
Exogenously Applied Chitosan and Chitosan Nanoparticles Improved Apple Fruit Resistance to Blue Mold, Upregulated Defense-Related Genes Expression, and Maintained Fruit Quality


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Pathogen and Pathogenicity Test
2.2. Conidial Suspension
2.3. Source of Compounds
2.4. Characterization of Chitosan Nanoparticles
2.5. Antifungal Effect of the Tested Chemical on P. expansum Growth In Vitro
- Xc: Mean of colony diameter in control plates.
- Xi: Mean of colony diameter of test plates.
2.6. The Effect of Pre-Harvest Spray with Tested Chemicals on the Development of Blue Mold on Apples under Storage Conditions
2.6.1. Field Application
2.6.2. Artificial Infection Experiments
2.7. Assessments
2.7.1. Disease Incidence
2.7.2. Natural Infection Experiments
2.7.3. Fruit Firmness (lb/in2)
2.7.4. Fruit Total Soluble Solids
2.7.5. Fruit Titratable Acidity
2.8. Total RNA Isolation and cDNA Synthesis
2.9. Differential Expression Analysis Using Quantitative Real-Time PCR
2.10. Statistical Analysis
3. Results
3.1. Antifungal Effect of Some Chemical on the Colony Growth of P. expansum In Vitro
3.2. Effect of Pre-Harvest Spray with Tested Chemicals on the Development of Blue Mold on Apples under Storage Condition
3.3. Effects of the Tested Chemicals on the Quality of Postharvest Apples
Fruit Firmness
3.4. Fruit Total Soluble Solids
3.5. Fruit Titratable Acidity
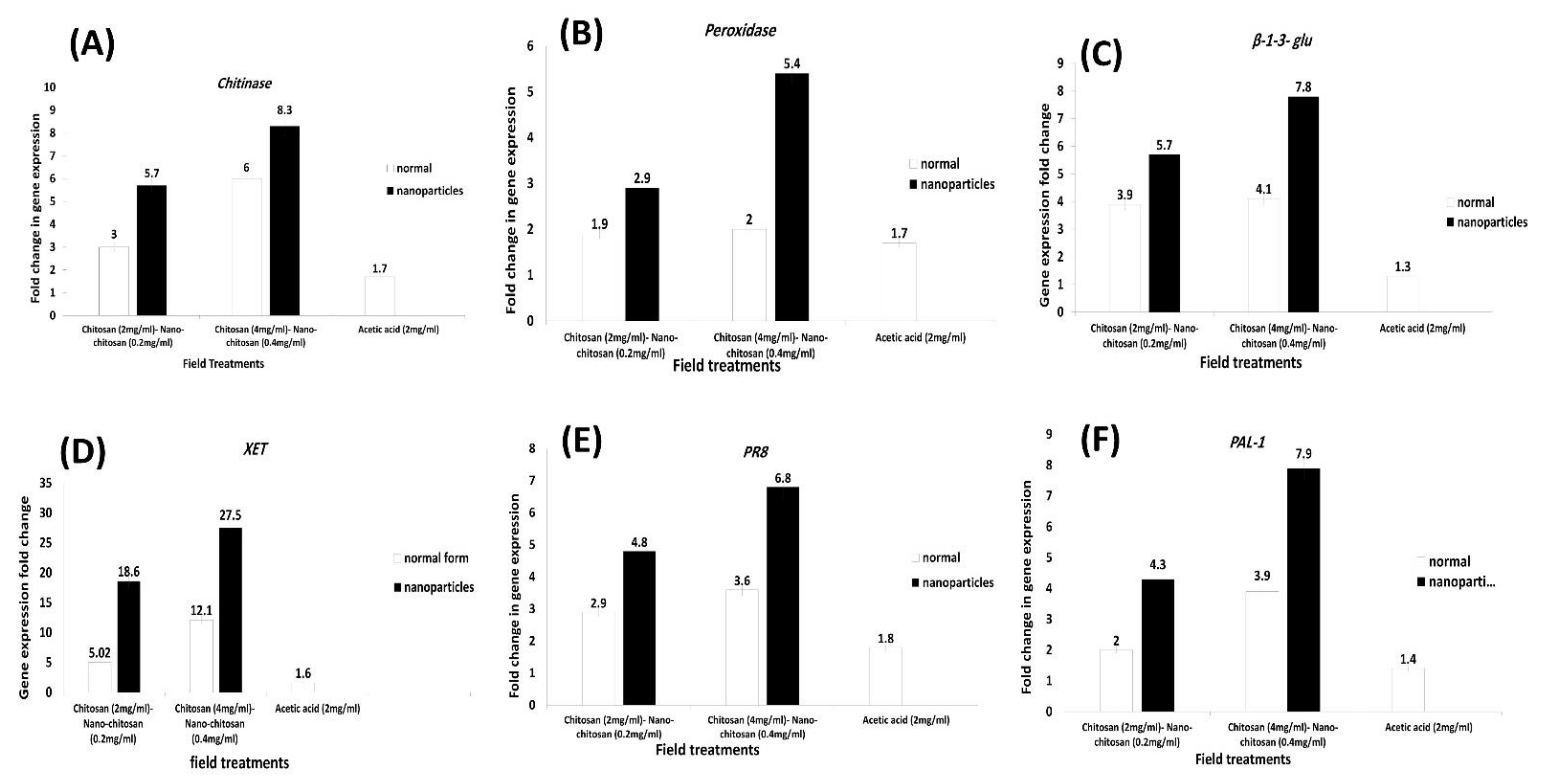
3.6. Expression Profiling of Some Defense-Related Genes in Leaf Samples of Apple Trees (cv. Anna) Responding to Application with Different Treatments
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT 2020. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 29 October 2020).
- Errampalli, D. Penicillium expansum (Blue Mold). In Postharvest Decay; Bautista-Baños, S., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 189–231. [Google Scholar] [CrossRef]
- Larous, L.; Hendel, N.; Abood, J.K.; Ghoul, M. The growth and production of patulin mycotoxin by Penicillium expansum on apple fruits and its control by the use of propionic acid and sodium benzoate. Arab J. Plant Prot. 2007, 25, 123–128. [Google Scholar]
- Zhong, L.; Carere, J.; Lu, Z.; Lu, F.; Zhou, T. Patulin in Apples and Apple-Based Food Products: The Burdens and the Mitigation Strategies. Toxins 2018, 10, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, H.; Marín, S.; Rovira, A.; Ramos, A.J.; Sanchis, V. Patulin accumulation in apples by Penicillium expansum during postharvest stages. Lett. Appl. Microbiol. 2007, 44, 30–35. [Google Scholar] [CrossRef]
- Torres, R.; Teixidó, N.; Viñas, I.; Mari, M.; Casalini, L.; Giraud, M.; Usall, J. Efficacy of Candida sake CPA-1 formulation for controlling Penicillium expansum decay on pome fruit from different Mediterranean regions. J. Food Prot. 2006, 69, 2703–2711. [Google Scholar] [CrossRef]
