chao, J.; Cheng, X.; Wang, R.; Sun, B.; Wang, H.; Luo, S.; Xu, X.; Wu, T.; Li, Y. Mapping of a novel race specific resistance gene to phytophthora root rot of pepper (Capsicum annuum) using bulked segregant analysis combined with specific length amplified fragment sequencing strategy. PLoS ONE 2016, 11, e0151401. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, L.; Guo, J.; Yang, W.; Shen, H. Molecular mapping of a gene conferring resistance to Phytophthora capsici Leonian race 2 in pepper line PI201234 (Capsicum annuum L.). Mol. Breed. 2016, 36, 66. [Google Scholar] [CrossRef]
- Lu, Q.; Liu, H.; Hong, Y.; Li, H.; Liu, H.; Li, X.; Wen, S.; Zhou, G.; Li, S.; Chen, X. Consensus map integration and QTL meta-analysis narrowed a locus for yield traits to 0.7 cM and refined a region for late leaf spot resistance traits to 0.38 cM on linkage group A05 in peanut (Arachis hypogaea L.). BMC Genom. 2018, 19, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano, J.M.; Alvaro, F. Discovering consensus genomic regions in wheat for root-related traits by QTL meta-analysis. Sci. Rep. 2019, 9, 1–14. [Google Scholar]
- Stokes, T.L.; Kunkel, B.N.; Richards, E.J. Epigenetic variation in Arabidopsis disease resistance. Genes Dev. 2002, 16, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Zhai, K.; Xie, Z.; Yang, D.; Zhu, X.; Liu, J.; Wang, X.; Qin, P.; Yang, Y.; Zhang, G. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science 2017, 355, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, Q.; Wang, W.; Li, Y.; Guo, Y.; Zhang, D.; Ma, X.; Song, W.; Zhao, J.; Xu, M. A transposon-directed epigenetic change in ZmCCT underlies quantitative resistance to Gibberella stalk rot in maize. N. Phytol. 2017, 215, 1503–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawoof, A.; Ahmad, I.; Ramchiary, N. Epigenome landscape in Capsicum genome. In The Capsicum Genome; Springer: Berlin/Heidelberg, Germany, 2019; pp. 187–199. [Google Scholar]
- Stewart, C., Jr.; Mazourek, M.; Stellari, G.M.; O’Connell, M.; Jahn, M. Genetic control of pungency in C. chinense via the Pun1 locus. J. Exp. Bot. 2007, 58, 979–991. [Google Scholar] [CrossRef]
- Li, J.Z.; Zhang, Z.W.; Li, Y.L.; Wang, Q.L.; Zhou, Y.G. QTL consistency and meta-analysis for grain yield components in three generations in maize. Theor. Appl. Genet. 2011, 122, 771–782. [Google Scholar] [CrossRef]
