Mango Endophyte and Epiphyte Microbiome Composition during Fruit Development and Post-Harvest Stages


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Trial Design and Sample Collection
2.2. Post-Harvest Fruit Quality and Decay Parameters of Mango Fruit
2.3. Mango Microbiome Sample Preparation
2.3.1. Epiphytic (Fructoplane) Microbiome
2.3.2. Endophytic (Stem-End Pulp) Microbiome
2.4. DNA Extraction, Amplification and Sequencing
2.5. Illumina Sequence Processing and Analysis
2.6. Statistical Analysis
3. Results
3.1. Physiological Maturity Parameters and Disease Incidence
3.2. Diversity of Fungal and Bacterial Communities during Different Stages of Mango Production
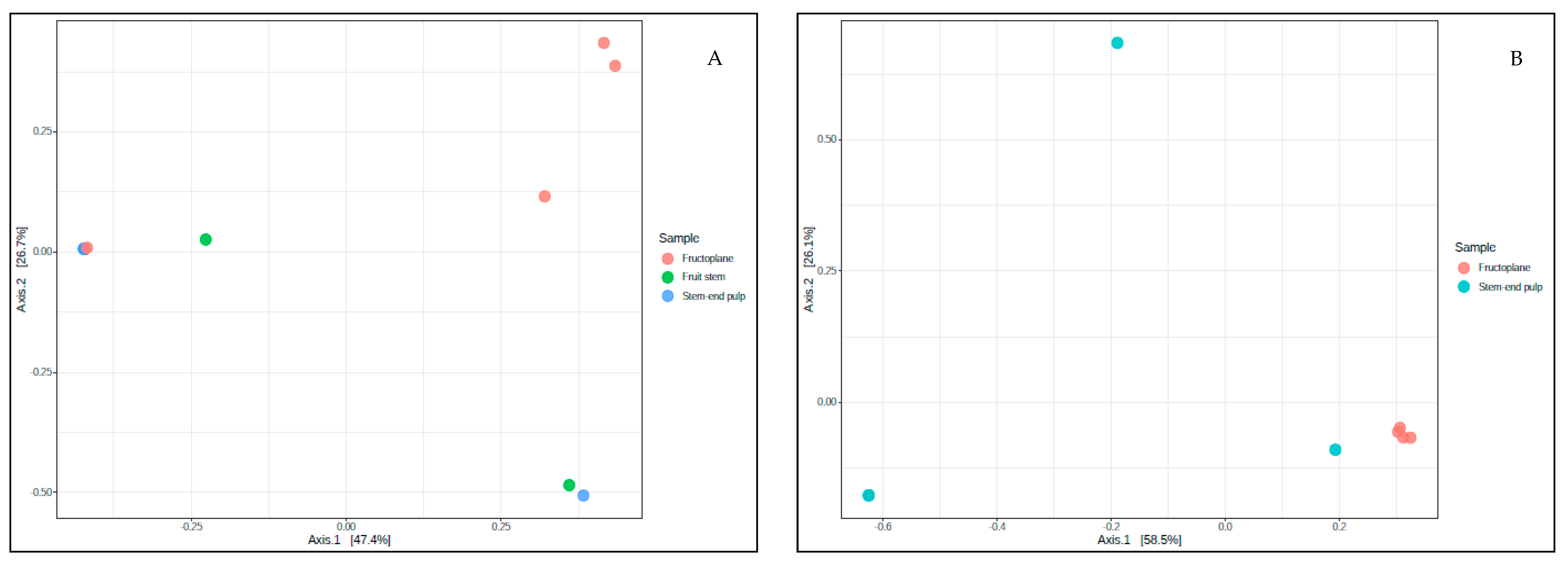
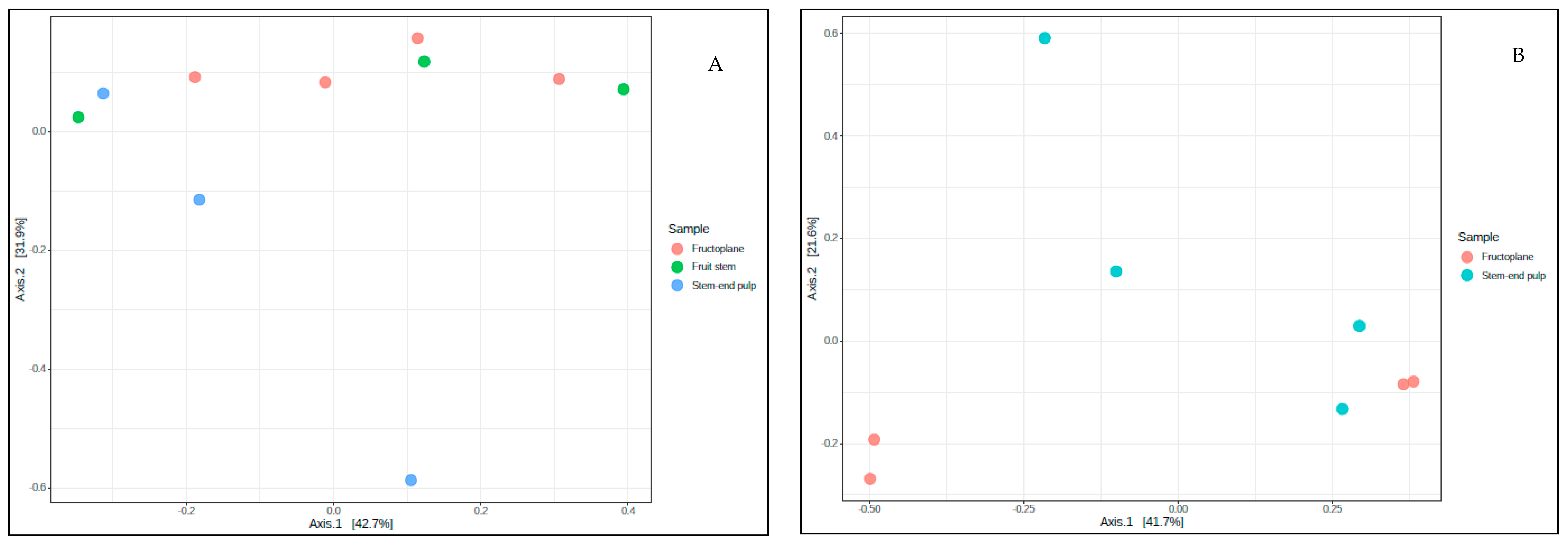
Beta Diversity
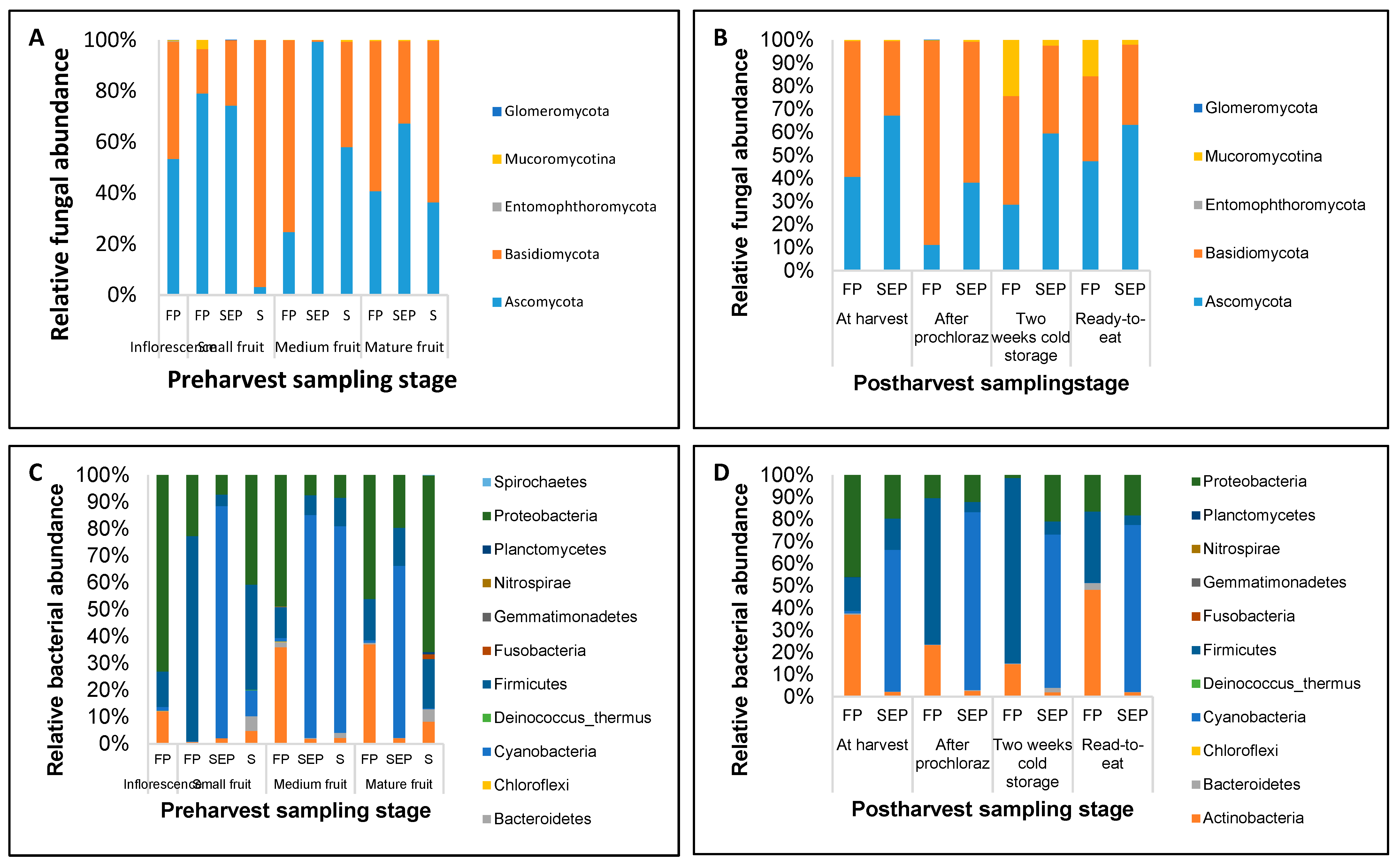
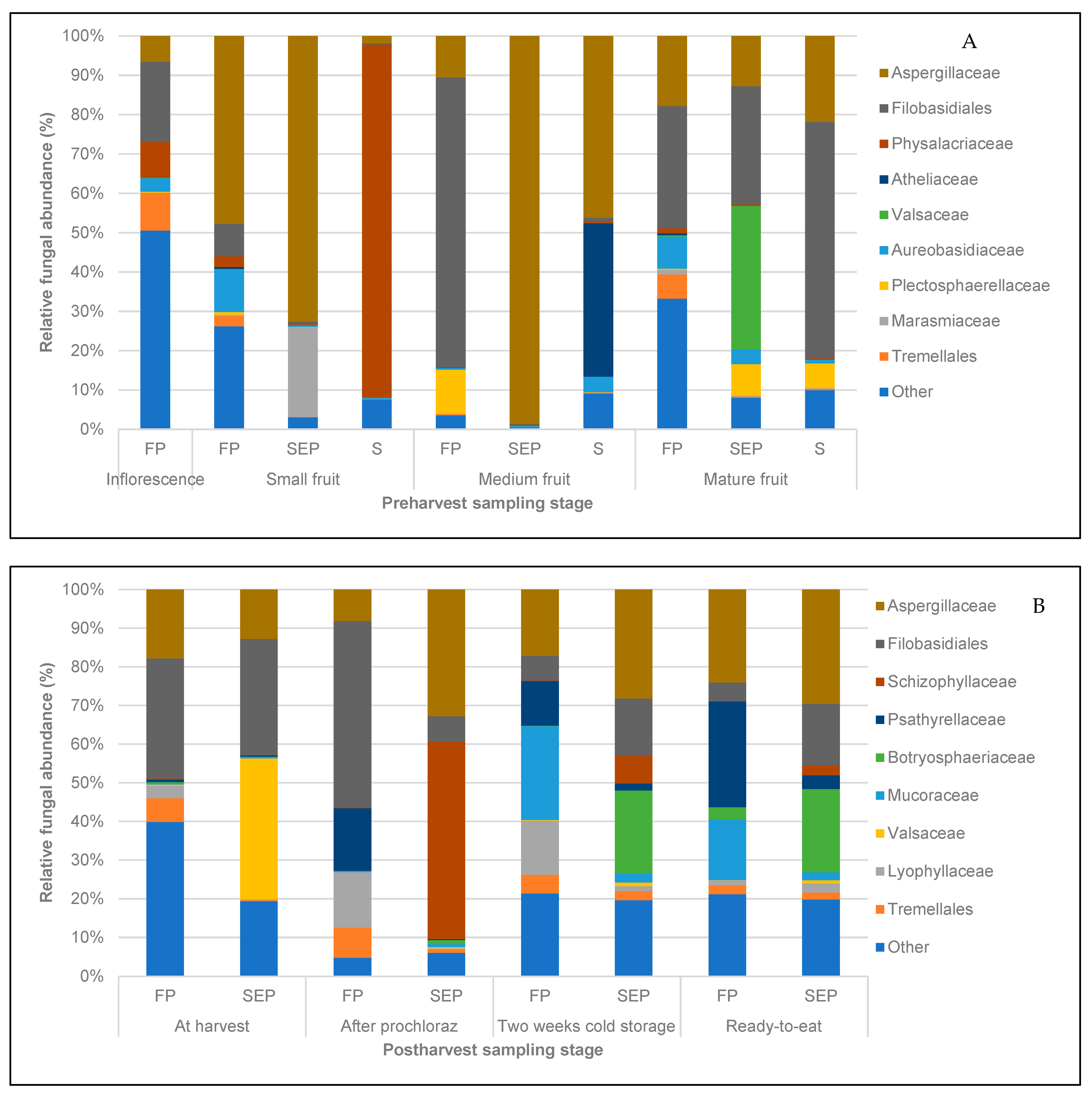
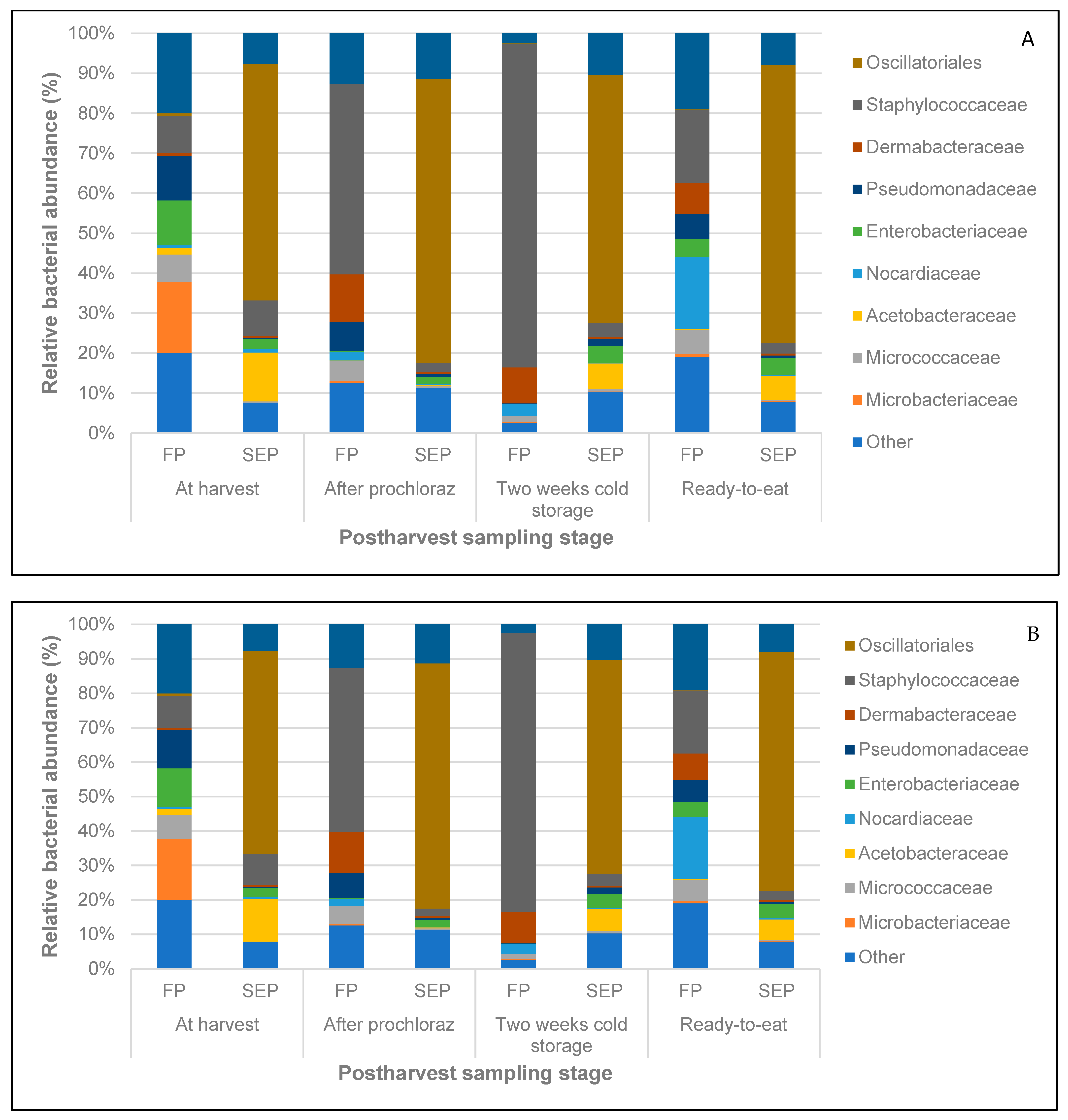
3.3. Structure of Fungal and Bacterial Communities during Different Stages of Mango Production
3.4. Identification of Pathogenic Fungal Taxa at the Pre- and Post-harvest Stages of Mango
3.5. Beneficial Bacterial Taxa Present at the Pre- and Post-Harvest Stages of Mango
4. Discussion
4.1. Microbial Diversity and Composition at the Pre- and Post-Harvest Stages of Mango
4.2. Pathogenic Fungal Taxa Composition at the Pre- and Post-Harvest Stages of Mango
4.3. Beneficial Bacterial Taxa Composition at the Pre- and Post-Harvest Stages of Mangoes
4.4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pal, R.K. Ripening and rheological properties of mango as influenced by ethereal and carbide. J. Food Sci. Technol. 1998, 35, 358–360. [Google Scholar]
- Dick, E.; Adopo, A.N.; Camara, B.; Moudioh, E. Influence of maturity stage of mango at harvest on its ripening quality. Fruits 2009, 64, 13–18. [Google Scholar] [CrossRef]
- De Jager, E.S.; Wehner, F.C.; Korsten, L. Microbial ecology of mando phylloplane. Microb. Ecol. 2001, 42, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, P.C.; Siyoum, N.; Chidamba, L.; Korsten, L. Characterization of fungal communities of developmental stages in table grape grown in the northern region of South Africa. J. Appl. Microbiol. 2017, 123, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- Dukare, A.S.; Paul, S.; Nambi, V.E.; Gupta, R.K.; Singh, R.; Sharma, K.; Vishwakarma, R.K. Explotation of microbial antagonists for the control postharvest diseases of fruits: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1498–1513. [Google Scholar] [CrossRef] [PubMed]
