
Water Use Efficiency and Nutritional Status of a New Grapevine Rootstock Selection


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Characterization of the Experimental Sites
2.3. Leaf Temperature and Gas Exchange
2.4. Mineral Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Plasticity of Traits in Response to the Environment
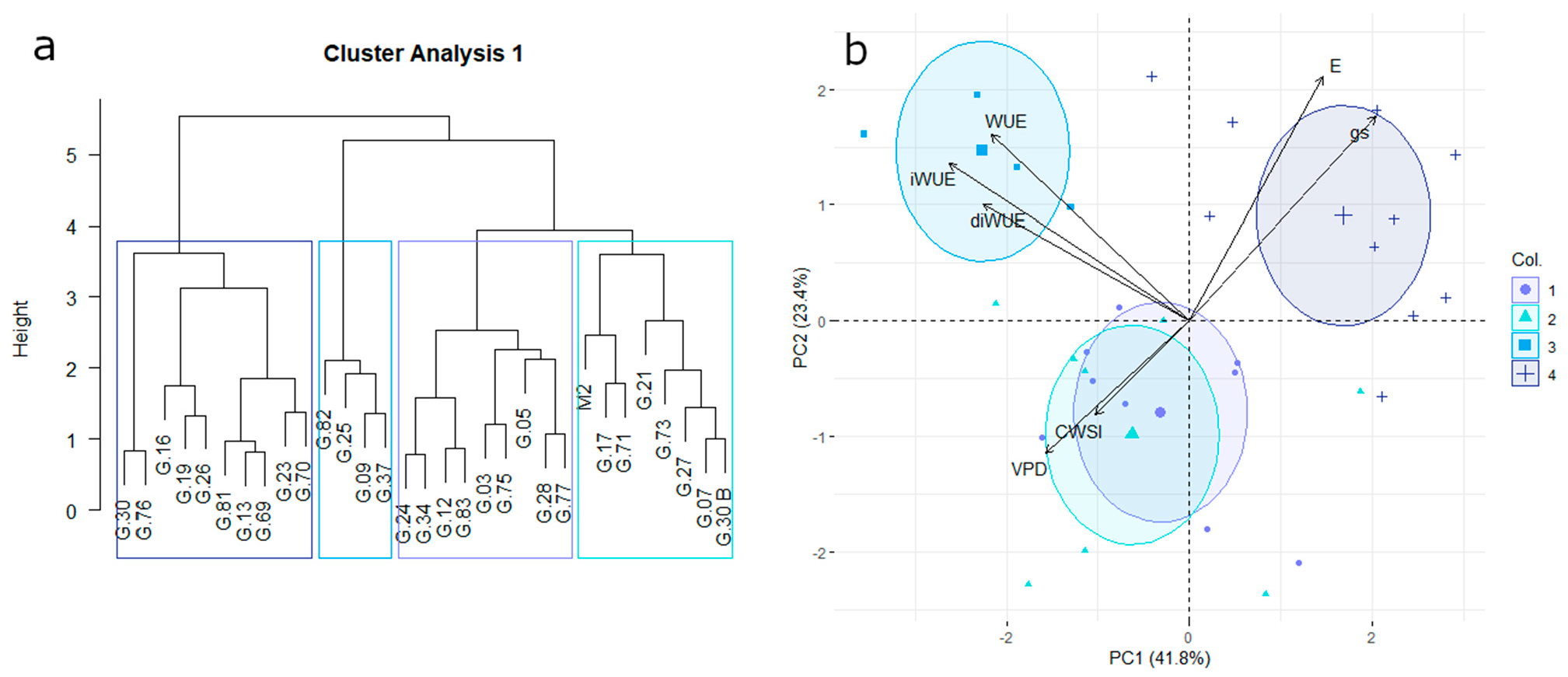
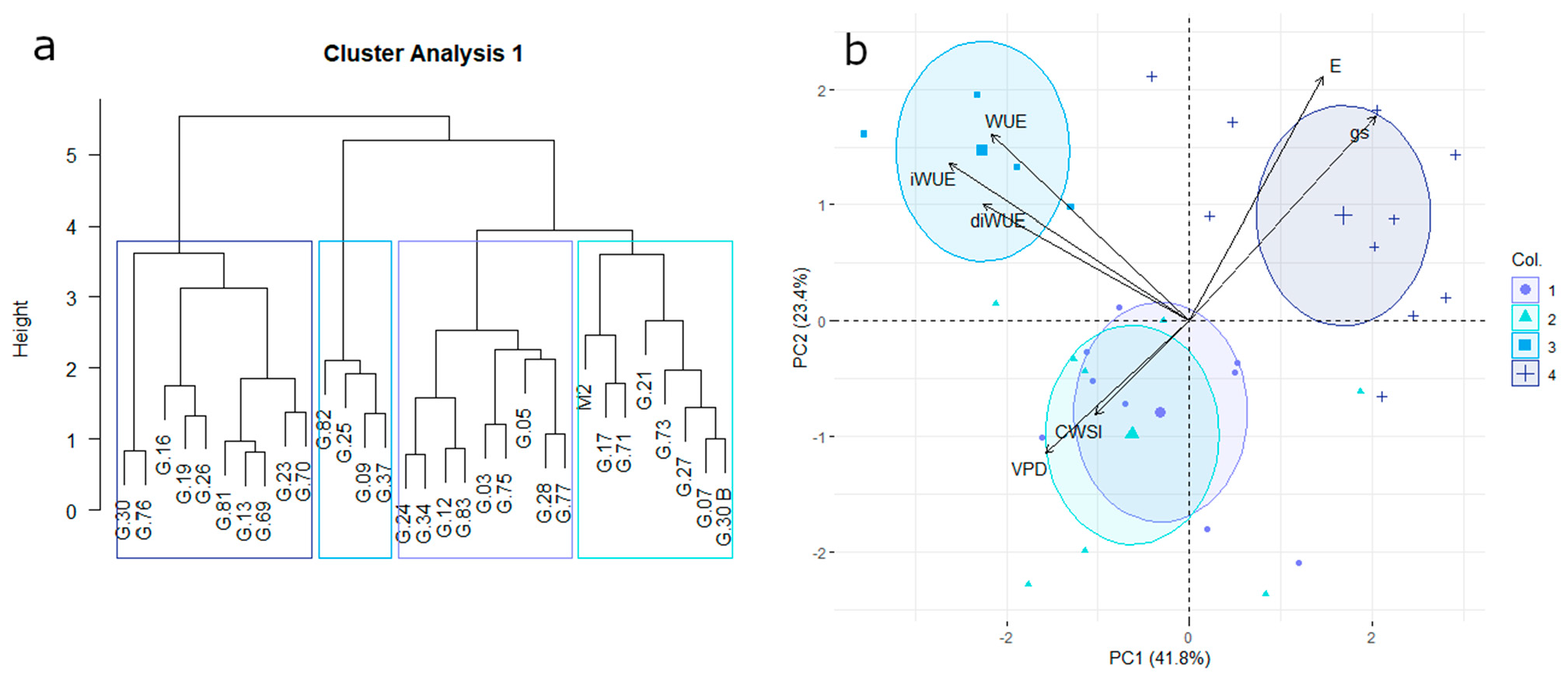
3.2. Water-Use Efficiency and Transpiration Control
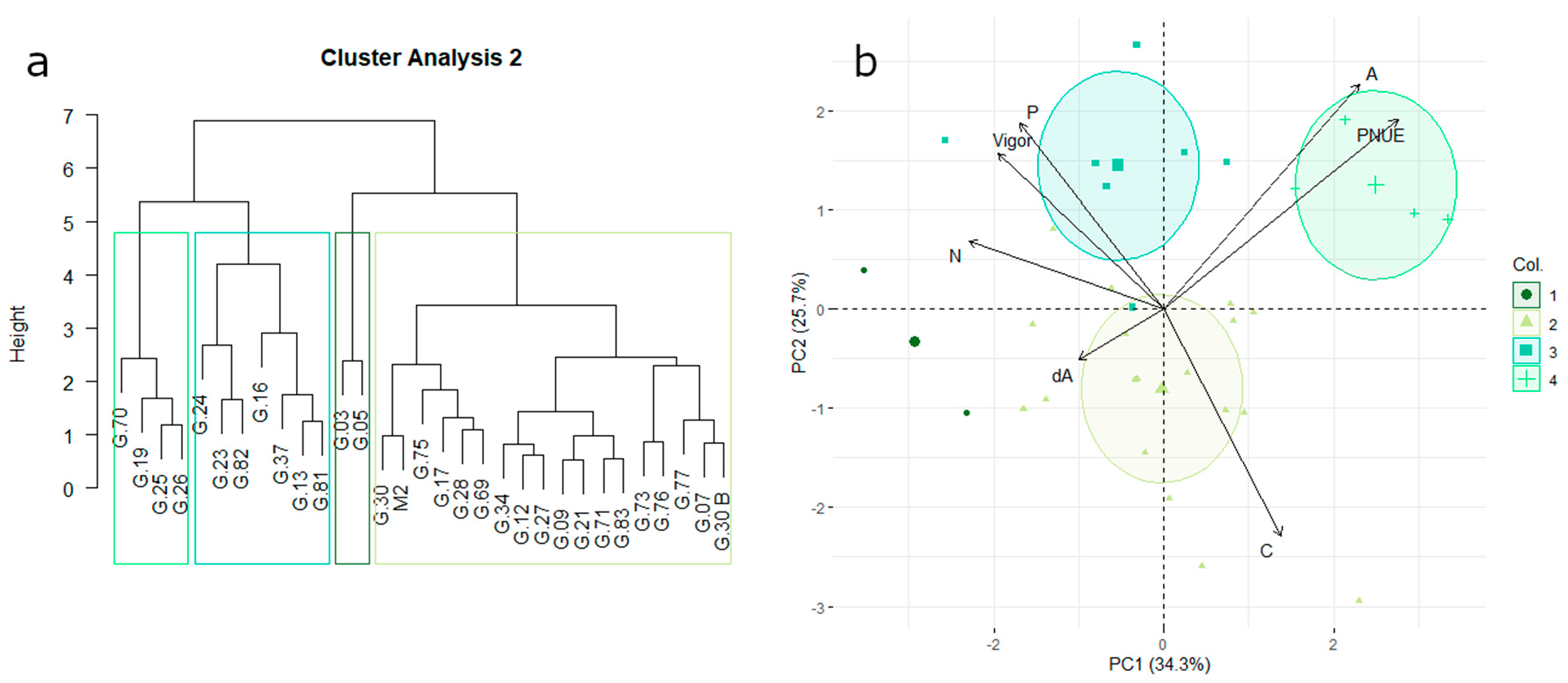
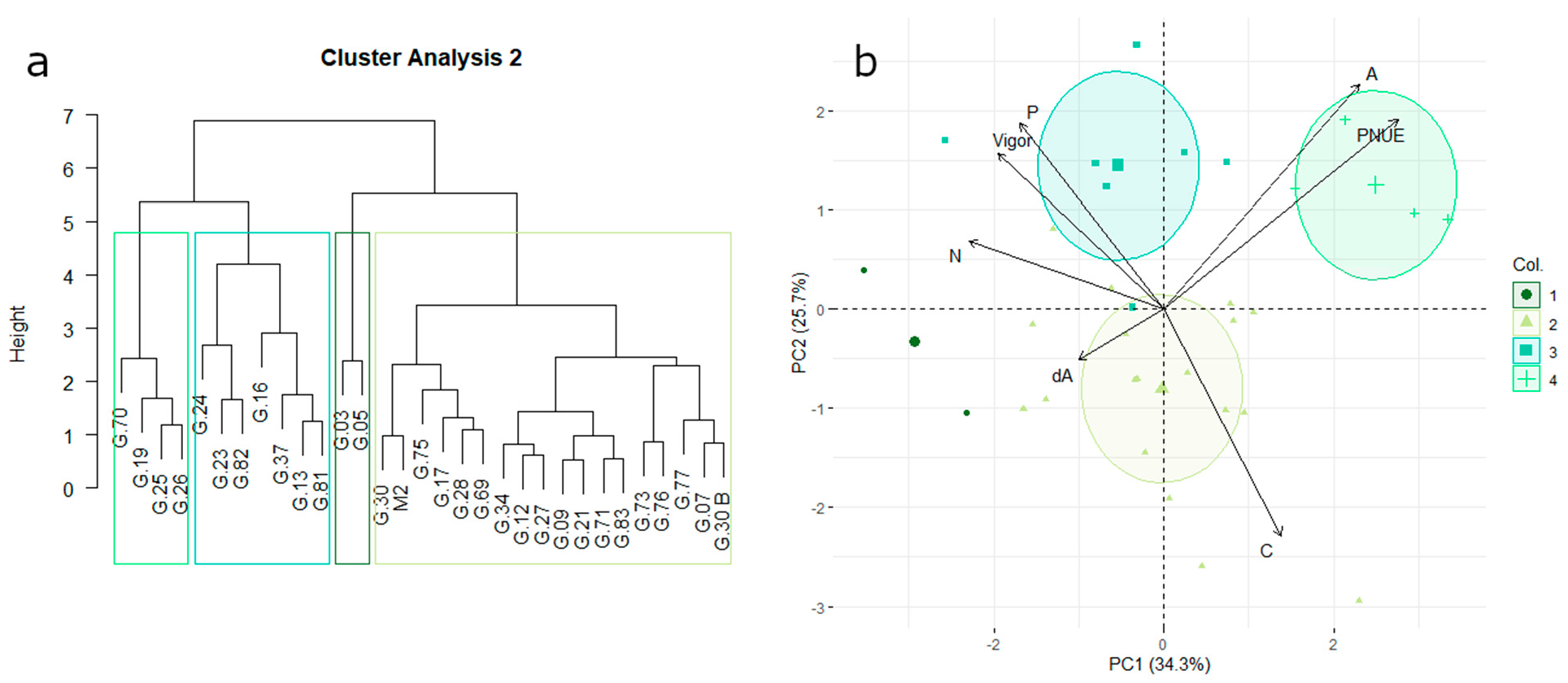
3.3. Vegetative Growth, Carbon Assimilation, and Nitrogen-Use Efficiency
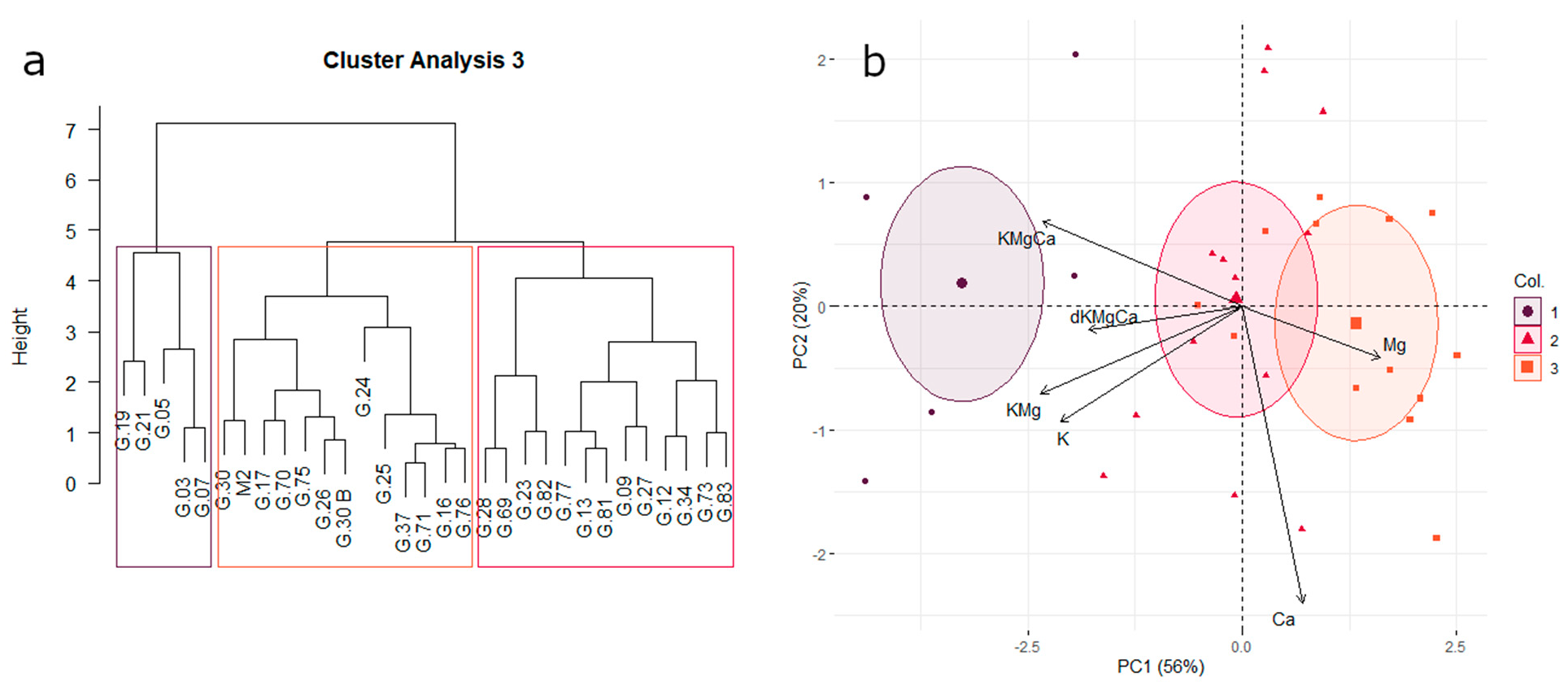
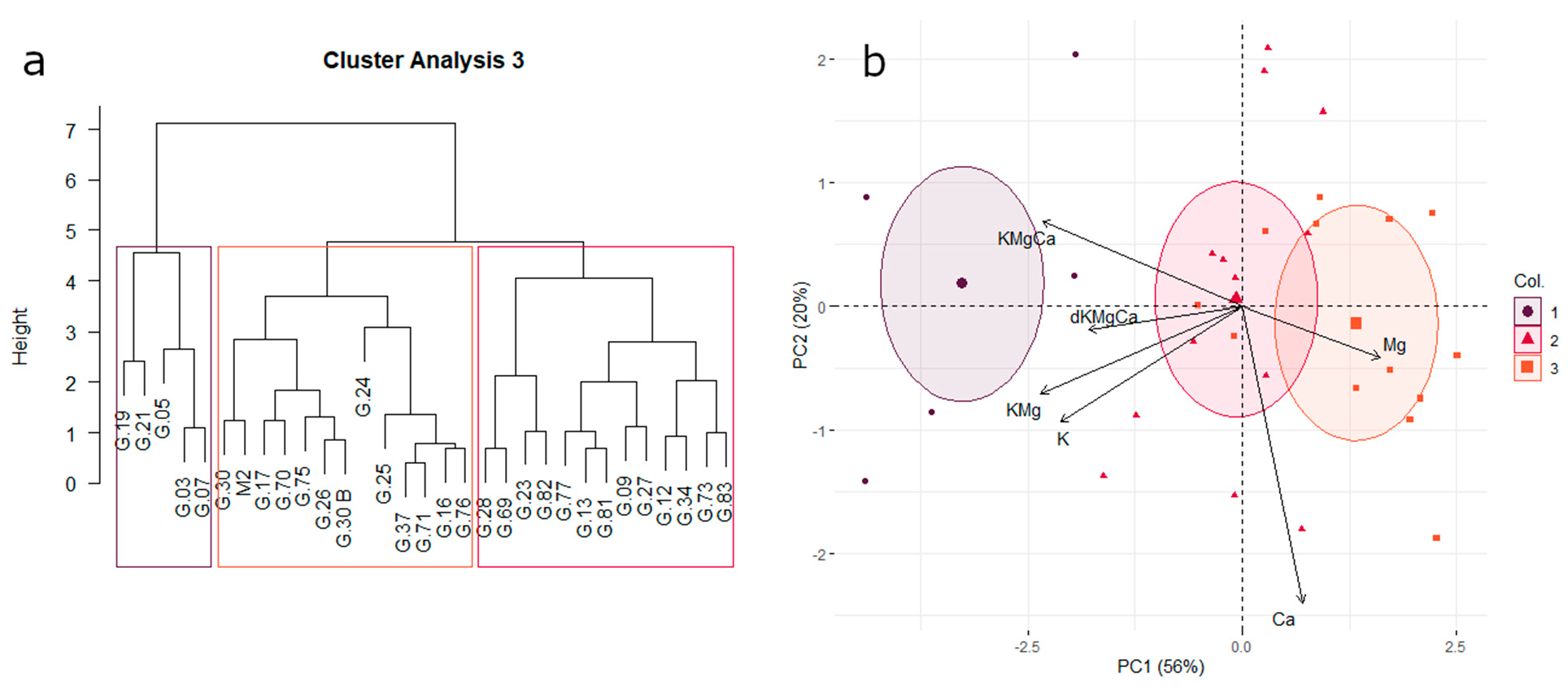
3.4. Affinity for Potassium or Magnesium Uptake
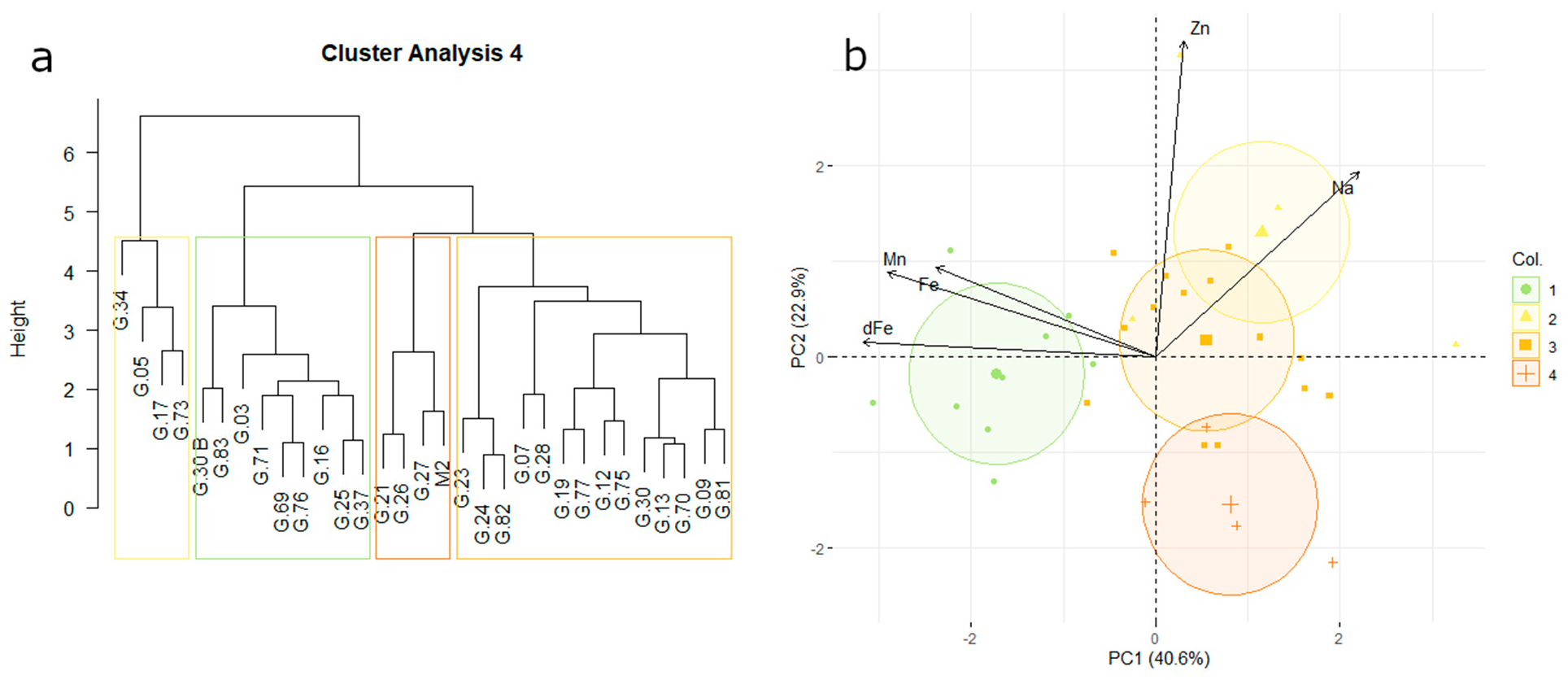
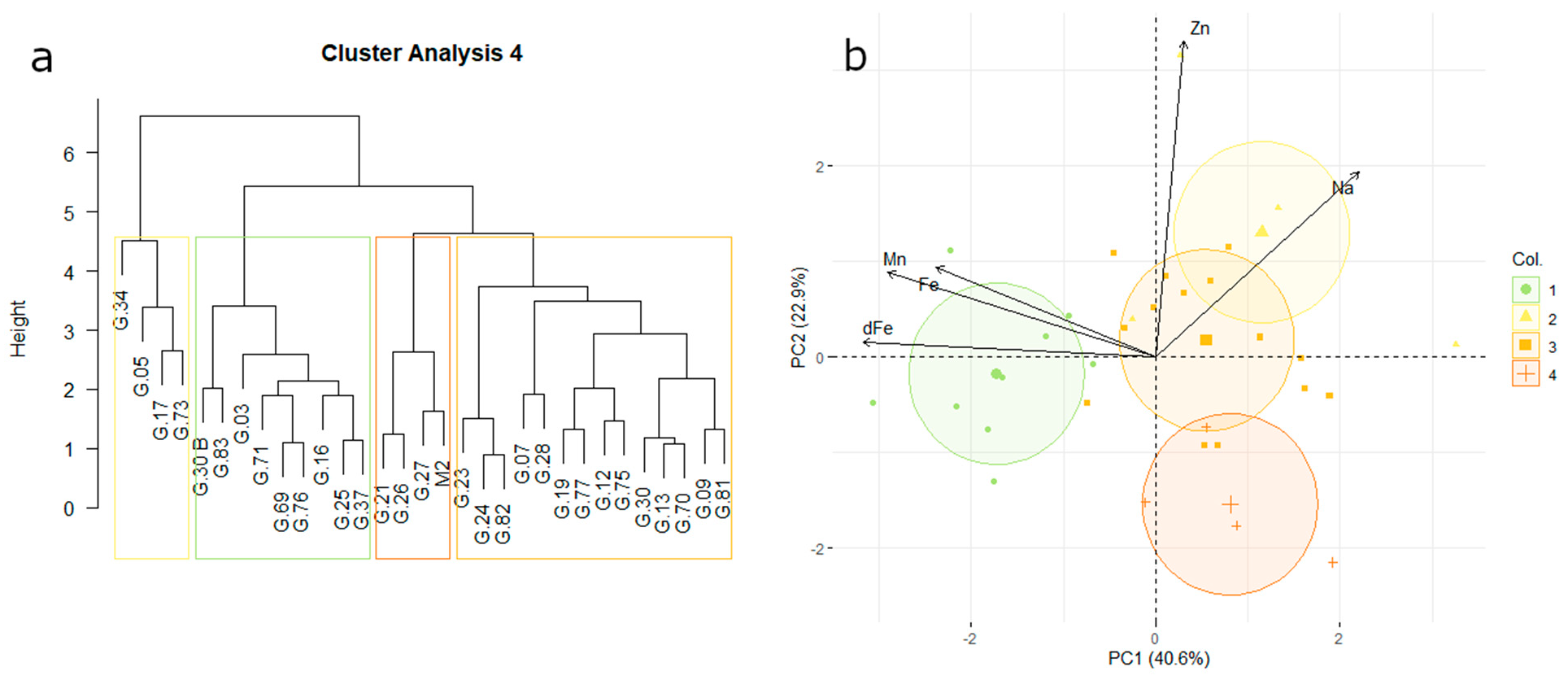
3.5. Uptake Affinity for Micronutrients
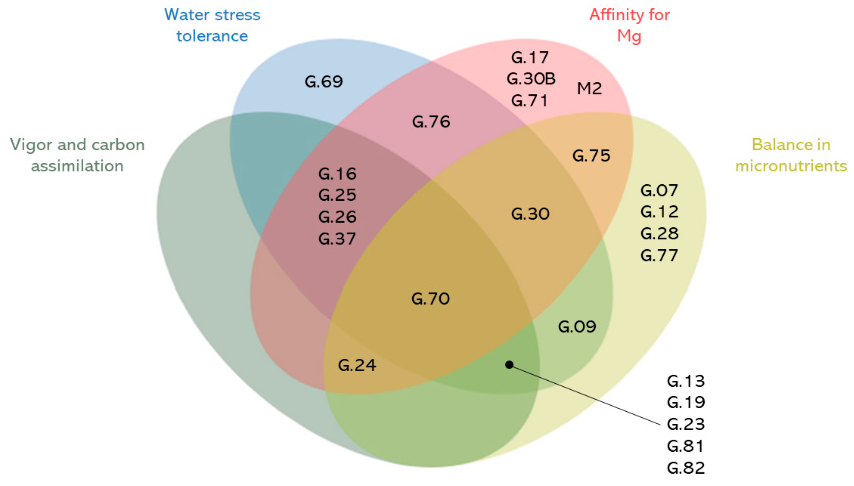
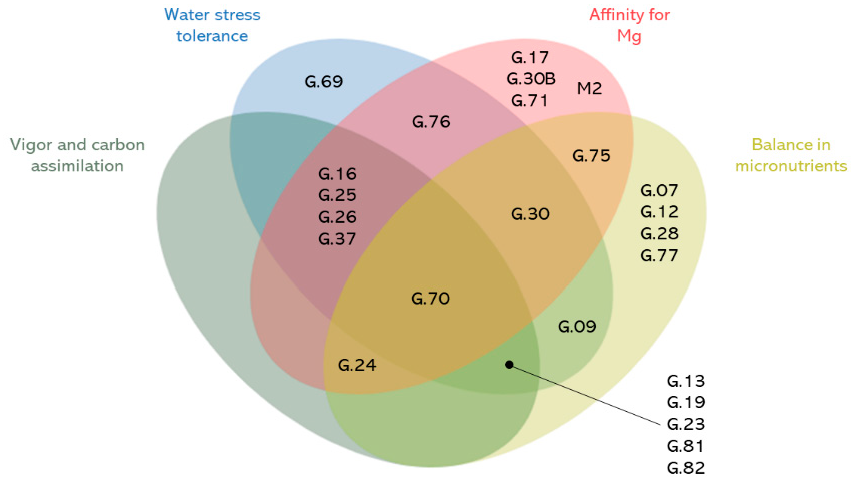
3.6. Promising Rootstocks for Adaptation to Abiotic Stresses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.M.; Akram, M.T.; Qadri, R.W.K.; Al-Yahyai, R. Role of grapevine rootstocks in mitigating environmental stresses: A review. J. Agric. Mar. Sci. 2020, 25, 1–12. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Cousins, P.; Jordan, L.M.; Myles, S.; Striegler, R.K.; Verdegaal, P.; Chitwood, D.H. Grapevine rootstocks affect growth-related scion phenotypes. Plant Direct 2021, 5, e00324. [Google Scholar] [CrossRef]
