
Analysis of Glucosinolate Content, Composition and Expression Level of Biosynthesis Pathway Genes in Different Chinese Kale Varieties


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. GLSs Extraction and Purification
2.3. HPLC-MS Analysis of GLSs
2.4. Expression Analysis of Genes Involving in the Glucosinolate Biosynthesis
2.5. Statistical Analysis
3. Results
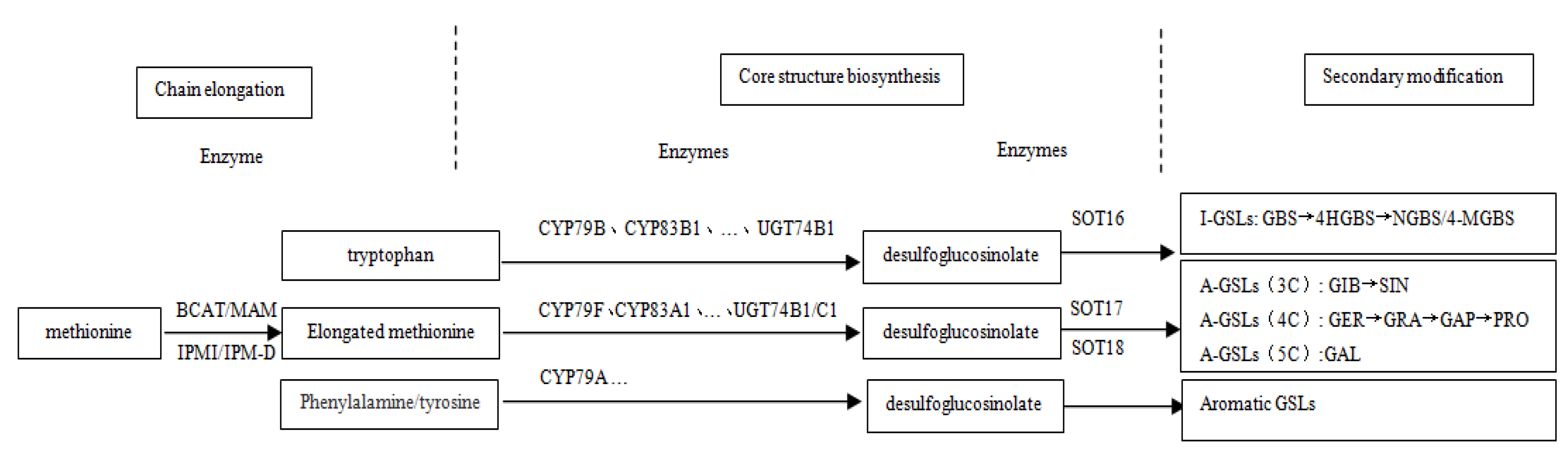
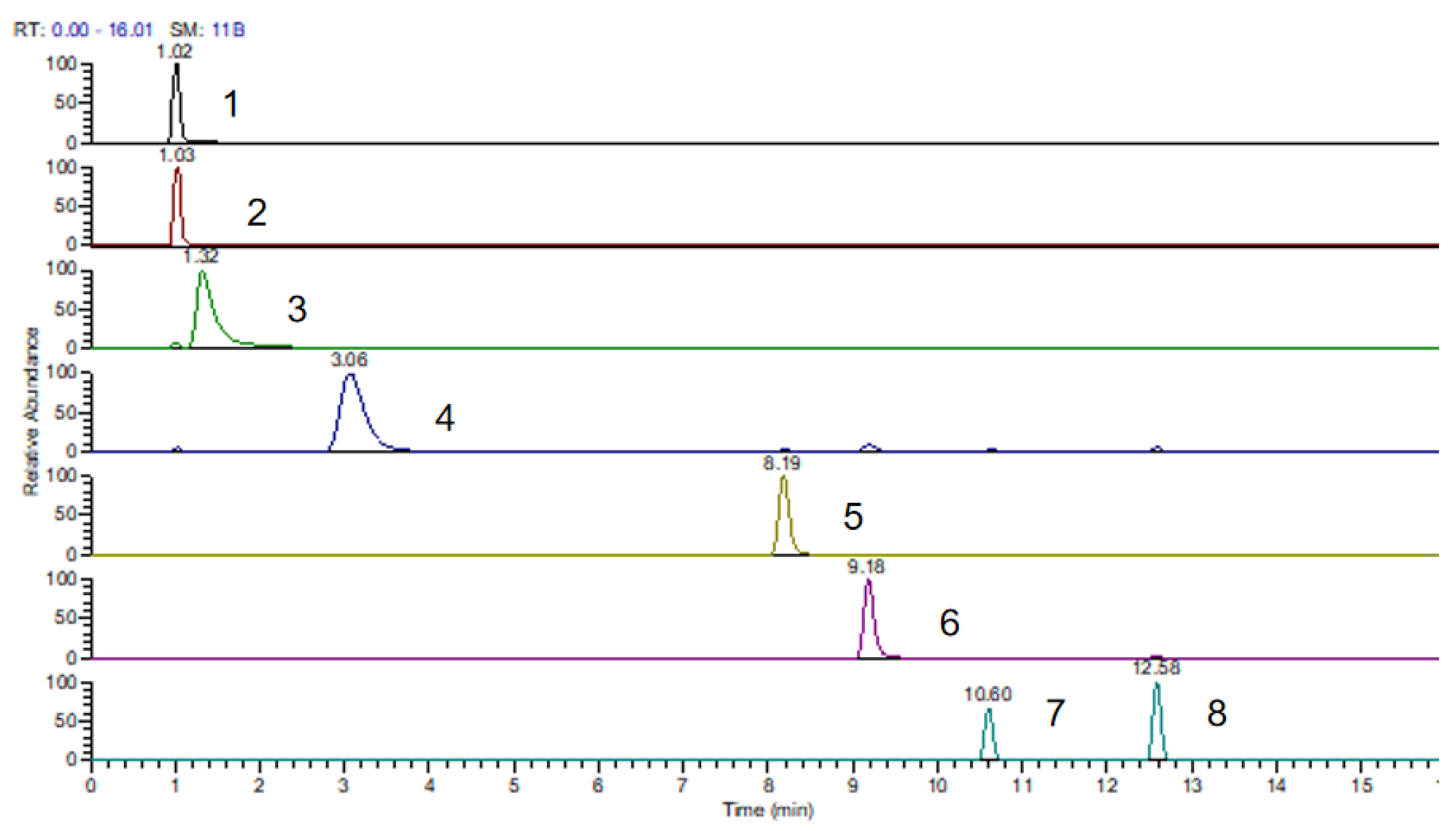
3.1. Confirmation of Chinese Kale Desulfo-Glucosinolates
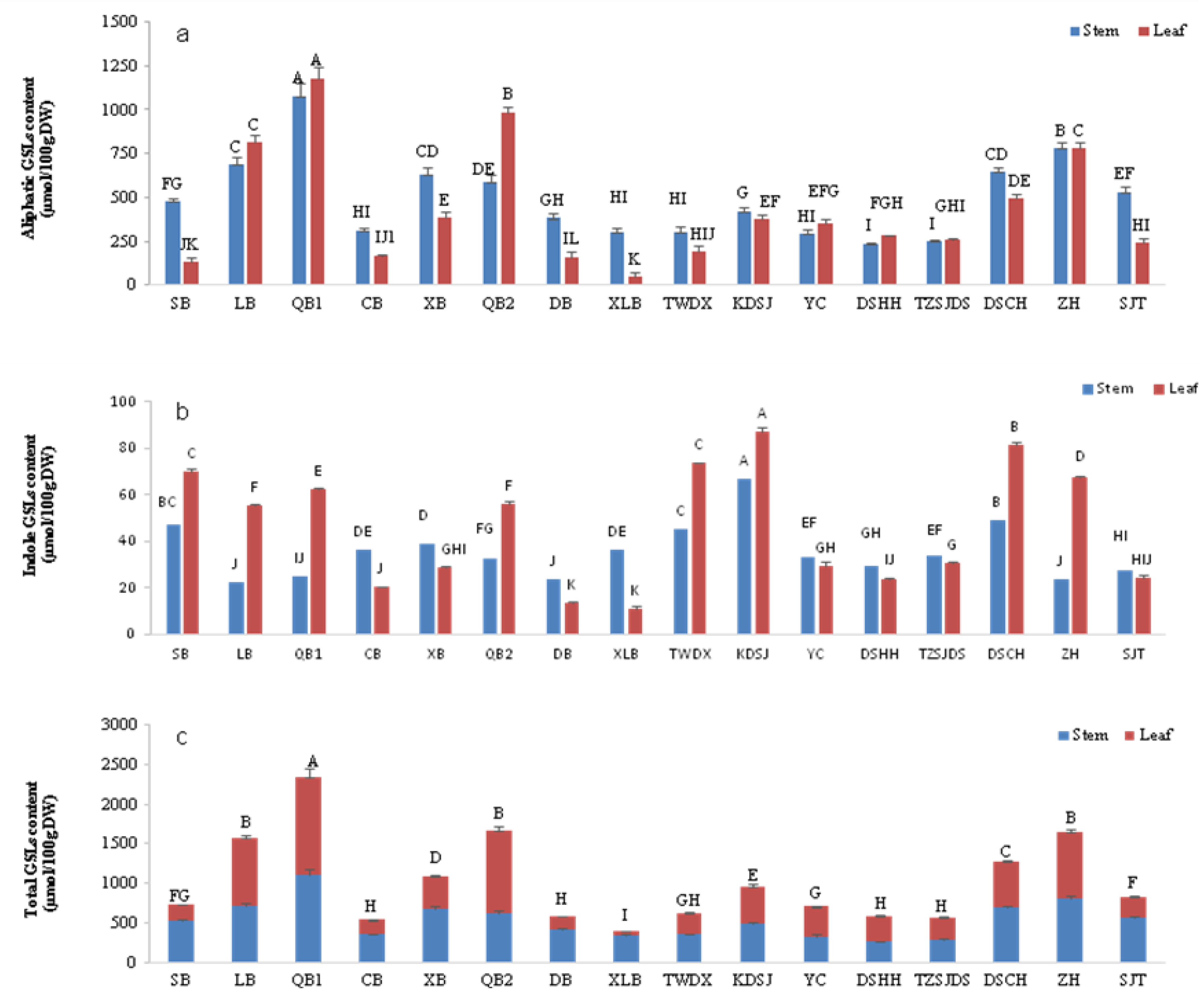
3.2. The Content and Component Analysis of GLSs in Edible Stems and Leaves of Sixteen Chinese Kale Varieties
3.3. Analysis of GLSs Components and Contents in Edible Leaves of Chinese Kale Varieties
3.4. Analysis of GLSs Components and Contents in Edible Stems of Chinese Kale Varieties
3.5. Expression Analysis of Key Genes in GLSs Biosynthesis Pathway
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Becerra-Moreno, A.; Alanís-Garza, P.A.; Mora-Nieves, J.L.; Mora-Mora, J.P.; Jacobo-Velázquez, D.A. Kale: An excellent source of vitamin C, pro-vitamin A, lutein and glucosinolates. CyTA J. Food 2014, 12, 298–303. [Google Scholar] [CrossRef]
- Abbaoui, B.; Lucas, C.R.; Riedl, K.M.; Clinton, S.K.; Mortazavi, A. Cruciferous vegetables, isothiocyanates, and bladder cancer prevention. Mol. Nutr. Food Res. 2018, 62, 1–14. [Google Scholar] [CrossRef]
- Feskanich, D.; Ziegler, R.G.; Michaud, D.S.; Giovannucci, E.L.; Speizer, F.E.; Willett, W.C.; Colditz, G.A. Prospective study of fruit and vegetable consumption and risk of lung cancer among men and women. J. Natl. Cancer Inst. 2000, 92, 1812–1823. [Google Scholar] [CrossRef]
- Joseph, M.A.; Moysich, K.B.; Freudenheim, J.L.; Shields, P.G.; Bowman, E.D.; Zhang, Y.; Marshall, J.R.; Ambrosone, C.B. Cruciferous vegetables, genetic polymorphisms in glutathione S-transferases M1 and T1, and prostate cancer risk. Nutr. Cancer 2004, 50, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Ménard, R.; Larue, J.P.; Silué, D.; Thouvenot, D. Glucosinolates in cauliflower as biochemical markers for resistance against downy mildew. Phytochemistry 1999, 52, 29–35. [Google Scholar] [CrossRef]
- Grubb, C.D.; Abel, S. Glucosinolate metabolism and its control. Trends Plant Sci. 2006, 11, 89–100. [Google Scholar] [CrossRef]
- Zeng, W.; Tao, H.; Li, Y.; Wang, J.; Xia, C.; Li, S.; Wang, M.; Wang, Q.; Miao, H. The flavor of Chinese kale sprouts is affected by genotypic variation of glucosinolates and their breakdown products. Food Chem. 2021, 359, 129824. [Google Scholar] [CrossRef]
- Sikorska-Zimny, K.; Beneduce, L. The glucosinolates and their bioactive derivatives in Brassica: A review on classification, biosynthesis and content in plant tissues, fate during and after processing, effect on the human organism and interaction with the gut microbiota. Crit. Rev. Food Sci. Nutr. 2021, 61, 2544–2571. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, T.; Bobo, G.; Vinas, I.; Collazo, C.; Aguilo-Aguayo, I. Effects of thermal and non-thermal processing of cruciferous vegetables on glucosinolates and its derived forms. J. Food Sci. Technol. 2018, 55, 1973–1981. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.B.; Williams, P.H.; Carlson, D.G.; Tookey, H.L. Variation in glucosinolates in oriental brassica’ vegetables. J. Am. Soc. Hortic. Sci. 1987, 112, 309–313. [Google Scholar]