- Perazzolli, M.; Antonielli, L.; Storari, M.; Puopolo, G.; Pancher, M.; Giovannini, O.; Pindo, M.; Pertot, I. Resilience of the natural phyllosphere microbiota of the grapevine to chemical and biological pesticides. Appl. Environ. Microbiol. 2014, 80, 3585–3596. [Google Scholar] [CrossRef]
- Sébastien, M.; Margarita, M.; Haissam, J.M. Biocontrol in the microbiome era: Challenges and opportunities. Biol. Control 2019, 89, 98–108. [Google Scholar]
- Blakeman, J.P. Ecological succession of leaf surface microorganisms in relation to biological control. In Biological Control on the Phylloplane; Windels, C.E., Lindow, S., Eds.; APS Press: New York, NY, USA, 1985; pp. 6–30. [Google Scholar]
- Stefani, F.O.; Bell, T.H.; Marchand, C.; de la Providencia, I.E.; El Yassimi, A.; St-Arnaud, M. Culture-dependent and -independent methods capture different microbial community fractions in hydrocarbon-contaminated soils. PLoS ONE 2015, 10, e0128272. [Google Scholar] [CrossRef]
- Chalupowicz, L.; Dombrovsky, A.; Gaba, V.; Luria, N.; Reuven, M.; Beerman, A.; Lachman, O.; Dror, O.; Nissan, G.; Manulis-Sasson, S. Diagnosis of plant diseases using the nanopore sequencing platform. Plant Pathol. 2019, 68, 229–238. [Google Scholar] [CrossRef]
- Takushi, T.; Arasaki, C.; Ajitomi, A.; Ooshiro, A. Stem-end rot of mango caused by Diaporthe pseudophoenicicola and Neofusicoccum parvum. Jpn. J. Phytopathol. 2016, 82, 81. [Google Scholar]
- Galsurker, O.; Diskin, S.; Maurer, D.; Feygenberg, O.; Alkan, N. Fruit stem-end Rot. Horticulturae 2018, 4, 50. [Google Scholar] [CrossRef]
- Li, L.; Mohd, M.H.; Mohamed Nor, N.M.I.; Subramaniam, S.; Latiffah, Z. Identification of Botryosphaeriaceae associated with stem-end rot of mango (Mangifera indica L.) in Malaysia. J. Appl. Microbiol. 2020, 130, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Alemu, K. Dynamics and management of major postharvest fungal diseases of mango fruits. J. Biol. Agric. Healthc. 2014, 4, 13–21. [Google Scholar]
- Giblin, F.R.; Tan, Y.P.; Mitchell, R.; Coates, L.M.; Irwin, J.A.G.; Shivas, R.G. Colletotrichum species associated with pre-and postharvest diseases of avocado and mango in eastern Australia. Australas. Plant Pathol. 2018, 47, 269–276. [Google Scholar] [CrossRef]
- Dembele, D.D.; Amari, L.N.; Camara, B.; Grechi, I.; Rey, J.; Kone, D.M. Pre and postharvest assessment of mango anthracnose incidence and severity in the north of Côte d’Ivoire. Int. J. Biol. Chem. 2020, 13, 2714–2725. [Google Scholar] [CrossRef]