- Zombardo, A.; Crosatti, C.; Bagnaresi, P.; Bassolino, L.; Reshef, N.; Puccioni, S.; Faccioli, P.; Tafuri, A.; Delledonne, M.; Fait, A.; et al. Transcriptomic and biochemical investigations support the role of rootstock-scion interaction in grapevine berry quality. BMC Genom. 2020, 21, 468. [Google Scholar] [CrossRef]
- Keller, M. From the ASEV Soil Environment and Vine Mineral Nutrition Symposium: Deficit irrigation and vine mineral nutrition. Am. J. Enol. Vitic. 2005, 56, 267–283. [Google Scholar]
- Quénol, H.; Marie, G.; Barbeau, G.; Van Leeuwen, C.; Foss, C.; Irimia, L.; Rochard, J.; Boulanger, J.; Tissot, C.; Quénol, H.; et al. Adaptation of viticulture to climate change: High resolution observations of adaptation scenario for viticulture: The adviclim european project. Bull. l’OIV 2014, 87, 395–406. [Google Scholar]
- Galbignani, M.; Merli, M.; Magnanini, E.; Bernizzoni, F.; Talaverano, I.; Gatti, M.; Tombesi, S.; Palliotti, A.; Poni, S. Gas exchange and water—Use efficiency of cv. Sangiovese grafted to rootstocks of varying water—Deficit tolerance. Irrig. Sci. 2016, 34, 105–116. [Google Scholar] [CrossRef]
- Frioni, T.; Biagioni, A.; Squeri, C.; Tombesi, S.; Gatti, M.; Poni, S. Grafting cv. grechetto gentile vines to new m4 rootstock improves leaf gas exchange and water status as compared to commercial 1103p rootstock. Agronomy 2020, 10, 708. [Google Scholar] [CrossRef]
- Mirás-Avalos, J.M.; Araujo, E.S. Optimization of Vineyard Water Management: Challenges, Strategies, and Perspectives. Water 2021, 13, 746. [Google Scholar] [CrossRef]