- Nugrahedi, P.Y.; Oliviero, T.; Heising, J.K.; Dekker, M.; Verkerk, R. Stir-frying of Chinese cabbage and pakchoi retains health- promoting glucosinolates. Plant Foods Hum. Nutr. 2017, 72, 439–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, H.; Ping, L.; Bonnema, G.; Dekker, M.; Verkerk, R. Genetic variation in glucosinote content within Brassica rapa vegetables. Acta Hortic. 2012, 944, 129–140. [Google Scholar]
- Frazie, M.D.; Kim, M.J.; Ku, K.M. Health-promoting phytochemicals from 11 mustard cultivars at baby leaf and mature stages. Molecules 2017, 22, 1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco, L.; Becker, H.C. Variability for seed glucosinolates in a germplasm collection of the genus Brassica. Genet. Resour. Crop Evol. 2000, 47, 231–238. [Google Scholar] [CrossRef]
- Velasco, L.; Becker, H.C. Analysis of total glucosinolate content and individual glucosinolates in Brassica spp. by near-infrared reflectance spectroscopy. Plant Breed. 1998, 117, 97–102. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Schreiner, M. Isothiocyanates, nitriles, and epithionitriles from glucosinolates are affected by genotype and developmental stage in Brassica oleracea varieties. Front. Plant Sci. 2017, 8, 1095. [Google Scholar] [CrossRef] [Green Version]
- Ku, K.M.; Jeffery, E.H.; Juvik, J.A. Optimization of methyl jasmonate application to broccoli florets to enhance health-promoting phytochemical content. J. Sci. Food Agri. 2014, 94, 2090–2096. [Google Scholar] [CrossRef]
- Carlson, D.G.; Daxenbichler, M.E.; Van Etten, C.H.; Kwolek, W.F.; Williams, P.H. Glucosinolates in crucifer vegetables: Broccoli, Brussels sprouts, cauliflower, collards, kale, mustard greens and kohlrabi. J. Am. Soc. Hortic. Sci. 1987, 112, 173–178. [Google Scholar]
- Oerlemans, K.; Barrett, D.M.; Suades, C.B.; Verkerk, R.; Dekker, M. Thermal degradation of glucosinolates in red cabbage. Food Chem. 2006, 95, 19–29. [Google Scholar] [CrossRef]
- He, H.; Song, S.; Wang, W.; Xu, X. HPLC identification of intact glucosinolates in Chinese kale. Mod. Instrum. 2002, 5, 10–12. [Google Scholar]
- Sun, B.; Liu, N.; Zhao, Y.; Yan, H.; Wang, Q. Variation of glucosinolates in three edible parts of Chinese kale (Brassica alboglabra Bailey) varieties. Food Chem. 2011, 124, 941–947. [Google Scholar] [CrossRef]
- Klopsch, R.; Witzel, K.; Artemyeva, A.; Rupperl, S.; Hanschen, F.S. Genotypic profiling of glucosinlats and their breakdown products in leaves of Brassica rapa. J. Agric. Food Chem. 2018, 66, 5481–5490. [Google Scholar] [CrossRef] [PubMed]
- Sønderby, I.E.; Geu-Flores, F.; Halkier, B.A. Biosynthesis of glucosinolates-gene discovery and beyond. Trends Plant Sci. 2010, 15, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Yuji, S.; Ayuko, A.; Mutsumi, N.; Tomoko, N.; Akane, S.; Kazuki, S.; Masamo, Y.H. Omics-based approaches to methionine side chain elongation in Arabidopsis: Characterization of the genes encoding methylthioalkylmalate isomerase and methylthioalkyl malate dehydrogenase. Plant Cell Physiol. 2009, 50, 1181–1190. [Google Scholar]
- Yuji, S.; Kiminori, T.; Ayuko, K.; Akane, S.; Mutsumi, N.; Kazuki, S.; Masami, Y.H. Arabidopsis bile acid: Sodium symporter family protein 5 is involved in methionine-derived glucosinolate biosynthesis. Plant Cell Physiol. 2009, 50, 1579–1586. [Google Scholar]