- Kodituwakku, T.D.; Abeywickrama, K.; Karunanayake, K.O.L.C. Pathogenicity of stem-end rot associated fungi isolated from Karthakolomban mango and their control by spray and fumigation treatments with selected essential oils. J. Agric. Sci. 2020, 15, 19–36. [Google Scholar] [CrossRef]
- Ajitomi, A.; Minoshima, A.; Takushi, T.; Truong, H.H.; Ooshiro, A.; Yamashiro, M.; Arasaki, C.; Hirooka, Y. First report of mango (Mangifera indica) stem-end rot caused by two Diaporthe species and their susceptibility to procymidone. J. Gen. Plant Pathol. 2020, 86, 237–244. [Google Scholar] [CrossRef]
- Tovar-Pedraza, J.M.; Mora-Aguilera, J.A.; Nava-Díaz, C.; Lima, N.B.; Michereff, S.J.; Sandoval-Islas, J.S.; Câmara, M.P.S.; Téliz-Ortiz, D.; Leyva-Mir, S.G. Distribution and pathogenicity of Colletotrichum species associated with mango anthracnose in Mexico. Plant Dis. 2020, 104, 137–146. [Google Scholar] [CrossRef]
- Johnson, G.I.; Cooke, A.W.; Mead, A.J.; Wells, I.A. Stem end rot of mango in australia: Causes and control. Acta Hortic. 1991, 291, 288–295. [Google Scholar] [CrossRef]
- Peterson, R.A.; Johnson, G.I.; Schipke, L.G.; Cooke, A.W. Chemical control of stem end rot in mango. Acta Hortic. 1991, 291, 304–311. [Google Scholar] [CrossRef]
- Diskin, S.; Sharir, T.; Feygenberg, O.; Maurer, D.; Alkan, N. Fludioxonil—A potential alternative for postharvest disease control in mango fruit. Crop Prot. 2019, 124, 104855. [Google Scholar] [CrossRef]
- Galsurker, O.; Diskin, S.; Duanis-Assaf, D.; Doron-Faigenboim, A.; Maurer, D.; Feygenberg, O.; Alkan, N. Harvesting mango fruit with a short stem-end altered endophytic microbiome and reduce stem-end rot. Microorganisms 2020, 8, 558. [Google Scholar] [CrossRef]
- Govender, V.; Korsten, L. Evaluations of different formulations of Bacillus licheniformis in mango pack house trials. Biol. Control 2006, 37, 237–242. [Google Scholar] [CrossRef]
- Senghor, A.L.; Liang, W.J.; Ho, W.C. Integrated control of Colletotrichum gloeosporioides on mango fruit in Taiwan by the combination of Bacillus subtilis and fruit bagging. Biocontrol Sci. Technol. 2007, 17, 865–870. [Google Scholar] [CrossRef]
- Kefialew, Y.; Ayalew, A. Postharvest biological control of anthracnose (Colletotrichum gloeosporioides) on mango (Mangifera indica). Postharvest Biol. Technol. 2008, 50, 8–11. [Google Scholar] [CrossRef]
- Ortega Morales, B.O.; Ortega Morales, F.N.; Lara-Reyna, J.; De la Rosa-Garcia, S.C.; Martinez-Hernández, A.; Jorge Montero, M. Antagonism of Bacillus spp. Isolated from marine biofilms against terrestrial phytopathogenic fungi. Mar. Biotechnol. 2009, 11, 375–383. [Google Scholar] [CrossRef]