- Zhang, L.; Marguerit, E.; Rossdeutsch, L.; Ollat, N.; Gambetta, G.A. The influence of grapevine rootstocks on scion growth and drought resistance. Theor. Exp. Plant Physiol. 2016, 28, 143–157. [Google Scholar] [CrossRef]
- Leão, P.C.d.S.; Chaves, A.R.d.M. Agronomic responses of grapevine ‘Chenin Blanc’ as a function of training systems and rootstocks. Sci. Agric. 2020, 78, e20180413. [Google Scholar] [CrossRef]
- Rossdeutsch, L.; Schreiner, R.P.; Skinkis, P.A.; Deluc, L. Nitrate Uptake and Transport Properties of Two Grapevine Rootstocks With Varying Vigor. Front. Plant Sci. 2021, 11, 608813. [Google Scholar] [CrossRef]
- Verdugo-Vásquez, N.; Gutiérrez-Gamboa, G.; Díaz-Gálvez, I.; Ibacache, A.; Zurita-Silva, A. Modifications induced by rootstocks on yield, vigor and nutritional status on vitis vinifera cv syrah under hyper-arid conditions in Northern Chile. Agronomy 2021, 11, 979. [Google Scholar] [CrossRef]
- Ibacache, A.; Verdugo-Vásquez, N.; Zurita-Silva, A. Rootstock: Scion combinations and nutrient uptake in grapevines. Fruit Crops Diagn. Manag. Nutr. Constraints 2019, 297–316. [Google Scholar] [CrossRef]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Gautier, A.T.; Cookson, S.J.; Lagalle, L.; Ollat, N.; Marguerit, E. Influence of the three main genetic backgrounds of grapevine rootstocks on petiolar nutrient concentrations of the scion, with a focus on phosphorus. Oeno One 2020, 54, 1–13. [Google Scholar] [CrossRef]