- Semagn, K.; Babu, R.; Hearne, S.; Olsen, M. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Overview of the technology and its application in crop improvement. Mol. Breed. 2014, 33, 1–14. [Google Scholar] [CrossRef]
- Sy, O.; Steiner, R.; Bosland, P.W. Recombinant inbred line differential identifies race-specific resistance to Phytophthora root rot in Capsicum annuum. Phytopathology 2008, 98, 867–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozada, D.N.; Bhatta, M.; Coon, D.; Bosland, P.W. Single Nucleotide Polymorphisms Reveal Genetic Diversity in New Mexican Chile Peppers (Capsicum Spp.). BMC Genom. 2021, 11, e0161464. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Study No. | Reference | Parents Used in Hybridization | No. of Individuals | Mapping Pop. 1 | Trait(s) | Marker Type 2 |
|---|---|---|---|---|---|---|
| 1 | Alimi et al. [37] | Yolo Wonder × CM-334 | 149 | RIL | Fruit and stem-related traits | SSR |
| 2 | Arjun et al. [38] | C. annuum FL 201 × C. galapagoense TC 07245 | 210 | F2 | Fruit length | SSR |
| 3 | Chunthawodtiporn et al. [39] | C. annuum Maor × CM-334 | 120 | RIL | Horticultural traits | SNP |
| 4 | Du at al. [40] | BVRC 25 × BVRC 1 | 440 | F2 | Bacterial wilt resistance | SNP, InDel |
| 5 | Dwivedi et al. [41] | California Wonder × LCA235 | 74 | RIL | Plant height and yield related traits | SSR, SCAR, RAPD |
| 6 | Dwivedi et al. [42] | California Wonder × LCA235 | 74 | RIL | Oleoresin content | SSR, SCAR, RAPD |
| 7 | Eggink et al. [43] | MT x PEN45 (MP); SM × PEN45 (SP); GNM 3 | 250 | BC | Agronomic, biochemical, physiological traits | AFLP, SNP |
| 8 | Han et al. [44] | Perennial × Dempsey (PD); TF68 × Habanero (TH) | 56 (PD), 85 (TH) | RIL | Capsaicinoid content | SNP |
| 9 | Kim et al. [45] | C. baccatum var. Pendulum × Golden-aji | 126 | F2 | Anthracnose resistance | SSR, AFLP, SRAP |
| 10 | Lee et al. [46] | NB1 × Bhut Jolokia | 175 | F2 | Capsaicinoid content | HRM, SSR, CAPS, Gene-based |
| 11 | Lee et al. [47] | CV4 × Jeju | 300 | F2 | Chili veinal mottle virus | SNP |
| 12 | Lee et al. [48] | Perennial × Dempsey | 120 | RIL | Fruit-related traits | SNP |
| 13 | Li et al. [49] | BJ0747 × XJ0630 | 195 | F2 | Cucumber mosaic virus resistance | SLAF-SNP |
| 14 | Liu et al. [50] | YCM334 × Taean | 128 | RIL | P. capsici resistance | SNP |
| 15 | Lu et al. [51] | YCM334 × Taean | 126 | RIL | Agronomic and morphological traits; P. capsici resistance | SNP |