- Hull, A.K.; Vij, R.; Celenza, J.L. Arabidopsis cytochrome P450s that catalyze the first step of tryptophan-dependent indole- 3-acetic acid biosynthesis. Proc. Natl. Acad. Sci. USA 2000, 97, 2379–2384. [Google Scholar] [CrossRef] [Green Version]
- Mikkelsen, M.D.; Hansen, C.H.; Wittstock, U.; Halkier, B.A. Cytochrome P450 CYP79B2 from Arabidopsis catalyzes the conversion of tryptophan to indole-3-acetaldoxime, a precursor of indole glucosinolates and indole-3-acetic acid. J. Biol. Chem. 2000, 275, 33712–33717. [Google Scholar] [CrossRef] [Green Version]
- Bak, S.; Feyereisen, R. The involvement of two P450 enzymes, CYP83B1 and CYP83A1, in auxin homeostasis and glucosinolate biosynthesis. Plant Physiol. 2001, 127, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Mikkelsen, M.D.; Naur, P.; Ahalkier, B. Arabidopsis mutants in the C–S lyase of glucosinolate biosynthesis establish a critical role for indole-3-acetaldoxime in auxin homeostasis. Plant J. 2004, 37, 770–777. [Google Scholar] [CrossRef]
- Grubb, C.D.; Zipp, B.J.; Kopycki, J.; Schubert, M.; Quint, M.; Lim, E.-K.; Bowles, D.J.; Pedras, M.S.C.; Abel, S. Comparative analysis of Arabidopsis UGT74 glucosyltransferases reveals a special role of UGT74C1 in glucosinolate biosynthesis. Plant J. 2014, 79, 92–105. [Google Scholar] [CrossRef]
- Grubb, C.D.; Zipp, B.J.; Ludwig-Muller, J.; Masuno, M.N.; Molinski, T.F.; Abel, S. Arabidopsis glucosyltransferase UGT74B1 functions in glucosinolate biosynthesis and auxin homeostasis. Plant J. 2005, 40, 893–908. [Google Scholar] [CrossRef]
- Piotrowski, M.; Schemenewitz, A.; Lopukhina, A.; Muller, A.; Janowltz, T.; Weiler, E.W.; Oecking, C. Desulfoglucosinolate sulfotransferases from Arabidopsis thaliana catalyze the final step in the biosynthesis of the glucosinolate core structure. J. Biol. Chem. 2004, 279, 50717–50725. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y. Function Analyses of Several Genes Involved in Biosynthesis and Regulation of Glucosinolate in Brassica Napus and Arabidopsis Thaliana. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2015. [Google Scholar]
- Li, Y.; Gao, M.; He, R.; Zhang, Y.; Song, S.; Su, W.; Liu, H. Far-red light suppresses glucosinolate profiles of Chinese kale through inhibiting genes related to glucosinolate biosynthesis. Environ. Exp. Bot. 2021, 188, 104507. [Google Scholar] [CrossRef]
- Naur, P.; Petersen, B.L.; Mikkelsen, M.D.; Bak, S.; Rasmussen, H.; Olsen, C.E.; Halkier, B.A. CYP83A1 and CYP83B1, two nonredundant cytochrome P450 enzymes metabolizing oximes in the biosynthesis of glucosinolates in Arabidopsis. Plant Physiol. 2003, 133, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D. Cloning and Functional Studies of the BocGTR1s Related to the Glucosinolate Transporter of Chinese Kale. Master’s Thesis, South China Agricultural University, Guangzhou, China, 2018. [Google Scholar]
- Yin, L. Molecular Cloning, Expression Pattern and Characterization on Glucoraphanin Biosynthetic Related Genes in Chinese Kale (Brassica oleracea var. alboglabra Bailey). Ph.D. Thesis, South China Agricultural University, Guangzhou, China, 2016. [Google Scholar]
- Kim, M.; Chiu, Y.C.; Kim, N.; Park, H.; Lee, C.; Juvik, J.; Ku, K.M. Cultivar-specific changes in primary and secondary metabolites in Pak choi by methyl jasmonate. Int. J. Mol. Sci. 2017, 18, 1–17. [Google Scholar]
