
Post-Harvest Non-Conventional and Traditional Methods to Control Cadophora luteo-olivacea: Skin Pitting Agent of Actinidia chinensis var. deliciosa (A. Chev.)




Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit
2.2. Biocontrol Agents and Pathogens
2.3. Chemical and Natural Substances
2.4. In Vitro Assays with Antagonists
2.5. In Vitro Assays: Amended Medium and Brassica nigra Defatted Meal Biofumigation
2.6. In Vivo Assays
2.7. FT-IR Spectroscopy
2.8. Statistical Analysis
3. Results
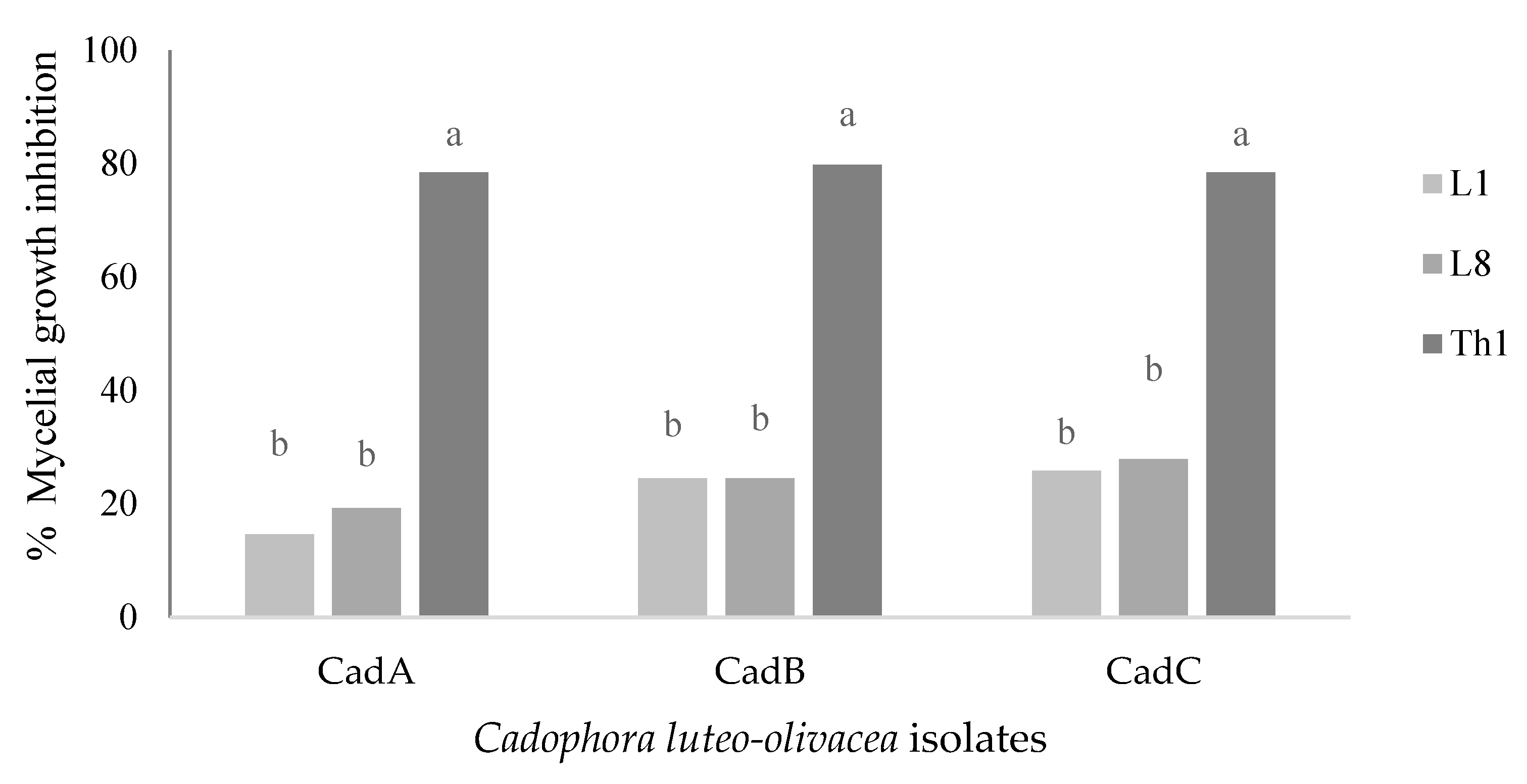
3.1. In Vitro Assays with Antagonists
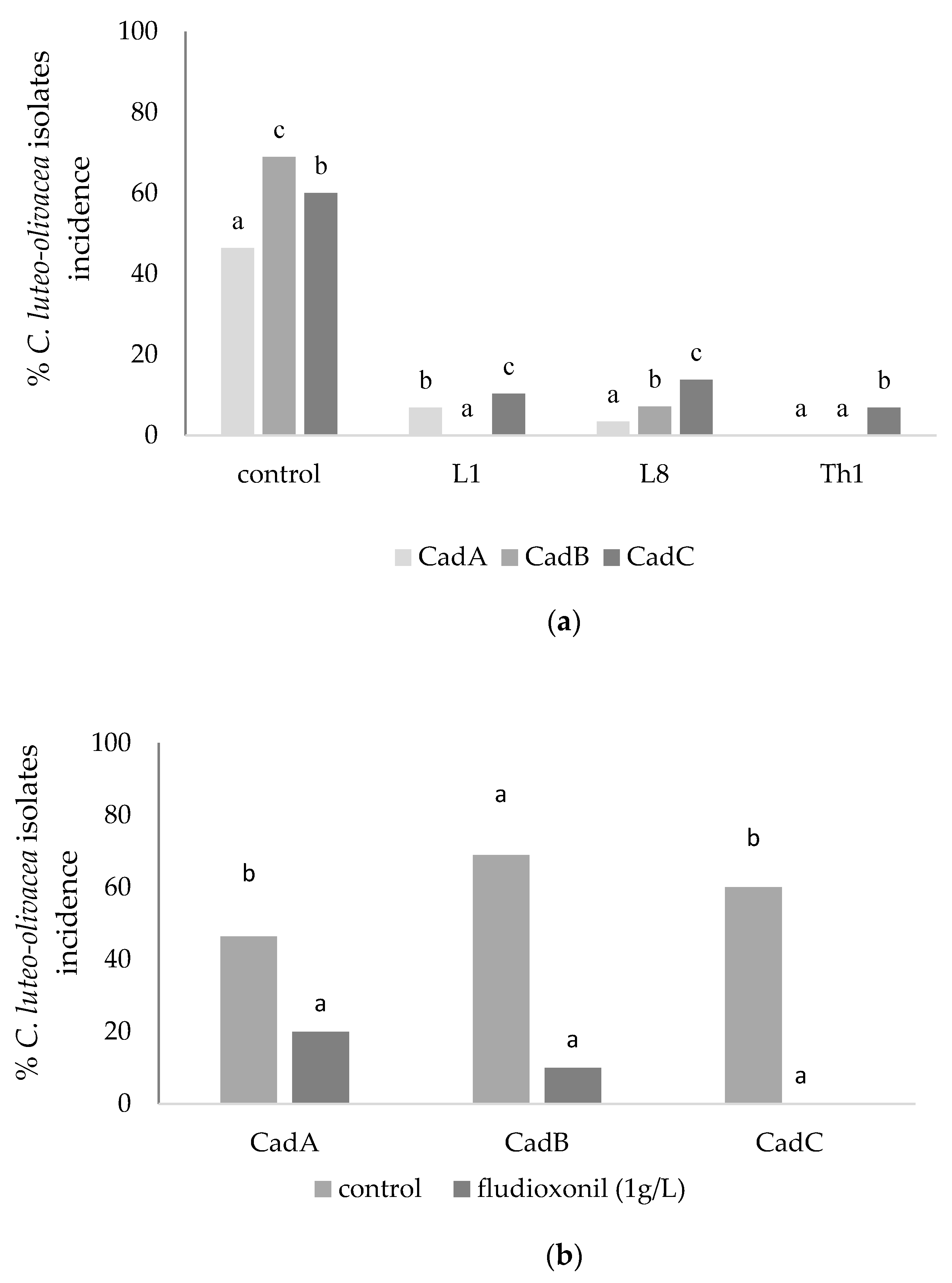
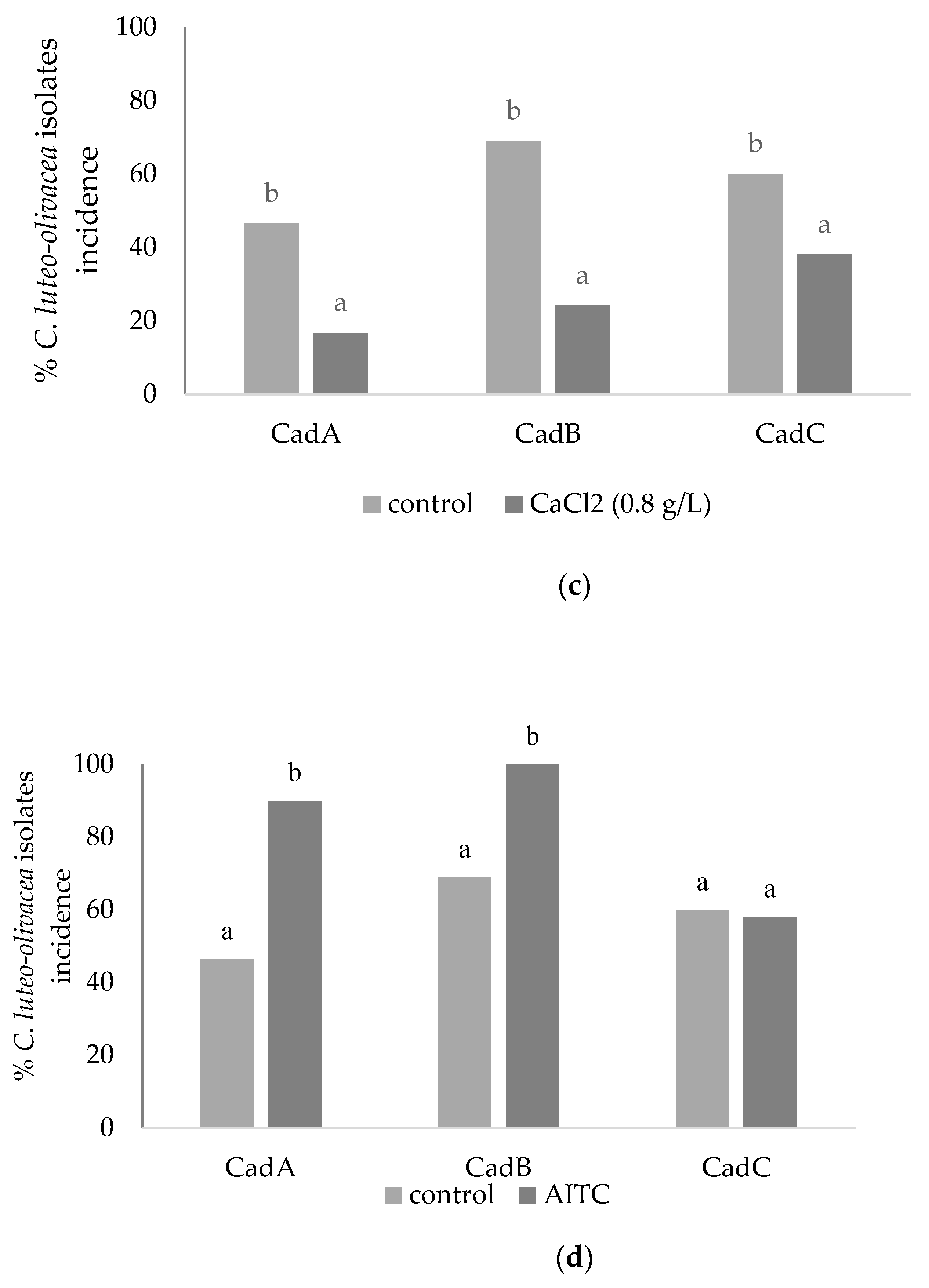
3.2. In Vitro Assays: Amended Medium (Fludioxonil and CaCl2) and Brassica nigra Defatted Meal Biofumigation
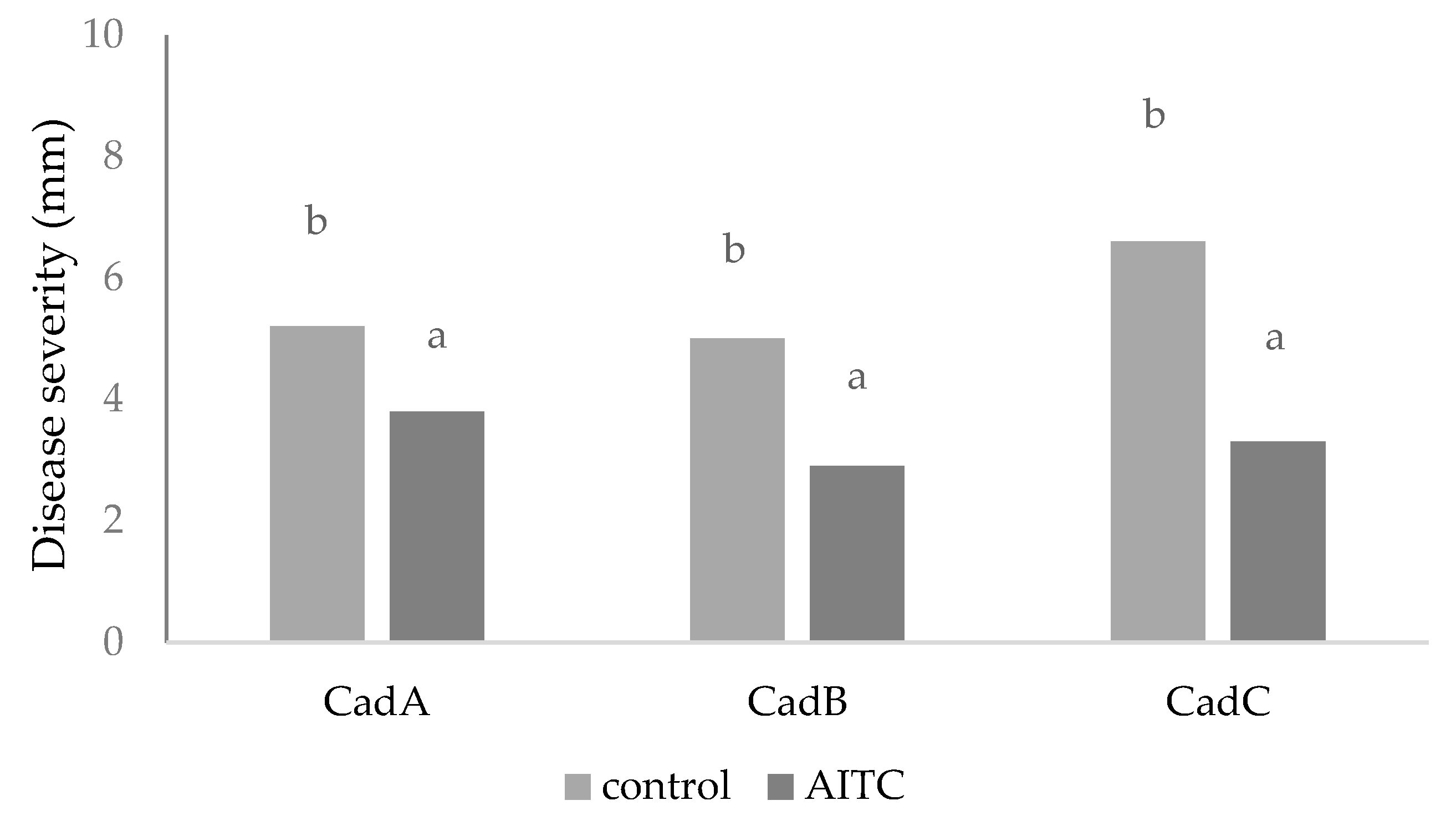
3.3. In Vivo Assays
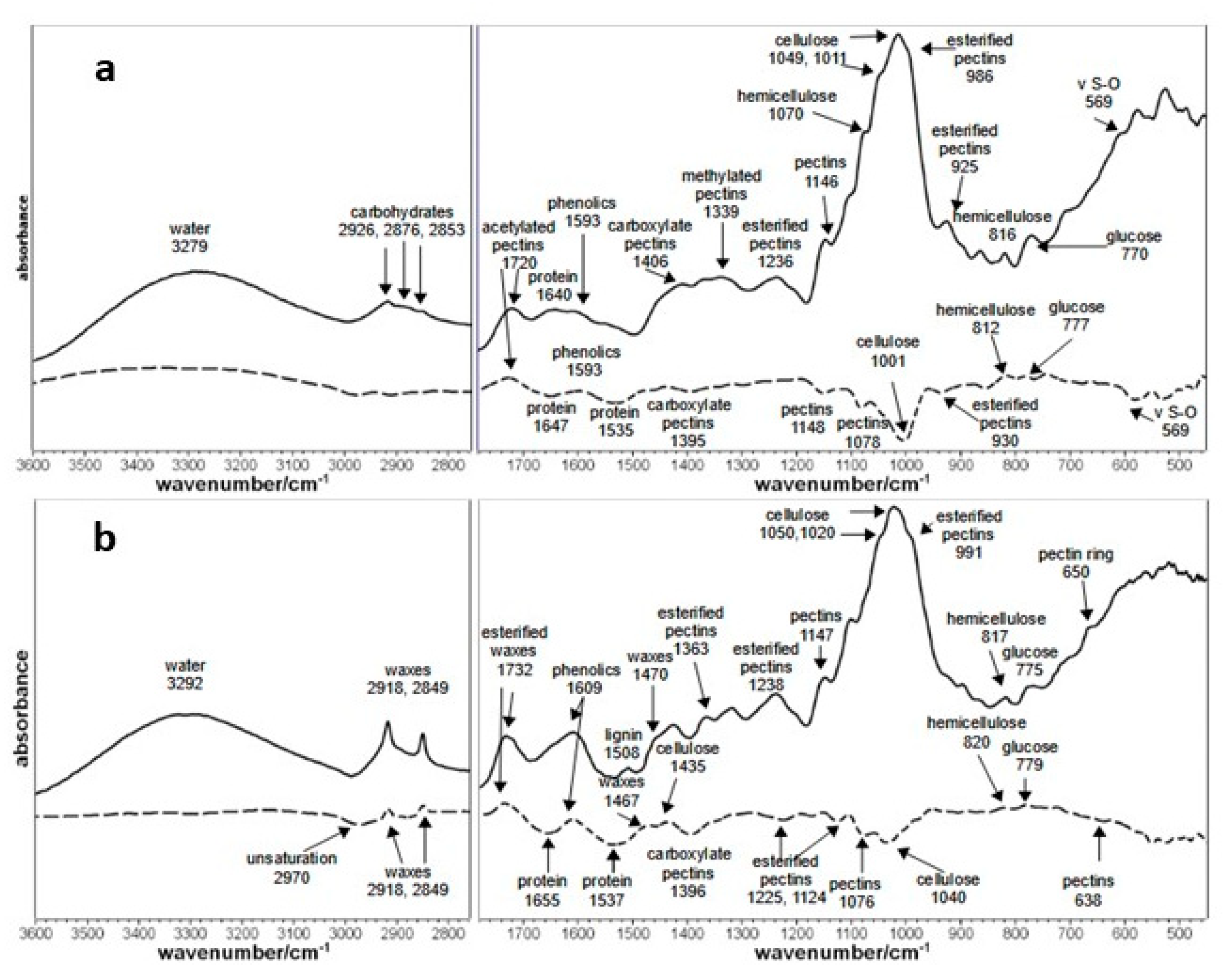
3.4. FT-IR Kiwifruit Analysis
4. Discussion
Supplementary Materials
Author Contributions
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gramaje, D.; Mostert, L.; Rmengol, J. Characterization of Cadophora luteo-olivacea and C. melinii isolates obtained from grapevines and environmental samples from grapevine nurseries in Spain. Phytopathol. Mediterr. 2011, 50, 112–126. [Google Scholar]