- Casanova-Gascón, J.; Martín-Ramos, P.; Martí-Dalmau, C.; Badía-Villas, D. Nutrients Assimilation and Chlorophyll Contents for Different Grapevine Varieties in Calcareous Soils in the Somontano DO (Spain). Beverages 2018, 4, 90. [Google Scholar] [CrossRef] [Green Version]
- Toumi, M.; Nedjimi, B.; Halitim, A.; Garcia, M. Effects of K-Mg ratio on growth and cation nutrition of Vitis vinifera L. cv. “Dattier de Beiruth” grafted on SO4 rootstock. J. Plant Nutr. 2016, 39, 904–911. [Google Scholar] [CrossRef]
- Brancadoro, L.; Reina, L.V.A.; Scienza, A. Potassium content of grapevine during the vegetative period: The role of the rootstock. J. Plant Nutr. 1994, 17, 2165–2175. [Google Scholar] [CrossRef]
- Kodur, S. Effects of juice pH and potassium on juice and wine quality, and regulation of potassium in grapevines through rootstocks (Vitis): A short review. Vitis J. Grapevine Res. 2011, 50, 1–6. [Google Scholar]
- Marastoni, L.; Lucini, L.; Miras-Moreno, B.; Trevisan, M.; Sega, D.; Zamboni, A.; Varanini, Z. Changes in physiological activities and root exudation profile of two grapevine rootstocks reveal common and specific strategies for Fe acquisition. Sci. Rep. 2020, 10, 18839. [Google Scholar] [CrossRef] [PubMed]
- Dal Santo, S.; Zenoni, S.; Sandri, M.; De Lorenzis, G.; Magris, G.; De Paoli, E.; Di Gaspero, G.; Del Fabbro, C.; Morgante, M.; Brancadoro, L.; et al. Grapevine field experiments reveal the contribution of genotype, the influence of environment and the effect of their interaction (G×E) on the berry transcriptome. Plant J. 2018, 93, 1143–1159. [Google Scholar] [CrossRef]