- Kim, S.; Chiami, K.; Ishii, G. Effect of ammonium: Nitrate nutrient ratio on nitrate and glucosinolate contents of hydroponically- grown rocket salad (Eruca sativa Mill.). Soil Sci. Plant Nutr. 2006, 52, 387–393. [Google Scholar] [CrossRef]
- Krumbein, A.B.; Schonhof, I.; Schreiner, M. Composition and contents of phytochemicals (glucosinolates, carotenoids and chlorophylls) and ascorbic acid in selected Brassica species (B. juncea, B. campestris, B. rapa). J. Appl. Bot. Food Qual. 2005, 79, 168–174. [Google Scholar]
- La, G.; Liu, G.; Fang, P. Glucosinolates in ten cultivars of Chinese kale (Brassica alboglabra LH Bailey) seperation and identification by HPLC-PAD-ESI/MS[C]//2011. In Proceedings of the International Conference on New Technology of Agricultural Engineering, Zibo, China, 27–29 May 2011; IEEE: Piscataway, NJ, USA, 2011; pp. 979–983. [Google Scholar]
- Yuan, W.; Yuan, S.; Chen, L.; Liu, Z.; Wang, K.; Qiu, Z. Identification and analysis of components of glucosinolates in succulent roots and leaves of fruity radish (Raphanus sativas L.). Asian Agric. Res. 2018, 10, 84–88. [Google Scholar]
- Zabaras, D.; Roohani, M.; Krishnamurthy, R.; Cochet, M.; Delahunty, C.M. Characterisation of taste-active extracts from raw Brassica oleracea vegetables. Food Funct. 2013, 4, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Zhang, W.; Wang, M.; Xiong, S.; Hu, D.; Sun, X.; Hu, L.; Meng, J.; Zou, J. Characterizing glucosinolates of four Brassica species and interspecific transferring of specific glucosinolates. J. Plant Genet. Resour. 2020, 21, 94–104. [Google Scholar]
- Qian, H.; Sun, B.; Miao, H.; Cai, C.; Xu, C.; Wang, Q. Variation of glucosinolates and quinone reductase activity among different varieties of Chinese kale and improvement of glucoraphanin by metabolic engineering. Food Chem. 2015, 168, 321–326. [Google Scholar] [CrossRef]
- Sun, B.; Yan, H.; Zhang, F.; Wang, Q. Effects of plant hormones on main health-promoting compounds and antioxidant capacity of Chinese kale. Food Res. Int. 2012, 48, 359–366. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, L.; Liu, G.; Zhang, D.; He, H. Evaluation of the nutritional quality of Chinese kale (Brassica alboglabra Bailey) using UHPLC-Quadrupole-Orbitrap MS/MS-Based metabolomics. Molecules 2017, 22, 1–17. [Google Scholar]
- Arora, R.; Kumar, R.; Mahajan, J.; Vig, A.P.; Singh, B.; Arora, S. 3-Butenyl isothiocyanate: A hydrolytic product of glucosinolate as a potential cytotoxic agent against human cancer cell lines. J. Food Sci. Technol. 2016, 53, 3437–3445. [Google Scholar] [CrossRef] [Green Version]
- Vallejo, F.; Tomás-Barberán, F.; García-Viguera, C. Health-promoting compounds in broccoli as influenced by refrigerated transport and retail sale period. J. Agric. Food Chem. 2003, 51, 3029–3034. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Zhang, Y.; Talalay, P. Broccoli sprouts: An exceptionally rich source of inducers of enzymes that protect against chemical carcinogens. Proc. Natl. Acad. Sci. USA 1997, 94, 10367–10372. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Beltrán, C.E.; Calderón-Oliver, M.; Pedraza-Chaverri, J.; Chirino, Y.I. Protective effect of sulforaphane against oxidative stress: Recent advances. Exp. Toxicol. Pathol. 2012, 64, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Pu, D.; Zhao, Y.; Chen, J.; Sun, Y.; Lv, A.; Zhu, S.; Luo, C.; Zhao, K.; Xiao, Q. Protective effects of sulforaphane on cognitive impairments and AD-like lesions in diabetic mice are associated with the upregulation of Nrf2 transcription activity. Neuroscience 2018, 381, 35–45. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.P.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef]