- Prusky, D.; McEvoy, J.L.; Saftner, R.; Conway, W.S.; Jones, R. Relationship between host acidification and virulence of Penicillium spp. on apple and citrus fruit. Phytopathology 2004, 94, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Papoutsis, K.; Mathioudakis, M.M.; Hasperué, J.H.; Ziogas, V. Non-chemical treatments for preventing the postharvest fungal rotting of citrus caused by Penicillium digitatum (green mold) and Penicillium italicum (blue mold). Trends Food Sci. Technol. 2019, 86, 479–491. [Google Scholar] [CrossRef]
- Hashim, A.F.; Youssef, K.; Abd-Elsalam, K.A. Ecofriendly nanomaterials for controlling gray mold of table grapes and maintaining postharvest quality. Eur. J. Plant Pathol. 2019, 154, 377–388. [Google Scholar] [CrossRef]
- Gardesh, A.S.K.; Badii, F.; Hashemi, M.; Ardakani, A.Y.; Maftoonazad, N.; Gorji, A.M. Effect of nanochitosanbased coating on climacteric behavior and post-harvest shelf-life extension of apple cv. Golab Kohanz. LWT 2016, 70, 33–40. [Google Scholar] [CrossRef]
- Pilon, L.; Spricigo, P.C.; Miranda, M.; de Moura, M.R.; Assis, O.B.G.; Mattoso, L.H.C.; Ferreira, M.D. Chitosannanoparticle coatings reduce microbial growth on fresh-cut apples while not aecting quality attributes. Int. J. Food Sci. Technol. 2015, 50, 440–448. [Google Scholar] [CrossRef]
- Youssef, K.; Hashim, A.F. Inhibitory effect of clay/chitosan nanocomposite against Penicillium digitatum on citrus and its possible mode of action. Jordan J. Biol. Sci. 2020, 13, 349–355. [Google Scholar]
- Pichyangkura, R.; Chatchawan, S. Bio stimulant activity of chitosan in horticulture. Sci. Hortic. 2015, 195, 49–65. [Google Scholar] [CrossRef]
- El-Garhy, H.A.; Abdel-Rahman, F.A.; Shams, A.S.; Osman, G.H.; Moustafa, M. Comparative analyses of four chemicals used to control black mold disease in tomato and its effects on defense signaling pathways, productivity and quality traits. Plants 2020, 9, 808. [Google Scholar] [CrossRef]
- Abdel-Rahman, F.A.; Eman, Y.K.; Maali, S.S.; Tahsin, S.; Yosra, A. Preharvest application of salicylic acid induces some resistant genes of sweet pepper against black mold disease. Eur. J. Plant Pathol. 2021, 159, 755–768. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, X.; Li, B.; Zhang, Q.; Liang, W.; Wang, C. Salicylic acid confers enhanced resistance to Glomerella leaf spot in apple. Plant Physiol. Biochem. 2016, 106, 64–72. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- González-Candelas, L.; Alamar, S.; Sánchez-Torres, P.; Zacarias, L.; Marcos, J.F. A transcriptomic approach highlights induction ofsecondary metabolism in citrus fruit in response to Penicillium digitatum infection. BMC Plant Biol. 2010, 10, 194–211. [Google Scholar] [CrossRef] [Green Version]