| 16 | Mahasuk et al. [52] | Bangchang × PBC932 (BP), PBC80 × CA1316 (PC) | 126 (BP), 146 (PC) | F2 | Anthracnose resistance | SNP |
| 17 | Mallard et al. [25] | H3 × Vania (HV), Perennial × Yolo Wonder (PY), YW × CM-334 (F5YC) | 101 (HV), 114 (PY), 297 (RIL) | DH (PY, HV), RIL (F5YC) | P. capsici resistance | AFLP, CAPS, RFLP |
| 18 | Naegele et al. [53] | CM-334 × EJ | 63 | RIL | P. capsici resistance | SNP |
| 19 | Park et al. [54] | Habanero × Jolokia (HJ), SNU11-001 × Jolokia (SJ) | 87 (HJ), 124 (SJ) | F2 | Capsaicinoid content | SNP |
| 20 | Park et al. [55] | M5 × AG13-3 | 96 | F2 | AGI activity | SNP |
| 21 | Rehrig et al. [56] | CM-334 × EJ | 66 | RIL | P. capsici resistance | SNP |
| 22 | Siddique et al. [57] | CM-334 × ECW30R | 188 | RIL | P. capsici resistance | SNP |
| 23 | Sun et al. [58] | C. annuum 77013 × C. chinense PBC932 | 186 | BC | Anthracnose resistance | SSR, InDel, CAPS |
| 24 | Tan et al. [59] | C. annuum BA3 × C. frutescens YNXML | 154 | F2 | Flowering time | SSR, InDel |
| 25 | Wei et al. [60] | C. annuum 007EA × C. frutescens P1512 | 120 | F2 | Horticultural traits | SNP |
| 26 | Yao et al. [61] | BJ0747-1-3-1-1 × XJ0630-2-1-2-1-1 | 334 | F2 | Cucumber mosaic virus resistance | SSR, ISSR |
| 27 | Yarnes at al. [62] | 2814-6 × NuMex R Naky | 105 | RIL | Plant architecture, phenology, fruit quality | SPP |
| 28 | Zhang et al. [63] | PM702 × FS871 | 146 | RIL | First flower node | SLAF/SNP |
| 29 | Zhu et al. [27] | C. chinense 740 × C. annuum CA1 | 150 | F2 | Flower number, flowering time | SLAF |
| Chr. | Mean Length (cM) | No. of SNP Markers |
|---|---|---|
| P1 | 134.90 | 986 |
| P2 | 113.95 | 1442 |
| P3 | 130.42 | 2231 |
| P4 | 116.43 | 707 |
| P5 | 147.70 | 1165 |
| P6 | 94.97 | 1711 |
| P7 | 127.70 | 1996 |
| P8 | 175.09 | 431 |
| P9 | 97.36 | 878 |
| P10 | 112.47 | 722 |
| P11 | 137.91 | 1529 |
| P12 | 120.63 | 1154 |
| Total | 1509.52 | 14,952 |
| Average | 125.79 | 1246 |
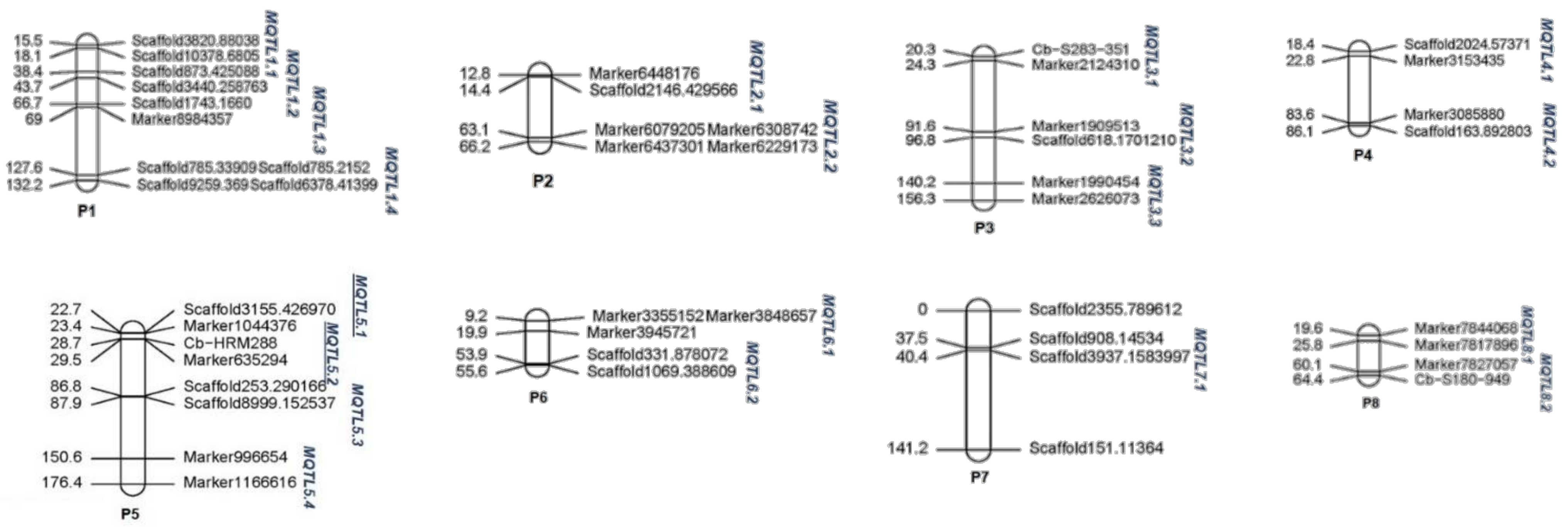
| Chr. | No. of QTL | No. of Projected QTL | No. of Meta-QTL | QTL Model | AIC Value 1 | Meta-QTL 2 | No. of QTL in the Meta-QTL | Mean Genetic Position (cM) | CI 3 (cM) |
|---|---|---|---|---|---|---|---|---|---|
| P1 | 73 | 61 | 4 | 5 | 555.66 | MQTL1.1 | 15 | 16.75 | 2.59 |
| MQTL1.2 | 6 | 40.95 | 5.27 | ||||||
| MQTL1.3 | 36 | 67.62 | 2.11 | ||||||