- Yi, G.E.; Arif, R.; Yang, K.; Jong-in, P.; Kang, J.K.; Yang, T.J.; Ill-Sup, N. Identification and expression analysis of glucosinolate biosynthetic genes and estimation of glucosinolate contents in edible organs of Brassica oleracea subspecies. Molecules 2015, 20, 13089–13111. [Google Scholar] [CrossRef]
- Yi, G.E.; Arif, R.; Yang, K.; Jong-in, P.; Byung, H.; Ill-Sup, N. Exogenous methyl jasmonate and salicylic acid induce subspecies- specific patterns of glucosinolate accumulation and gene expression in Brassica oleracea L. Molecules 2016, 21, 1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.; Wang, M.; Jian, Y.; Zhang, F.; Sun, B. Health-promoting phytochemicals and antioxidant capacity in different organs from six varieties of Chinese kale. Sci. Rep. 2019, 9, 20344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, Y.; Chen, G.; Lei, J.; Cao, B.; Feng, A. Analysis on composition and content of glucosinolates in different genotypes of Chinese kale. China Veg. 2009, 6, 7–13. [Google Scholar]
- Chen, X.; Zhu, Z.; Yang, J.; Liu, Y.H. Composition and content of glucosinolates in leaves and bolting stem s of Chinese kale quantified by HPLC. Acta Hortic. Sin. 2006, 4, 741–744. [Google Scholar]
- Wu, Q.; Wang, J.; Mao, S.; Xu, H.; Wu, Q.; Liang, M.; Yuan, Y.; Liu, M.; Huang, K. Comparative transcriptome analyses of genes involving in sulforaphane metabolism at different treatment in Chinese kale using full-length transcriptome sequencing. BMC Genom. 2019, 20, 377. [Google Scholar] [CrossRef]
- Zheng, H.; Zhang, C.; Wang, Y.; Zhou, W.; Sun, B. Overexpression of the glucosyltransferase gene BoaUGT74B1 enhances the accumulation of indole glucosinolates in Chinese kale. Sci. Hortic. 2021, 288, 110302. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Abbreviation | Source | Name | Abbreviation | Source |
|---|---|---|---|---|---|
| Shun bao | SB | Japan | Tai wan di xian | TWDX | China |
| Lv bao | LB | Japan | Kuai da si ji | KDSJ | China |
| Qi bao | QB1 | Japan | Yicai | YC | China |
| Cui bao | CB | Japan | Da sun huang hua | DSHH | China |
| Xiang bao | XB | Japan | Te zao si ji da sun | TZSJDS | China |
| Qi bao | QB2 | Japan | Da sun chi hua | DSCH | China |
| De ba | DB | Japan | Zhong hua | ZH | China |
| Xi lai bao | XLB | Japan | Si ji tian | SJT | China |
| Primer | Sequence 5′—3′ | Reference Genes ID and Sources of Species |
|---|---|---|
| Actin1 | F:5′-GGAATATGATGAGTCAGGCCC | XM_009117825.3 Brassica rapa (field mustard) |
| Actin1 | R:5′-CCCAGTAAGGATGAATTGGAAA | |
| CYP83A1 | F:5′-GGTTCTCCTCTTCTTCCTCTCT | KM111290.1 Brassica oleracea (wild cabbage) |
| CYP83A1 | R:5′-CCACCATTGTTTGACTTCCTAT | |
| MAM1 | F:5′-TATTTGATACGACGCTCCG | KP295465.1 Brassica oleracea (wild cabbage) |
| MAM1 | R:5′-CAACTTCCATGATGTCTACTCTG | |
| UGT74B1 | F:5′-CCGAAACCTTAACCCGAGTA | KP693685.1 Brassica oleracea var. italica |