- Pinto, D.L.P.; Brancadoro, L.; Dal Santo, S.; De Lorenzis, G.; Pezzotti, M.; Meyers, B.C.; Pè, M.E.; Mica, E. The influence of genotype and environment on small RNA profiles in grapevine berry. Front. Plant Sci. 2016, 7, 1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, D.; Grossi, D.; Tincani, D.T.G.; Simone Di Lorenzo, G.; Brancadoro, L.; Rustioni, L. Multi-parameter characterization of water stress tolerance in Vitis hybrids for new rootstock selection. Plant Physiol. Biochem. 2018, 132, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, D.; Grossi, D.; Simone Di Lorenzo, G.; Zi Ying, Y.; Rustioni, L.; Brancadoro, L. Phenotyping of the “G series” Vitis hybrids: First screening of the mineral composition. Sci. Hortic. 2020, 264, 109155. [Google Scholar] [CrossRef]
- Migliaro, D.; De Lorenzis, G.; Simone Di Lorenzo, G.; De Nardi, B.; Gardiman, M.; Failla, O.; Brancadoro, L.; Crespan, M. Grapevine non-vinifera genetic diversity assessed by SSR markers as a starting-point for new rootstock breeding programs. Am. J. Enol. Vitic. 2019, in press. [Google Scholar] [CrossRef]
- Grant, O.M.; Chaves, M.M.; Jones, H.G. Optimizing thermal imaging as a technique for detecting stomatal closure induced by drought stress under greenhouse conditions. Physiol. Plant. 2006, 127, 507–518. [Google Scholar] [CrossRef]