- Vilanova, L.; Wisniewski, M.; Norelli, J.; Viñas, I.; Torres, R.; Usall, J.; Phillips, J.; Droby, S.; Teixidó, N. Transcriptomic profiling of apple in response to inoculation with a pathogen (Penicillium expansum) and a non-pathogen (Penicillium digitatum). Plant Mol. Biol. Rep. 2014, 32, 566–583. [Google Scholar] [CrossRef]
- Samson, R.A.; Hoekstra, E.S.; Frisvad, J.C.; Filtenborg, O. (Eds.) Introduction to Food- and Airborne Fungi, 6th ed.; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2002; p. 389. [Google Scholar]
- Jabnoun-Khiareddine, H.; El-Mohamedy, R.S.R.; Abdel-Kareem, F.; Aydi Ben Abdallah, R.; Gueddes-Chahed, M.; Daami-Remadi, M. Variation in chitosan and salicylic acid efficacy towards soil-borne and air-borne fungi and their suppressive effect of tomato wilt severity. J. Plant Pathol. Microbiol. 2015, 6, 325. [Google Scholar] [CrossRef] [Green Version]
- Algam, S.A.E.; Xie, G.; Li, B.; Yu, S.; Su, T.; Larsen, J. Effects of Paenibacillus strains and chitosan on plant growth promotion and control of Ralstonia wilt in tomato. J. Plant Pathol. 2010, 92, 593–600. [Google Scholar]
- Guo, Z.; Chen, R.; Xing, R.; Liu, S.; Yu, H.; Wang, P.; Li, C.; Li, P. Novel derivatives of chitosan and their antifungal activities in-vitro. Carbohydr. Res. 2006, 341, 351–354. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley and Sons: New York, NY, USA, 1984; p. 680. [Google Scholar]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- El-Garhy, H.A.S.; Elsisi, A.A.; Mohamed, S.A.; Morsy, O.M.; Osman, G.; Abdel-Rahman, F.A. Transcriptomic changes in green bean pods against grey mould and white rot diseases via field application of chemical elicitor nanoparticles. IET Nanobiotechnol. 2020, 14, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Esmailzadeh, M.; Soleimani, M.J.; Rouhani, H. Exogenous application of salicylic acid for inducing systemic acquired resistance against tomato stem canker disease. J. Biol. Sci. 2008, 8, 1039–1044. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Chen, C.; Xia, X.; Garba, B.; Shang, L.; Wang, Y. Proteomic analysis of the inhibitory effect of chitosan on Penicillium expansum. Food Sci. Technol. 2020, 40, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Alghuthaymi, M.A.; Abd-Elsalam, K.A.; Shami, A.; Said-Galive, E.; Shtykova, E.V.; Naumkin, A.V. Silver/chitosan nanocomposites: Preparation and characterization and their fungicidal activity against dairy cattle toxicosis Penicillium expansum. J. Fungi 2020, 6, 51. [Google Scholar] [CrossRef] [Green Version]
- Chookhongkha, N.; Sopondilok, T.; Photchanachai, S. Effect of chitosan and chitosan nanoparticles on fungal growth and chilli seed quality. Acta Hortic. 2013, 973, 231–237. [Google Scholar] [CrossRef]
- Raliya, R.; Tarafdar, J.C.; Singh, S.K.; Gautam, R.; Choudhary, K.; Maurino, V.G.; Saharan, V. MgO nanoparticles biosynthesis and its effect on chlorophyll contents in the leaves of clusterbean (Cyamopsis tetragonoloba L.). Adv. Sci. Eng. Med. 2014, 6, 538–545. [Google Scholar] [CrossRef]