- Khol, J.; Wenneker, M.; Groenenboom-de Haas, B.H.; Anbergen, R.; Goossen-van de Geijn, H.M.; Lombaers-van der Plas, C.H.; Pinto, F.A.M.F.; Kastelein, P. Dynamics of post-harvest pathogens Neofabraea spp. And Cadophora spp. in plant residues in Dutch apple and pear orchards. Plant Pathol. 2018, 67, 1264–1277. [Google Scholar] [CrossRef] [Green Version]
- Spadaro, D.; Galliano, A.; Pellegrino, C.; Gilardi, G.; Garibaldi, A.; Gullino, M.L. Dry matter and mineral composition, together with commercial storage practices, influence the development of skin pitting caused by Cadophora luteo-olivacea on kiwifruit ‘Hayward’. J. Plant Pathol. 2010, 92, 339–346. [Google Scholar]
- Snowdon, A.L. A Color Atlas of Post-Harvest Diseases and Disorders of Fruits and Vegetables General Introduction and Fruits; CRC Press: Boca Raton, FL, USA, 1990; Volume 1. [Google Scholar]
- Di Francesco, A.; Mari, M.; Ugolini, L.; Baraldi, E. Effect of Aureobasidium pullulans strains against Botrytis cinerea on kiwifruit during storage and on fruit nutritional composition. Food Microbiol. 2018, 72, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.W.M.; Long, P.G.; Ganesh, S. The combined effect of delayed application of yeast biocontrol agents and fruit curing for the inhibition of the post-harvest pathogen Botrytis cinerea in kiwifruit. Postharvest Biol. Technol. 1999, 16, 233–243. [Google Scholar] [CrossRef]