| MQTL1.4 | 4 | 129.82 | 4.25 | ||||||
| P2 | 64 | 27 | 2 | 5 | 424.59 | MQTL2.1 | 13 | 12.19 | 1.6 |
| MQTL2.2 | 14 | 71.44 | 2.9 | ||||||
| P3 | 43 | 27 | 3 | 5 | 307.38 | MQTL3.1 | 6 | 22.33 | 4.05 |
| MQTL3.2 | 6 | 94.16 | 5.14 | ||||||
| MQTL3.3 | 15 | 146.12 | 1.65 | ||||||
| P4 | 35 | 29 | 2 | 5 | 295.78 | MQTL4.1 | 10 | 20.52 | 3.79 |
| MQTL4.2 | 19 | 84.92 | 2.02 | ||||||
| P5 | 51 | 45 | 4 | 5 | 316.37 | MQTL5.1 | 13 | 23.03 | 0.79 |
| MQTL5.2 | 25 | 29.16 | 0.55 | ||||||
| MQTL5.3 | 7 | 87.33 | 1.07 | ||||||
| MQTL5.4 | 2 | 156.87 | 12.42 | ||||||
| P6 | 27 | 20 | 2 | 5 | 217.58 | MQTL6.1 | 2 | 14.7 | 10.49 |
| MQTL6.2 | 19 | 54.73 | 1.65 | ||||||
| P7 | 11 | 8 | 1 | 3 | 82.13 | MQTL7.1 | 8 | 38.99 | 2.53 |
| P8 | 15 | 14 | 2 | 5 | 124.72 | MQTL8.1 | 9 | 22.13 | 5.61 |
| MQTL8.2 | 2 | 62.24 | 4.01 | ||||||
| P9 | 17 | 10 | 2 | 5 | 124.95 | MQTL9.1 | 4 | 43.8 | 7.87 |
| MQTL9.2 | 3 | 74.89 | 9.43 | ||||||
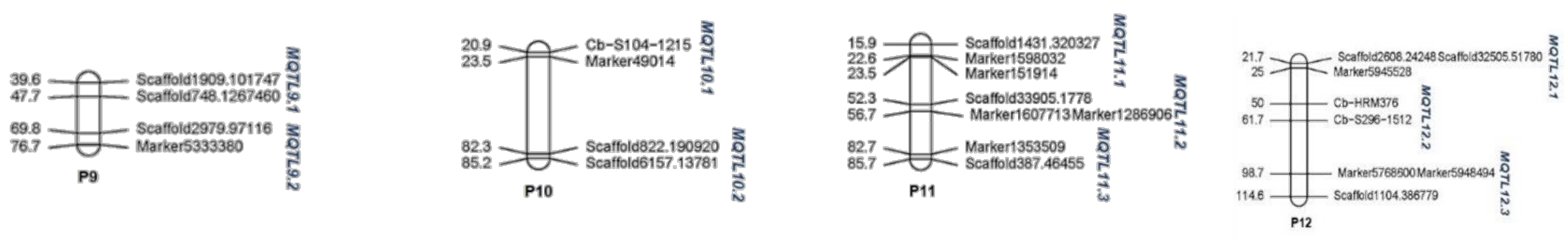
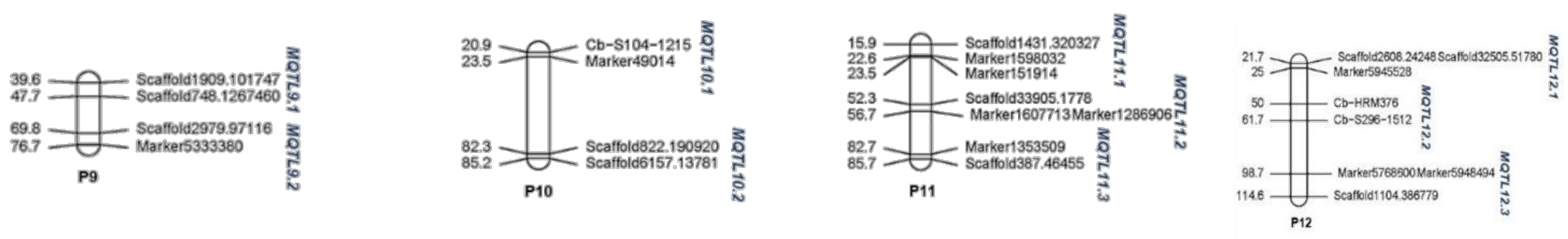
| P10 | 35 | 26 | 2 | 5 | 231.24 | MQTL10.1 | 13 | 22.39 | 1.86 |
| MQTL10.2 | 13 | 83.87 | 2.29 | ||||||
| P11 | 24 | 23 | 3 | 5 | 214.53 | MQTL11.1 | 5 | 19.3 | 5.72 |
| MQTL11.2 | 11 | 54.56 | 4.01 | ||||||
| MQTL11.3 | 7 | 84.22 | 3.0 | ||||||
| P12 | 24 | 21 | 3 | 4 | 242.70 | MQTL12.1 | 13 | 23.51 | 2.82 |
| MQTL12.2 | 2 | 55.69 | 10.97 | ||||||
| MQTL12.3 | 6 | 106.76 | 14.90 | ||||||
| Total | 419 | 311 | 30 | 308 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozada, D.N.; Whelpley, M.; Acuña-Galindo, A. Genetic Architecture of Chile Pepper (Capsicum spp.) QTLome Revealed Using Meta-QTL Analysis. Horticulturae 2021, 7, 227. https://doi.org/10.3390/horticulturae7080227
Lozada DN, Whelpley M, Acuña-Galindo A. Genetic Architecture of Chile Pepper (Capsicum spp.) QTLome Revealed Using Meta-QTL Analysis. Horticulturae. 2021; 7(8):227. https://doi.org/10.3390/horticulturae7080227
Chicago/Turabian StyleLozada, Dennis N., Madelin Whelpley, and Andrea Acuña-Galindo. 2021. "Genetic Architecture of Chile Pepper (Capsicum spp.) QTLome Revealed Using Meta-QTL Analysis" Horticulturae 7, no. 8: 227. https://doi.org/10.3390/horticulturae7080227
APA StyleLozada, D. N., Whelpley, M., & Acuña-Galindo, A. (2021). Genetic Architecture of Chile Pepper (Capsicum spp.) QTLome Revealed Using Meta-QTL Analysis. Horticulturae, 7(8), 227. https://doi.org/10.3390/horticulturae7080227