- Bautista-Rosales, P.U.; Calderon-Santoyo, M.; Servín-Villegas, R.; Ochoa-Alverez, N.A.; Ragazzo-Sánchez, J.A. Action mechanisms of the yeast Meyerozyma caribbica for the control of the phytopathogen Colletotrichum gloeosporioides in mangoes. Biol. Control 2013, 65, 293–301. [Google Scholar] [CrossRef]
- Akem, C.; MacManus, G.; Dann, L.; Coates, L.; Cooke, T.; Lakhesar, D. The use of plant activators in mango postharvest diseases management. Acta Hortic. 2013, 992, 369–375. [Google Scholar] [CrossRef]
- Terao, D.; Nechet, K.D.L.; Frighetto, R.T.S.; Anjos, V.D.D.A.; Benato, E.A.; Halfeld-Vieira, B.D.A. Physical postharvest treatments in the control of stem-end rot of mango. J. Phytopathol. 2018, 166, 581–589. [Google Scholar] [CrossRef]
- Diskin, S.; Feygenberg, O.; Maurer, D.; Droby, S.; Prusky, D.; Alkan, N. Microbiome alterations are correlated with occurrence of postharvest stem-end rot in mango fruit. Phytobiomes 2017, 1, 117–127. [Google Scholar] [CrossRef]
- Woolf, A.B.; Requejo-Tapia, C.; Cox, K.A.; Jackman, R.C. 1-MCP reduces physiological storage disorders of ‘Hass’ avocados. Postharvest Biol. Technol. 2005, 35, 43–60. [Google Scholar] [CrossRef]
- Nisansala, Y.M.C.; Jayakody, L.K.R.R.; Sarananda, H.A.; Somaratne, S. Effect of pre-harvest potassium treatment on stem-end rot disease development of mango (Mangifera indica L.) cv. TomEJC during fruit ripening. Sabaragamuwa Univ. J. 2015, 14, 119–132. [Google Scholar] [CrossRef]
- Montesinos-Herrero, C.; del Río, M.A.; Pastor, C.; Brunetti, O.; Palou, L. Evalua-tion of brief potassium sorbate dips to control postharvest Penicillium decay onmajor citrus species and cultivars. Postharvest Biol. Technol. 2009, 52, 117–125. [Google Scholar] [CrossRef]
- Gomba, A.; Chidamba, L.; Korsten, L. Prevalence and serovar diversity of Salmonella spp. in primary horticultural fruit production environments. Food Control 2016, 69, 13–19. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations be denaturing gradient gel electrophoresis analysis lof polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergentproperties: Endophytes and microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef]