- Cheng, L.; Nie, X.; Jiang, C.; Li, S. The combined use of the antagonistic yeast Hanseniaspora uvarum with β-aminobutyric acid for the management of post-harvest diseases of kiwifruit. Biol. Control 2019, 137, 104019. [Google Scholar] [CrossRef]
- Sharma, R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Mari, M.; Leoni, O.; Bernardi, R.; Neri, F.; Palmieri, S. Control of brown rot on stonefruit by synthetic and glucosinolate-derived isothiocyanates. Postharvest Biol. Technol. 2008, 47, 61–67. [Google Scholar] [CrossRef]
- Ugolini, L.; Martini, C.; Lazzeri, L.; D’Avino, L.; Mari, M. Control of post-harvest grey mould (Botrytis cinerea Per.: Fr.) on strawberries by glucosinolate-derived allyl-isothiocyanate treatments. Postharvest Biol. Technol. 2014, 90, 34–39. [Google Scholar] [CrossRef]
- Chen, H.; Gao, H.; Fang, X.Y.L.; Zhou, Y.; Yang, H. Effects of allyl isothiocyanate treatment on post-harvest quality and the activities of antioxidant enzymes of mulberry fruit. Postharvest Biol. 2015, 108, 61–67. [Google Scholar] [CrossRef]
- Thomidis, T.; Prodromu, I. Evaluation of the effectiveness of three fungicides against pathogens causing post-harvest fruit rot of kiwifruit. Australas. Plant Pathol. 2018, 47, 485–489. [Google Scholar] [CrossRef]