- Jones, H.G.; Stoll, M.; Santos, T.; De Sousa, C.; Chaves, M.M.; Grant, O.M. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 53, 2249–2260. [Google Scholar] [CrossRef] [PubMed]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J., Jr.; Reginato, R.J.; Hatfield, J.L. Normalizing the stress-degree-day parameter for environmental variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Jones, H.G. Use of thermography for quantitative studies of spatial and temporal variation of stomatal conductance over leaf surfaces. Plant Cell Environ. 1999, 22, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. SOFTWARE Open Access jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef] [Green Version]
- Cifre, J.; Bota, J.; Escalona, J.M.; Medrano, H.; Flexas, J. Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.) An open gate to improve water-use efficiency? Agric. Ecosyst. Environ. 2005, 106, 159–170. [Google Scholar] [CrossRef]
- Bellvert, J.; Zarco-Tejada, P.J.; Girona, J.; Fereres, E. Mapping crop water stress index in a ‘Pinot-noir’ vineyard: Comparing ground measurements with thermal remote sensing imagery from an unmanned aerial vehicle. Precis. Agric. 2014, 15, 361–376. [Google Scholar] [CrossRef]
- Rustioni, L.; Bianchi, D. Drought increases chlorophyll content in stems of Vitis interspecific hybrids. Theor. Exp. Plant Physiol. 2021, 33, 69–78. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Stage | u.m. | Arcagna | Riccagioia | E | G | G × E |
|---|---|---|---|---|---|---|---|
| gs | r | mmol H2O m−2 s−1 | 201.71 ± 10.47 | 46.18 ± 3.49 | *** | *** | *** |