- Agnihotri, S.A.; Mallikarjuna, N.N.; Aminabhavi, T.M. Recent advances in chitosan-based micro-and nanoparticles in drug delivery. J. Control. Rel. 2004, 100, 5–28. [Google Scholar] [CrossRef]
- Momin, J.K.; Jayakumar, C.; Prajapati, J.B. Potential of nanotechnology in functional foods. Emir. J. Food Agric. 2013, 25, 10–19. [Google Scholar] [CrossRef]
- Bautista-Baños, S.; Hernández-López, M.; Bosquez-Molina, E. Growth inhibition of selected fungi by chitosan and plant extracts. Rev. Mex. Fitopatol. 2004, 22, 178–186. [Google Scholar]
- Abd-El-Kareem, F.; Haggag, W.M.; Saied, N.M.; Elshahawy, I.E. Postharvest application of acetic acid vapours and chitosan solution for controlling gray and blue moulds of apple fruits. J. Chem. Pharm. Res. 2015, 7, 581–588. [Google Scholar]
- Abdel-Rahman, F.A. Safe Technologies For Controlling Post-Harvest Diseases of Green Bean Prepared For Exportation. Ph.D. Thesis, Faculty of Agriculture, Benha University, Benha, Egypt, 2015; p. 250. [Google Scholar]
- Zeb, S.; Muhammad, S.; Syed, T.; Mazhar, A.; Shahid, A.; Zohra, N.; Kamran, R.; Aamir, K.; Hajra, S.; Syed, A.; et al. Influence of post-harvest application of chitosan on physico-chemical changes of apple fruit during storage. Pure Appl. Biol. 2020, 9, 2554–2562. [Google Scholar] [CrossRef]
- Ranjbar, S.; Rahemi, M.; Ramezanian, A. Comparison of nano-calcium and calcium chloride spray on post-harvest quality and cell wall enzymes activity in apple cv. Red Delicious. Sci. Hortic. 2018, 240, 57–64. [Google Scholar] [CrossRef]
- Moussa, S.H.; Tayel, A.A.; Alsohim, A.S.; Abdallah, R.R. Botryticidal growth of nanosized silver–chitosan composite and its application for the control of gray mold in strawberry. J. Food Sci. 2013, 78, 1589–1594. [Google Scholar] [CrossRef]
- Youssef, K.; de Oliveira, A.G.; Tischer, C.A.; Hussain, I.; Roberto, S.R. Synergistic effect of a novel chitosan/silica nanocomposites-based formulation against gray mold of table grapes and its possible mode of action. Int. J. Biol. Macromol. 2019, 141, 247–258. [Google Scholar] [CrossRef]
- Liu, K.; Liu, J.; Li, H.; Yuan, C.; Zhong, J.; Chen, Y. Influence of postharvest citric acid and chitosan coating treatment onripening attributes and expression of cell wall related genes incherimoya (Annona cherimola Mill.) fruit. Sci. Hortic. 2016, 198, 1–11. [Google Scholar] [CrossRef]
- Miedes, E.; Lorences, E.P. Apple (Malus domestica) and tomato (Lycopersicum esculentum) fruits cell-wall hemicelluloses and xyloglucan degradation during Penicillium expansum infection. J. Agric. Food Chem. 2004, 52, 7957–7963. [Google Scholar] [CrossRef]