- Conway, W.S. The effects of post-harvest infiltration of calcium, magnesium or strontium on decay, firmness, respiration and ethylene production in apples. J. Am. Soc. Hortic. Sci. 1987, 112, 300–303. [Google Scholar]
- Mari, M.; Spadoni, A.; Ceredi, G. Alternative technologies to control postharvest diseases of kiwifruit. Stewart Postharvest Rev. 2015, 11, 1–5. [Google Scholar]
- Di Francesco, A.; Calassanzio, M.; Ratti, C.; Mari, M.; Folchi, A.; Baraldi, E. Molecular characterization of the two post-harvest biological control agents Aureobasidium pullulans L1 and L8. Biol. Control 2018, 123, 53–59. [Google Scholar] [CrossRef]
- Di Francesco, A.; Milella, F.; Mari, M.; Roberti, R. A preliminary investigation into Aureobasidium pullulans as a potential biocontrol agent against Phytophthora infestans of tomato. Biol. Control 2017, 114, 144–149. [Google Scholar] [CrossRef]
- Bonora, S.; Francioso, O.; Tugnoli, V.; Prodi, A.; Di Foggia, M.; Righi, V.; Nipoti, P.; Filippini, G.; Pisi, A. Structural characteristics of “Hayward” kiwifruit from elephantiasis-affected plants studied by DRIFT, FT-Raman, NMR and SEM techniques. J. Agric. Food Chem. 2009, 57, 4827–4832. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Di Foggia, M.; Baraldi, E. Aureobasidium pullulans volatile organic compounds as alternative post-harvest method to control brown rot of stone fruits. Food Microbiol. 2020, 87, 103395. [Google Scholar] [CrossRef]