| UGT74B1 | R:5′-GTTGGTGAAGAAAGCAGCAG | |
| SOT16 | F:5′-CTAATCCTTTGCCGTTTGTG | KP055788.1 Brassica napus (rape) |
| SOT16 | R:5′-CCTCCCTCTCCTTATCTCCTT | |
| SOT17 | F:5′-AATAAAGGGGAGAAAGATAGGG | KP055792.1 Brassica napus (rape) |
| SOT17 | R:5′-TGAAGCAAGAAAGCCAGTT | |
| SOT18 | F:5′-CCCAAAGACAGGCACCACTT | XM_013733021.1 Brassica oleracea (wild cabbage) |
| SOT18 | R:5′-GGAATCGTCGAAGCGAGATC | |
| CYP83B1 | F:5′-AGACATCCCCAATCTCCCTT | AF528175.1 Brassica rapa (field mustard) |
| CYP83B1 | R:5′-CCACTCCTTTCTGCTCGTTC | |
| CYP79A2 | F:5′-TGGCTAAAGACACCAACGG | EU877074.1 Brassica rapa (field mustard) |
| CYP79A2 | R:5′-CGAATACTACATGCTACACGCT |
| No. a | RT b (min) | Semi-Systematic Names of R-Groups | Trivial Name | Abbreviation | Compound Groups | m/z Value | Response Factor d | |||
|---|---|---|---|---|---|---|---|---|---|---|
| [M − G + H]+ | [M + H]+ | [M + Na]+ | [M + K]+ | |||||||
| 1 | 1.02 | 4-methylsulphinylbutyl | Glucoraphanin | GRA | Aliphatic | 196.0455 | 258.0968 | 380.0801 | ND c | 1.07 |
| 2 | 1.03 | 2-propenyl | Sinigrin | SIN | Aliphatic | 118.0319 | 280.0841 | ND | ND | 1.00 |
| 3 | 1.32 | 3-butenyl | Gluconapin | GNA | Aliphatic | 132.0472 | 294.0992 | 316.0811 | 332.0549 | 1.11 |
| 4 | 3.06 | 4-hydroxy-3-indolylmethyl | 4-Hydroxyglucobrassicin | 4-HGBS | Indolyl | 223.0528 | 385.1051 | 407.0870 | 423.0816 | 0.28 |
| 5 | 8.19 | 4-methylthiobutyl | Glucoerucin | GER | Aliphatic | 180.0503 | 342.1025 | 364.0841 | 380.0789 | 1.00 |
| 6 | 9.16 | 3-methylindolyl | Glucobrassicin | GBS | Indolyl | 207.0578 | 369.1098 | 391.0916 | 407.0657 | 0.29 |
| 7 | 10.60 | 4-methoxy-3-indolylmethyl | 4-Methoxyglucobrassicin | 4-MGBS | Indolyl | 237.0682 | 399.1205 | 421.1022 | 437.0761 | 0.25 |
| 8 | 12.58 | 1-methoxy-3-indolylmethyl | Neoglucobrassincin | NGBS | Indolyl | 237.0682 | 399.1205 | 421.1022 | 437.0763 | 0.20 |
| Abbreviation of Chinese Kale Varieties | Aliphatic Glucosinolates | Indole Glucosinolates | ||||||
|---|---|---|---|---|---|---|---|---|
| GRA | SIN | GNA | GER | 4-HGBS | GBS | 4-MGBS | NGBS | |
| SB | 55.02 ± 2.96F | 11.5 ± 0.43F | 67.11 ± 5.55G | ND b | 1.74 ± 0.12B | 65.61 ± 2.71B | 2.18 ± 0.09IJ | 0.66 ± 0.07DE |
| LB | 13.29 ± 1.2G | 33.1 ± 1.22B | 764.74 ± 22.41B | ND | 0.74 ± 0.03D | 50.51 ± 1.89D | 4.02 ± 0.11DEF | 0.38 ± 0.02EF |
| QB1 | 63.93 ± 2.92F | 37.53 ± 1.43A | 1077.68 ± 96.73A | 0.081 ± 0.003CDE | 2.43 ± 0.22A | 55.49 ± 2.55DE | 4.06 ± 0.25DEF | 0.51 ± 0.06EF |
| CB | 49.64 ± 2.94F | 10.21 ± 0.95FG | 104.56 ± 6.45EFG | 0.001 ± 0.001E | 0.74 ± 0.03D | 17.92 ± 0.55F | 1.47 ± 0.06K | 0.11 ± 0.01F |
| XB | 110.08 ± 10.46E | 11.7 ± 0.77F | 264.32 ± 18.79D | 0.061 ± 0.002DE | 0.77 ± 0.01D | 24.64 ± 0.52E | 2.9 ± 0.29GH | 0.18 ± 0.02F |
| QB2 | 149.25 ± 8.33BC | 26.36 ± 1.63C | 809.65 ± 49.64B | 0.291 ± 0.218B | 2.34 ± 0.1A | 50.01 ± 1.33D | 3.62 ± 0.35EF | 0.2 ± 0.01F |
| DB | 22.07 ± 1.93G | 20.84 ± 0.74D | 119.36 ± 6.78EFG | ND | 0.2 ± 0.01F | 8.76 ± 0.48G | 4.49 ± 0.22BCD | 0.24 ± 0.01F |
| XLB | 10.48 ± 0.36G | 3.22 ± 0.1IJ | 33.42 ± 1.72G | ND | 0.21 ± 0.01F | 8.85 ± 0.3G | 1.62 ± 0.23JK | 0.08 ± 0.01F |
| TWDX | 137.26 ± 4.72CD | 5.54 ± 0.77HI | 49.98 ± 12.76G | 0.12 ± 0.004BCDE | 0.67 ± 0.09DE | 67.25 ± 1.81B | 3.82 ± 0.07EF | 1.82 ± 0.11C |
| KDSJ | 169.12 ± 4.07B | 17.19 ± 0.45C | 190.52 ± 27.65DEF | 0.235 ± 0.012BCD | 1.69 ± 0.06B | 77.36 ± 0.61A | 4.74 ± 0.17BC | 3.34 ± 0.35B |