- De la Cova, C.; Townley, R.; Regot, S.; Greenwald, I.A. Real-time biosensor for ERK activity reveals signaling dynamics during C. elegans cell fate specification. Dev. Cell 2017, 42, 542–553.e4. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.P.; Yao, H.J.; Deng, X.; Xu, X.B.; Qin, G.Z.; Chan, Z.L. Characterization and expression of beta-1,3-glucanase genes in jujube fruit induced by the microbial biocontrol agent Cryptococcus laurentii. Phytopathology 2007, 97, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Wang, P.; Li, M.; Ke, X.; Li, C.; Liang, D.; Wu, S.; Ma, X.; Li, C.; Zou, Y.; et al. Exogenous melatonin improves Malus resistance to Marssonina apple blotch. J. Pineal Res. 2013, 54, 426–434. [Google Scholar] [CrossRef]
- El-Garhy, H.A.S.; Rashid, I.A.S.; Abou-Ali, R.M.; Moustafa, M.M.A. Field application of safe chemical elicitors induced the expression of some resistance genes against grey mold and cottony rot diseases during snap bean pods storage. Gene 2016, 576, 358–365. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Gene Name | Accession No. | Oligonucleotide Sequence (5–3) | Reference |
|---|---|---|---|
| PR8 | DQ318214 | F-CCACTGCAACCCCGCTAGT’ | Zhang 2016 |
| R-CGGGCGCGAATCTGACTGA | |||
| Chitinase (Chi) | FJ422811 | F-GGGTGAGTTGGATGGGTC | Zhang 2016 |
| R-TGGAGGATGGGAAAGTGC | |||
| β-1,3-glucanase (β-1,3-glu) | FJ598140 | F-TGCCGTAGGAAACGAAAT | Zhang 2016 |
| R-TGATGGAGGAAAGGAATT | |||
| Xyloglucan endotransglycosylase (XET) | XM008370279 | F-TGCTCTCTGTTTCCCACACCAT | Cova et al., 2017 |
| R-GAACTCAGACGGTCCACTACACGATGAG5TC | |||
| Phenylalanine ammonia lyase-1 (PAL1) | AT2G37040 | F-TTTGTGAGGGAGGAGTTGG | Vilanova et al., 2014 |
| R-GTTGAATGTGAAGGAATGCAG | |||
| Peroxidase | AT5G42180 | F-CCTCCTCAAAACAAGCGTTC | Vilanova et al., 2014 |
| R-CACAGCACACACAAACAATTTC | |||
| Elongation Factor (EF-1α) | XM008387060 | F-GGTCATTGGTCATGTCGACTCTGG | Cova et al., 2017 |
| (reference gene) | R-GCACCCAGGCATACTTGAATGACC |
| Treatments | Con. | Linear Growth | |
|---|---|---|---|
| Mm | Ef | ||
| Chitosan | 2 g/L | 76.50 b | 15 |
| 4 g/L | 68.53 c | 23.86 | |
| Nano-Chitosan | 0.2 g/L | 61.45 d | 31.72 |
| 0.4 g/L | 50.11 e | 44.32 | |
| Acetic acid | 2 mL/L | 0.00 f | 100 |
| Control | 90.00 a | ||
| Treatments | Con. | Season 2019 | Season 2020 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Artificial Infection | Natural Infection | Artificial Infection | Natural Infection | ||||||
| D.I (%) | EF (%) | D.I (%) | EF (%) | D.I (%) | EF (%) | D.I (%) | EF (%) | ||
| Chitosan | 2 g/L | 51.76 b | 38.95 | 11.34 b | 70.97 | 49.07 b | 40.61 | 9.27 c | 74.44 |
| 4 g/L | 44.26 c | 47.81 | 6.53 c | 83.27 | 39.78 d | 51.86 | 5.28 d | 85.43 | |
| Nano-chitosan | 0.2 g/L | 36.30 d | 57.19 | 6.85 c | 82.45 | 34.57 e | 58.16 | 5.40 d | 85.11 |
| 0.4 g/L | 30.77 e | 63.71 | 1.73 d | 95.56 | 28.82 f | 65.12 | 1.33 e | 96.33 | |
| Acetic acid | 2 m/L | 45.02 c | 46.91 | 14.28 b | 63.44 | 46.03 c | 44.29 | 12.51 b | 65.49 |
| Control (water) | - | 84.79 a | - | 39.05 a | - | 82.63 a | - | 36.25 a | - |
| Treatments | Con. mg/mL | Season 2019 | Season 2020 | ||
|---|---|---|---|---|---|