- Ugolini, L.; Righetti, L.; Carbone, K.; Paris, R.; Malaguti, L.; Di Francesco, A.; Micheli, L.; Paliotta, M.; Mari, M.; Lazzeri, L. Postharvest application of brassica meal-derived allyl-isothiocyanate to kiwifruit: Effect on fruit quality, nutraceutical parameters and physiological response. J. Food Sci. Technol. 2017, 54, 751–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Ghaouth, A.; Wilson, C.L. Biologically-based technol-ogies for the control of post-harvest diseases. Postharvest News Inf. 1995, 6, 5N–11N. [Google Scholar]
- Antunes, M.D.C.; Neves, N.; Curado, F.; Rodrigues, S.; Franco, J.; Panagopoulos, T. The effect of calcium applications on kiwifruit quality preservation during storage. ISHS Acta Hortic. 2005, 753, 727–732. [Google Scholar] [CrossRef]
- Roy, S.; Conway, W.S.; Watada, A.E.; Sams, C.E.; Pooley, C.D.; Werning, W.P. Distribution of the anionic sites in the cell wall of apple fruit after calcium treatment. Quantification and visualization by a cationic colloidal gold probe. Protoplasma 1994, 178, 156–167. [Google Scholar] [CrossRef]
- Kim, J.O.; Shin, J.H.; Gumilang, A.; Chung, K.; Choi, K.Y.; Kim, S.K. Effectiveness of different classes of fungicides on Botrytis cinerea causing gray mold on fruit and vegetables. Plant Pathol. 2016, 32, 570–574. [Google Scholar] [CrossRef]