- Gu, J.; Zhou, Z.; Li, Z.; Chen, Y.; Wang, Z.; Zhang, H. Rice (Oryza sativa L.) with reduced chlorophyll content exhibit higher photosynthetic rate and efficiency, improved canopy light distribution, and greater yields than normally pigmented plants. Field Crop Res. 2017, 200, 58–70. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 24602461. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W108–W188. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-R.; Shin, J.; Guevarra, R.B.; Lee, J.H.; Kim, D.W.; Seol, K.-H.; Lee, J.-H.; Kim, H.B.; Isaacson, R.E. Deciphering diversity indices for a better understanding of microbial communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef]
- Aleklett, K.; Hart, M.; Shade, A. The microbial ecology of flowers: An emerging frontier in phyllosphere research. Botany 2014, 92, 253–266. [Google Scholar] [CrossRef]
- Ottesen, A.R.; Gonzalez Pena, A.; White, J.R.; Pettengill, J.B.; Li, C.; Allard, S.; Rideout, S.; Allard, M.; Hill, T.; Evans, P. Baseline survey of the anatomical microbial ecology of an important food plant: Solanum lycopersicum (tomato). BMC Microbiol. 2013, 13, 114. [Google Scholar] [CrossRef]
- Pusey, P.L.; Stockwell, V.O.; Mozzola, M. Epiphytic bacteria and yeast on apple blossoms and their potential as antagonists of Erwinia amylovorai. Phytopathology 2009, 99, 571–581. [Google Scholar] [CrossRef]
- De Vega, C.; Herrera, C.M. Relationships among nectar-dwelling yeasts, flowers and ants: Patterns and incidence on nectar traits. OIKOS 2012, 121, 1878–1888. [Google Scholar] [CrossRef]
- Carmicheal, P.C.; Siyoum, N.; Chidamba, L.; Korsten, L. Exploring the microbial communities associated with Botrytis cinerea during berry development in table grape with emphasis on potential biocontrol yeasts. Eur. J. Plant Pathol. 2019, 154, 919–930. [Google Scholar] [CrossRef]
- Shade, A.; McManus, P.S.; Handelsman, J. Unexpected diversity during community succession in the apple flower microbiome. mBio 2013, 4, e00602-12. [Google Scholar] [CrossRef]
- Quintana, E.G.; Nanthacai, P.; Hiranpradit, P.; Mendoza, D.B.; Ketsa, S. Changes in mango during growth and maturation. In Mango: Fruit Development, Postharvest Physiology and Marketing in ASEAN; Mendoza, D.B., Wills, R.B.H., Eds.; ASEAN Food Handling Bureau: Kuala Lumpur, Malaysia, 1984; pp. 21–38. [Google Scholar]
- Shen, Y.; Nie, J.; Li, Z.; Li, H.; Wu, Y.; Yafeng Dong, Y.; Zhang, J. Differentiated surface fungal communities at point of harvest on apple fruits from rural and peri-urban orchards. Sci. Rep. 2018, 8, 2165. [Google Scholar] [CrossRef] [PubMed]
- Vadkertiová, R.; Molnarova, J.; Vranova, D.; Slavokova, E. Yeasts and yeast-like organisms associated with fruits and blossoms of different fruit trees. Can. J. Microbiol. 2012, 58, 1344–1352. [Google Scholar] [CrossRef]