| E | r | mmol H2O m−2 s−1 | 3.22 ± 0.12 | 1.2 ± 0.07 | *** | *** | *** |
| WUE | r | mmol CO2 mol−1 H2O | 3.48 ± 0.13 | 3.17 ± 0.26 | n.s. | *** | *** |
| iWUE | r | µmol CO2 mol−1 H2O | 61.68 ± 3.11 | 93.24 ± 8.47 | *** | *** | *** |
| VPD | r | kPa | 1.94 ± 0.06 | 2.95 ± 0.09 | *** | *** | *** |
| CWSI | f | - | 0.39 ± 0.01 | 0.52 ± 0.01 | *** | ** | *** |
| CWSI | v | - | 0.20 ± 0.01 | 0.71 ± 0.01 | *** | *** | *** |
| CWSI | r | - | 0.23 ± 0.01 | 0.63 ± 0.01 | *** | *** | *** |
| Vigor | d | kg | 1.20 ± 0.06 | 0.48 ± 0.03 | *** | *** | *** |
| A | r | µmol CO2 m−2 s−1 | 10.43 ± 0.32 | 3.84 ± 0.34 | *** | *** | *** |
| C | f | % | 44.72 ± 0.13 | 43.48 ± 0.13 | *** | n.s. | * |
| C | r | % | 43.68 ± 0.38 | 45.08 ± 0.17 | ** | n.s. | n.s. |
| N | f | % | 2.99 ± 0.06 | 2.63 ± 0.04 | *** | *** | *** |
| N | r | % | 1.62 ± 0.03 | 1.86 ± 0.02 | *** | *** | *** |
| PNUE | r | µmol CO2 g−1 N s−1 | 4.64 ± 0.14 | 1.98 ± 0.16 | *** | *** | *** |
| P | f | % | 0.32 ± 0.01 | 0.37 ± 0.02 | ** | *** | ** |
| P | r | % | 0.22 ± 0.01 | 0.22 ± 0.01 | n.s. | *** | n.s. |
| K | f | % | 0.98 ± 0.02 | 0.76 ± 0.03 | *** | *** | ** |
| K | r | % | 0.95 ± 0.04 | 0.44 ± 0.01 | *** | *** | *** |
| Mg | f | % | 0.18 ± 0.01 | 0.27 ± 0.01 | *** | *** | *** |
| Mg | r | % | 0.31 ± 0.01 | 0.52 ± 0.01 | *** | *** | *** |
| Ca | f | % | 1.54 ± 0.05 | 1.38 ± 0.05 | ** | *** | ** |
| Ca | r | % | 2.57 ± 0.07 | 2.51 ± 0.05 | n.s. | *** | *** |
| KMg | f | - | 6.03 ± 0.34 | 2.99 ± 0.14 | *** | *** | *** |
| KMg | r | - | 3.75 ± 0.33 | 0.93 ± 0.05 | *** | *** | *** |