| YC | 118.52 ± 9.43DE | 8.28 ± 0.68GH | 222.29 ± 9.23D | 0.092 ± 0.002CDE | 0.58 ± 0.03DE | 22 ± 0.78EF | 5.57 ± 0.13A | 0.97 ± 0.14D |
| DSHH | 49.74 ± 4.11F | 39.6 ± 1.58A | 194.48 ± 22.61DE | ND | 0.45 ± 0.03E | 18.95 ± 0.71F | 4.1 ± 0.08DE | 0.13 ± 0.01F |
| TZSJDS | 161.04 ± 9.01BC | 1.59 ± 0.05J | 92.82 ± 10.98FG | 0.868 ± 0.047A | 0.66 ± 0.07DE | 25.75 ± 0.51E | 3.41 ± 0.31FG | 0.68 ± 0.06DE |
| DSCH | 211.83 ± 14.51A | 15.72 ± 1.84E | 265.72 ± 19.78D | 0.265 ± 0.011BC | 0.78 ± 0.01D | 64.93 ± 4.14B | 5.01 ± 0.05AB | 10.75 ± 0.31A |
| ZH | 120.01 ± 2.16DE | 28.5 ± 0.77C | 629.1 ± 41.2C | 0.072 ± 0.004DE | 1.14 ± 0.02C | 62.26 ± 1.8B | 2.61 ± 0.24HI | 1.63 ± 0.09C |
| SJT | 201.88 ± 16.00A | 3.19 ± 0.21IJ | 38.41 ± 6.82G | 0.265 ± 0.05BC | 0.67 ± 0.07DE | 18.49 ± 0.42F | 4.26 ± 0.27CDE | 1.08 ± 0.02D |
| Abbreviation of Chinese Kale Varieties | Aliphatic Glucosinolates | Indole Glucosinolates | ||||||
|---|---|---|---|---|---|---|---|---|
| GRA | SIN | GNA | GER | 4-HGBS | GBS | 4-MGBS | NGBS | |
| SB | 194.34 ± 11.38ABC | 21.56 ± 2.12DE | 246.47 ± 10.75DE | 13.23 ± 0.62FG | 7.2 ± 0.11A | 34.28 ± 1.05B | 3.72 ± 0.08EFG | 2.06 ± 0.32C |
| LB | 63.49 ± 4.21I | 36.7 ± 4.68AB | 587.66 ± 45.52B | 2.28 ± 0.22J | 1.84 ± 0.03F | 17.32 ± 0.34GH | 2.76 ± 0.23I | 0.58 ± 0.11F |
| QB1 | 74.97 ± 3.76I | 29.43 ± 2.35BC | 972.53 ± 61.82A | 2.71 ± 0.2J | 1.76 ± 0.07F | 17.45 ± 0.31GH | 4.56 ± 0.17BCD | 0.91 ± 0.06EF |
| CB | 138.08 ± 8.23EFG | 15.83 ± 0.66EF | 151.6 ± 14.11FG | 8.46 ± 0.46H | 5.51 ± 0.17BC | 25.69 ± 0.21D | 3.23 ± 0.21FGHI | 1.72 ± 0.19CD |
| XB | 162 ± 10.81CDEF | 33.86 ± 1.62B | 429.69 ± 24.86C | 7.34 ± 0.2H | 4.75 ± 0.13C | 31.9 ± 0.57C | 2.04 ± 0.07J | 0.5 ± 0.01F |
| QB2 | 130.96 ± 7.38EF | 35.13 ± 4.81B | 417.33 ± 35.59C | 8.19 ± 0.18H | 3.14 ± 0.15E | 23.19 ± 0.65E | 4.6 ± 0.18BCD | 1.3 ± 0.05DE |
| DB | 90.47 ± 11.13HI | 43.18 ± 5.41A | 244.07 ± 23.02DE | 4.76 ± 0.25I | 3.38 ± 0.17DE | 15.48 ± 0.49H | 3.59 ± 0.14EFGH | 1.28 ± 0.14DE |
| XLB | 125.72 ± 11.69FGH | 15.88 ± 0.8EF | 156.14 ± 21.34FG | 7.55 ± 0.36H | 4.9 ± 1.21C | 26.64 ± 0.33D | 2.84 ± 0.13HI | 1.76 ± 0.1CD |
| TWDX | 147.53 ± 4.14DEF | 9.46 ± 0.52FG | 128.96 ± 7.87FG | 17.22 ± 0.61BC | 4.3 ± 0.31CDE | 30.48 ± 0.79C | 3.82 ± 0.02DEF | 6.58 ± 0.21A |
| KDSJ | 179.08 ± 11.23BCD | 29.69 ± 2.02BC | 202.29 ± 11.04EF | 12.02 ± 0.49G | 6.23 ± 0.59AB | 52.78 ± 1.31A | 4.11 ± 0.12CDE | 3.97 ± 0.06B |
| YC | 152.03 ± 14.14DEF | 8.61 ± 0.59FG | 114.62 ± 15.38FG | 16.51 ± 0.21CD | 6.22 ± 0.61AB | 19.98 ± 0.77F | 4.8 ± 0.29BC | 2.27 ± 0.6C |
| DSHH | 124.92 ± 7.39FGH | 24.67 ± 1.81CD | 74.86 ± 7.65G | 13.81 ± 0.37EF | 4.35 ± 0.33CED | 19.82 ± 0.46F | 3.66 ± 0.21EFG | 1.29 ± 0.08DE |
| TZSJDS | 127.82 ± 13.13FG | 1.28 ± 0.1G | 105.33 ± 7.97G | 15.27 ± 0.24DE | 4.6 ± 0.11CD | 24.84 ± 0.32DE | 2.92 ± 0.57GHI | 1.61 ± 0.09CD |
| DSCH | 173.57 ± 8.03BCDE | 33.52 ± 4.16B | 424.22 ± 25.7C | 15.62 ± 0.61CD | 5.18 ± 0.24BC | 34.77 ± 1.31B | 4.99 ± 0.35B | 3.95 ± 0.24B |
| ZH | 109.96 ± 12.88GH | 19.91 ± 0.55DE | 633.92 ± 50.2B | 16.49 ± 0.54CD | 1.47 ± 0.03F | 9.84 ± 0.16I | 6.34 ± 0.19A | 6.2 ± 0.17A |
| SJT | 201.05 ± 16.52AB | 3.57 ± 0.13G | 299.54 ± 10.45D | 27.47 ± 0.69A | 4.57 ± 0.07CD | 18.02 ± 0.34FG | 3.03 ± 0.36FGHI | 1.72 ± 0.18CD |