- Martinson, E.O.; Herre, E.A.; Machado, C.A.; Arnold, A.E. Culture-free survey reveals diverse and distinctive fungal comminities associated with developing figs (Ficus spp.) in Panama. Microb. Ecol. 2012, 64, 1072–1084. [Google Scholar] [CrossRef]
- Shiea, C.; Huang, Y.L.; Liu, D.L.; Chou, C.C.; Chou, J.H.; Chen, P.Y.; Shiea, J.; Huang, M.Z. Rapid screening of residual pesticides on fruits and vegetables using thermal desorption electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2015, 29, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Vionnet, L.; De Vrieze, M.; Agnés, D.; Gfeller, A.; Luthi, A.; Haridon, F.L.; Weisskopf, L. Microbial life in the grapevine: What can we expect from the leaf microbiome? OENO One 2018, 52, 219–224. [Google Scholar] [CrossRef]
- Postmaster, A.; Sivasithamparam, K.; Turner, D.W. Enumeration and identity of microorganisms isolated from te surface of banana fruits at three developmental stages. Sci. Hortic. 1997, 69, 189–197. [Google Scholar] [CrossRef]
- Lindow, S.E.; Andersen, G.L. Influence of immigration on epiphytic bacterial populations on navel orange leaves. Appl. Environ. Microbiol. 1996, 62, 2978–2987. [Google Scholar] [CrossRef] [PubMed]
- Alkan, N.; Fortes, A.M. Insights into molecular and metabolic events associated with fruit response to post-harvest fungal pathogens. Front. Plant Sci. 2015, 6, 889. [Google Scholar] [CrossRef] [PubMed]
- Du Plooy, G.W. Aspects of Mango (Mangifera indica L.) Fruit Rind Morphology and Chemistry and Their Implication for Postharvest Quality. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2006. [Google Scholar]
- Abdelfattah, A.; Wisniewski, M.; Li Destri Nicosia, M.G.; Cacciola, S.O.; Schena, L. Metagenomic analysis of fungal diversity on strawberry plants and the effect of management practices on the fungal community structure of aerial organs. PLoS ONE 2016, 11, e0160470. [Google Scholar] [CrossRef] [PubMed]
- Stirling, A.M.; Pegg, K.; Hayward, A.C.; Stirling, G.R. Effect of copper fungicide on Colletotrichum gloeosporioides and other microorganisms on avocado leaves and fruit. Aust. J. Agric. Res. 1999, 50, 1459–1468. [Google Scholar] [CrossRef]
- Lonsdale, J.; Kotze, J. Chemical control of mango blossom diseases and the effect on fruit set and yield. Plant Dis. 1993, 77, 558. [Google Scholar] [CrossRef]