- Lolas-Caneo, M.; Diaz, G.A.; Mendez, R.; Caceres, M.; Neubauer, L. Evaluation of the efficacy of fungicide fludioxonil in the post-harvest control of bull’s eye rot (Neofabraea alba) in Chile. Acta Hortic. 2016, 1144, 461–464. [Google Scholar] [CrossRef]
- Mari, M.; Bertolini, P.; Pratella, G.C. Non-conventional methods for the control of post-harvest pear diseases. J. Appl. Microbiol. 2003, 94, 761–766. [Google Scholar] [CrossRef] [Green Version]
- Isshiki, K.; Tokuora, K.; Mori, R.; Chiba, S. Preliminary examination of allyl isothiocyanate vapor for food preservation. Biosci. Biotechnol. Biochem. 1992, 56, 1476–1477. [Google Scholar] [CrossRef]
- Le Cam, B.; Massiot, P.; Campion, C.; Rouxel, F. Susceptibility of carrot cultivars to Mycocentrospora acerina and the structure of cell wall polysaccharides. Physiol. Mol. Plant Pathol. 1994, 45, 139–215. [Google Scholar] [CrossRef]
- McMillan, G.P.; Hedley, D.; Fyffe, L.; Pérombelon, M.C.M. Potato resistance to soft-rot erwinias is related to cell wall pectin esterification. Physiol. Mol. Plant Pathol. 1993, 42, 279–289. [Google Scholar] [CrossRef]