| Resources of Varieties of Chinese Kale and Measuring Site | Types of GLSs | Main Components of GLSs A-GLSs | The Average Content or Range of T-GLSs (the Proportion of A-GLSs) | Determination Method | References |
|---|---|---|---|---|---|
| 16 Chinese kale Varieties, bolting stems, edible leaves | A-: GRA, SIN, GNA, GER I-: GBS, 4-MGBS, NGBS, 4-HGBS | GNA, GRA, GBS | 1005.4 μmol/100 g DW (80.69–96.28%) | LC-MS | This study |
| 22 Chinese kale varieties, sprout | A-: GNA, SIN, GIB, PRO, GRA, GER, GAL, GNL I-: GBS, 4-MGBS, NGBS, 4-HGBS R-: GST | GNA, PRO, GIB, GBS | 5.51–104.57 μmol·g−1 DW (85–94%) | HPLC+GC-MS | [7] |
| 6 Chinese kale cultivars, mixed sample of stems leaves | A-: GNA, SIN, GIB, PRO, GRA, GER, GAL, GNL I-:GBS, 4-MGBS, NGBS, 4-HGBS AR-:GST | GNA, GER, GST, SIN, GBS | 18.93 μmol·g−1 DW (68.1%) | HPLC | [56] |
| 7 Chinese kale cultivars, edible part | A-: GNA, GRA, SIN, PRO, GER, 2-MBG, 3-MPG I-:GBS, 4-MGBS, NGBS, 4-HGBS R-:GST, GBR | GNA, GRA, SIN, GBS | --- | UHPLC-Quadrupole-Orbitrap MS/MS | [47] |
| 27 Chinese kale Varieties, sprout, rosette leaf, bolting stem | A-:GNA, SIN, GIB, PRO, GRA, GER, GAL, GNL I-:GBS, -MGBS, NGBS, 4-HGBS R-:GST | GNA, SIN, GIB, GBS | Sprout: 116.25 μmol·g−1 DW (92.5%) rosette leaf: 8.80 (74.0%) bolting stem: 11.61 (85.6%) | LC-MS | [21] |
| 10 Chinese kale Varieties, bolting stem | A-:GIB, PRO, GRA, SIN, GAL, GNA, GER I-: GBS,4-MGBS, NGBS, 4-HGBS | GNA, GRA, SIN, GBS | 8.38 μmol·g−1 DW (81.63%) | HPLC-PAD-ESI/MS | [41] |
| 43 Chinese kale Varieties, edible parts of leaves and flower stalks | A-:GNA, PRO, SIN, GRA, 4-pentenyl GLSs I-:GBS, 4-MGBS, NGBS, 4-HGBS | GNA, NGBS, GBS, PRO, SIN | 8.39 mg·g−1 DW (67.3%) | HPLC | [57] |
| 1 v Chinese kale variety, leaves, bolting stem | A-:GIB, PRO, SIN, GRA, GAL, GNA I-:GBS, 4-MGBS, NGBS, 4-HGBS | GNA, SIN, GIB | 1074.2 μg·g−1 DW in leaves (72.8%) 4907.1 μg·g−1 DW in stems (82.1%) | HPLC | [58] |
| 7 Chinese kale, bolting stem | A-:PRO, GRA, GNA I-:GBS, 4-MGBS, NGBS, 4-HGBS | GNA, GRA, NGBS, PRO, GBS | 412.47 μmol/100 g FW (72.08%) | HPLC | [20] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, L.; Cheng, S.; Lv, H.; Zhao, C.; Zhu, J.; Liu, P.; Wang, Z.; Yang, L.; Zhang, Y. Analysis of Glucosinolate Content, Composition and Expression Level of Biosynthesis Pathway Genes in Different Chinese Kale Varieties. Horticulturae 2021, 7, 398. https://doi.org/10.3390/horticulturae7100398
Tong L, Cheng S, Lv H, Zhao C, Zhu J, Liu P, Wang Z, Yang L, Zhang Y. Analysis of Glucosinolate Content, Composition and Expression Level of Biosynthesis Pathway Genes in Different Chinese Kale Varieties. Horticulturae. 2021; 7(10):398. https://doi.org/10.3390/horticulturae7100398
Chicago/Turabian StyleTong, Lu, Shanhan Cheng, Honghao Lv, Chengzhi Zhao, Jie Zhu, Pingwu Liu, Zhiwei Wang, Limei Yang, and Yangyong Zhang. 2021. "Analysis of Glucosinolate Content, Composition and Expression Level of Biosynthesis Pathway Genes in Different Chinese Kale Varieties" Horticulturae 7, no. 10: 398. https://doi.org/10.3390/horticulturae7100398
APA StyleTong, L., Cheng, S., Lv, H., Zhao, C., Zhu, J., Liu, P., Wang, Z., Yang, L., & Zhang, Y. (2021). Analysis of Glucosinolate Content, Composition and Expression Level of Biosynthesis Pathway Genes in Different Chinese Kale Varieties. Horticulturae, 7(10), 398. https://doi.org/10.3390/horticulturae7100398