| Artificial Infection | Natural Infection | Artificial Infection | Natural Infection | ||
| Chitosan | 2 g/L | 7.9 bc | 10.0 b | 8.1 cd | 10.5 a |
| 4 g/L | 8.2 b | 9.0 c | 8.6 b | 9.4 b | |
| Nano-chitosan | 0.2 g/L | 9.1 a | 10.7 a | 9.4 a | 10.9 a |
| 0.4 g/L | 8.8 a | 9.3 c | 9.2 a | 9.5 b | |
| Acetic acid | 2 m/L | 8.0 b | 9.2 c | 8.5 bc | 9.2 b |
| Control | 7.5 c | 8.1 d | 7.7 d | 8.0 c | |
| Treatments | Con. | Season 2019 | Season 2020 | ||
|---|---|---|---|---|---|
| Artificial Infection | Natural Infection | Artificial Infection | Natural Infection | ||
| Chitosan | 2 g/L | 10.1 a | 11.0 a | 10.6 a | 10.7 a b |
| 4 g/L | 8.2 c | 10.2 c | 8.7 de | 10.4 b | |
| Nano-chitosan | 0.2 g/L | 8.6 b | 10.6 b | 9.1 cd | 10.9 a |
| 0.4 g/L | 9.0 b | 8.8 e | 9.6 b | 8.4 d | |
| Acetic acid | 2 m/L | 8.9 b | 9.2 d | 9.2 bc | 9.7 c |
| Control | 8.1 c | 10.0 c | 8.5 e | 10.3 b | |
| Treatments | Con. | Season 2019 | Season 2020 | ||
|---|---|---|---|---|---|
| Artificial Infection | Natural Infection | Artificial Infection | Natural Infection | ||
| Chitosan | 2 g/L | 0.31 d | 0.72 a | 0.34 d | 0.74 a |
| 4 g/L | 0.52 c | 0.61 c | 0.54 c | 0.65 b | |
| Nano-chitosan | 0.2 g/L | 0.57 b | 0.57 d | 0.65 a | 0.58 c |
| 0.4 g/L | 0.64 a | 0.63 bc | 0.67 a | 0.62 b | |
| Acetic acid | 2 m/L | 0.59 b | 0.66 b | 0.6 b | 0.63 b |
| Control | 0.33 d | 0.46 e | 0.35 d | 0.43 d | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Rahman, F.A.; Monir, G.A.; Hassan, M.S.S.; Ahmed, Y.; Refaat, M.H.; Ismail, I.A.; El-Garhy, H.A.S. Exogenously Applied Chitosan and Chitosan Nanoparticles Improved Apple Fruit Resistance to Blue Mold, Upregulated Defense-Related Genes Expression, and Maintained Fruit Quality. Horticulturae 2021, 7, 224. https://doi.org/10.3390/horticulturae7080224
Abdel-Rahman FA, Monir GA, Hassan MSS, Ahmed Y, Refaat MH, Ismail IA, El-Garhy HAS. Exogenously Applied Chitosan and Chitosan Nanoparticles Improved Apple Fruit Resistance to Blue Mold, Upregulated Defense-Related Genes Expression, and Maintained Fruit Quality. Horticulturae. 2021; 7(8):224. https://doi.org/10.3390/horticulturae7080224
Chicago/Turabian StyleAbdel-Rahman, Fayz A., Gehan A. Monir, Mabrouk S. S. Hassan, Yosra Ahmed, Mohamed H. Refaat, Ismail A. Ismail, and Hoda A. S. El-Garhy. 2021. "Exogenously Applied Chitosan and Chitosan Nanoparticles Improved Apple Fruit Resistance to Blue Mold, Upregulated Defense-Related Genes Expression, and Maintained Fruit Quality" Horticulturae 7, no. 8: 224. https://doi.org/10.3390/horticulturae7080224
APA StyleAbdel-Rahman, F. A., Monir, G. A., Hassan, M. S. S., Ahmed, Y., Refaat, M. H., Ismail, I. A., & El-Garhy, H. A. S. (2021). Exogenously Applied Chitosan and Chitosan Nanoparticles Improved Apple Fruit Resistance to Blue Mold, Upregulated Defense-Related Genes Expression, and Maintained Fruit Quality. Horticulturae, 7(8), 224. https://doi.org/10.3390/horticulturae7080224