- Marty, P.; Jouan, B.; Bertheau, Y.; Vian, B.; Goldberg, R. Charge density in stem cell walls of solanum tuberosum genotypes and susceptibility to blackleg. Phytochemistry 1997, 44, 1435–1441. [Google Scholar] [CrossRef]
- Wydra, K.; Berl, H. Structural changes of homogalacturonan, rhamnogalacturonan I and arabinogalactan protein in xylem cell walls of tomato genotypes in reaction to Ralstonia solanacearum. Physiol. Mol. Plant Pathol. 2006, 68, 41–50. [Google Scholar] [CrossRef]
- Di Francesco, A.; Mari, M. Use of biocontrol agents in combination with physical and chemical treatments: Efficacy assessment. Stewart Postharvest Rev. 2014, 1, 1–4. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Pathogen Isolates (mm) | |||
|---|---|---|---|---|
| CadA | CadB | CadC | ||
| VOCs | Control | 30 ± 0.70 c | 30 ± 2.0 c | 29 ± 1.25 d |
| L1 | 20 ± 1.92 b | 23 ± 2.82 b | 25 ± 2.08 c | |
| L8 | 21 ± 2.07 b | 24 ± 1.25 b | 21 ± 1.73 b | |
| Th1 | 9 ± 1.70 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| No-VOCs | Control | 30 ± 0.70 d | 30 ± 2.0 d | 29 ± 1.25 c |
| L1 | 16 ± 1.85 b | 16 ± 0.66 b | 15 ± 1.20 b | |
| L8 | 21 ± 2.33 c | 20 ± 1.45 c | 17 ± 1.52 b | |
| Th1 | 8 ± 0.33 a | 10 ± 0.00 a | 8 ± 0.88 a | |
| EC50 Values (g/L) | |||
|---|---|---|---|
| Isolate | Fludioxonil | CaCl2 | |
| CFU | Mycelium | Mycelium | |
| CadA | 0.13 a | 0.18 a | 16.8 a |
| CadB | 0.65 c | 1.4 b | 28.0 c |