- Eilenberg, J.; Hokkanen, H.M.T. An Ecological and Societal Approach to Biological Control; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Danderson, M. Omega* (Prochloraz), a fungicide for post-harvest control of anthracnose, the dothiorella/colletotrichum complex and stem-end rot in avocados. S. Afr. Avocado Grow. Assoc. Yearb. 1986, 9, 27–30. [Google Scholar]
- Sudheer, K.P.; Chandra, P.; Gajbhiye, V.T.; Kumar, R. Uptake of benomyl fungicide from postharvest treatment and its persistance in modified atmosphere stored banana fruits. In Occupational Safety, Health & Environment and Sustainable Economic Development; Chaturvedi, P., Ed.; Concept Pubishing Company: New Delhi, India, 2007; pp. 31–36. [Google Scholar]
- Lievens, B.; Hallsworth, J.E.; Pozo, M.I.; Belgacem, Z.B.; Stevenson, A.; Willems, K.A.; Jacquemyn, H. Microbiology of sugar-rich environments: Diversity, ecology and system constraints. Environ. Microbiol. 2015, 17, 278–298. [Google Scholar] [CrossRef]
- Johnson, G.I.; Kotzé, J.M. Stem-end rot. In Compendium of Tropical Fruit Diseases; Ploetz, R.C., Zentmyer, G.A., Nishijima, W.T., Rohrbach, K.G., Ohr, H.D., Eds.; APS Press: St Paul, MN, USA, 1994; pp. 81–83. [Google Scholar]
- Mphahlele, M.P. Honey-Bee Dissemination of Bacillus subtilis to Citrus Flowers for Control of Alternaria. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 2001. [Google Scholar]
- Prakash, O.M.; Misra, A.K. Diseases of mango and their management. In Diseases of Fruits and Vegetables and Their Management; Thind, T.S., Ed.; Kalyani Publishers: Ludhiana, India, 2001; pp. 47–72. [Google Scholar]
- Nayak, G.; Altekar, N. Effect of biofield treatment on plant growth and adaptation. J. Environ. Health Sci. 2015, 1, 1–9. [Google Scholar]
- Hakizimana, J.D.; Gryzenhout, M.; Coutinho, T.A.; van den Berg, N. Endophytic diversity in Persea americana (avocado) trees and their ability to display bicontrol activity against Phytophtora cinnamomi. In Proceedings of the VII World Avocado Congress 2011, Cairns, Australia, 5–9 September 2011. [Google Scholar]
- Sanderson, P.G.; Spotts, R.A. Postharvest decay of winter pear and apple fruit caused by species of Penicillium. Phytopathology 1995, 85, 103–110. [Google Scholar] [CrossRef]
- Louw, J.P.; Korsten, L. Pathogenic Penicillium spp. on apple and pear. Plant Dis. 2014, 98, 590–598. [Google Scholar] [CrossRef]
- Kwon, J.-H.; Hong, S.-B. Soft rot of tomato caused by Mucor racemosus in Korea. Mycobiology 2005, 33, 240–242. [Google Scholar] [CrossRef]