| KMgCa | f | - | 0.62 ± 0.03 | 0.51 ± 0.03 | *** | *** | *** |
| KMgCa | r | - | 0.36 ± 0.02 | 0.15 ± 0.01 | *** | ** | ** |
| Na | f | ppm | 37.68 ± 2.86 | 46.96 ± 3.92 | n.s. | n.s. | ** |
| Na | r | ppm | 59.36 ± 7.33 | 34.56 ± 2.27 | ** | n.s. | n.s. |
| Fe | f | ppm | 235.51 ± 7.44 | 120.5 ± 5.94 | *** | *** | *** |
| Fe | r | ppm | 313.79 ± 8.95 | 276.03 ± 6.53 | *** | *** | ** |
| Mn | f | ppm | 179.16 ± 7.34 | 144.02 ± 7.43 | *** | *** | *** |
| Mn | r | ppm | 156.26 ± 6.94 | 62.21 ± 2.61 | *** | *** | *** |
| Zn | f | ppm | 21.75 ± 1.02 | 40.06 ± 3.22 | *** | *** | *** |
| Zn | r | ppm | 24.91 ± 1.49 | 25.15 ± 1.46 | n.s. | n.s. | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianchi, D.; Brancadoro, L. Water Use Efficiency and Nutritional Status of a New Grapevine Rootstock Selection. Horticulturae 2021, 7, 503. https://doi.org/10.3390/horticulturae7110503
Bianchi D, Brancadoro L. Water Use Efficiency and Nutritional Status of a New Grapevine Rootstock Selection. Horticulturae. 2021; 7(11):503. https://doi.org/10.3390/horticulturae7110503
Chicago/Turabian StyleBianchi, Davide, and Lucio Brancadoro. 2021. "Water Use Efficiency and Nutritional Status of a New Grapevine Rootstock Selection" Horticulturae 7, no. 11: 503. https://doi.org/10.3390/horticulturae7110503
APA StyleBianchi, D., & Brancadoro, L. (2021). Water Use Efficiency and Nutritional Status of a New Grapevine Rootstock Selection. Horticulturae, 7(11), 503. https://doi.org/10.3390/horticulturae7110503