| CadC | 0.32 b | 1.2 b | 22.7 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Francesco, A.; Di Foggia, M.; Vittoria, A.; Baraldi, E. Post-Harvest Non-Conventional and Traditional Methods to Control Cadophora luteo-olivacea: Skin Pitting Agent of Actinidia chinensis var. deliciosa (A. Chev.). Horticulturae 2021, 7, 169. https://doi.org/10.3390/horticulturae7070169
Di Francesco A, Di Foggia M, Vittoria A, Baraldi E. Post-Harvest Non-Conventional and Traditional Methods to Control Cadophora luteo-olivacea: Skin Pitting Agent of Actinidia chinensis var. deliciosa (A. Chev.). Horticulturae. 2021; 7(7):169. https://doi.org/10.3390/horticulturae7070169
Chicago/Turabian StyleDi Francesco, Alessandra, Michele Di Foggia, Alessio Vittoria, and Elena Baraldi. 2021. "Post-Harvest Non-Conventional and Traditional Methods to Control Cadophora luteo-olivacea: Skin Pitting Agent of Actinidia chinensis var. deliciosa (A. Chev.)" Horticulturae 7, no. 7: 169. https://doi.org/10.3390/horticulturae7070169
APA StyleDi Francesco, A., Di Foggia, M., Vittoria, A., & Baraldi, E. (2021). Post-Harvest Non-Conventional and Traditional Methods to Control Cadophora luteo-olivacea: Skin Pitting Agent of Actinidia chinensis var. deliciosa (A. Chev.). Horticulturae, 7(7), 169. https://doi.org/10.3390/horticulturae7070169