- Saito, S.; Michhailides, T.J.; Xiao, C.L. (Mucor rot- An emerging postharvest disease of mandarin fruit caused by Mucor piriformis and other Mucor spp. in Califonia. Plant Dis. 2016, 100, 1054–1063. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef]
- Johnson, G.I.; Gosbee, M.J.; Joyce, D.C.; Irwin, J.A.G. Control of stem-end rot in mangoes. In Proceedings Mango 2000 Marketing Seminar and Production Workshop, Townsvile, QLD; Department of Primary Industries Queensland Conference and Workshop Series QC95006: Townsville, Australia, 1995; pp. 223–229. [Google Scholar]
- Prusky, D.; Kobiler, I.; Miyara, L.; Alkan, N. Fruit diseseses. In The Mango: Botany, Production and Uses; Litz, R., Ed.; CABI International: Wallingford, UK, 2009. [Google Scholar]
- Louw, J.P. Pathogenicity and Host Specificity of Penicillium spp. on Pome and Citrus Fruit. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 2014. [Google Scholar]
- Mandic-Mulec, I.; Stefanic, P.; van Elsas, J.D. Ecology of Bacillaceae. Microbiol. Spectr. 2015, 3, 1–24. [Google Scholar] [CrossRef]
- Korsten, L.; De Villiers, E.E.; Wehner, F.C.; Kotzé, J.M. Field sprays of Bacillus subtilis and fungicides for control of preharvest fruit diseases of avocado in South Africa. Plant Dis. 1997, 81, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant, developments and prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Hu, X.; Xu, X. Dispersal of Bacillus subtilis and its effect on strawberry phyllosphere microbiota under open field and protection conditions. Sci. Rep. 2016, 6, 22611. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aspect | Pre-Harvest | At Harvest | Post-Harvest | ||||
|---|---|---|---|---|---|---|---|
| Inflorescence | Small Fruit | Medium Fruit | Mature Fruit | Prochloraz Dip 1 | Cold Storage | Ripe and Ready-to-Eat | |
| Time | 0 | 100 days AFB 2 | 125 days AFB | 150 days AFB | 2 h post-harvest | 2 weeks postharvest | 4 weeks post-harvest |
| Average temperature (°C) | ±23 | ±28 | ±29.5 | ±25 | ±15 | ±8 | ±20 |
| Rainfall (mm) during sampling | 0–10 | 0–10 | 10–25 | 0–10 | - | - | - |
| Commercial intervention or observations | Insect activity | Developing fruit | Copper oxide spray 3 | Commercial harvesting | Post-harvest handling | Cold storage (8 °C) | Ripening at room temperature (20 °C) |
| Sampling Stage | TSS 1 (%) | pH | Firmness (kg) | Anthracnose Incidence (%) | Stem-End Rot Incidence (%) |
|---|---|---|---|---|---|
| At harvest | 14.8 c * | 3.8 c | 1.3 a | 0 ns | 0 b |
| Two weeks in cold storage (8 °C) | 15.9 b | 4.4 b | 0.9 b | 0 | 0 b |
| Ripe and ready-to-eat | 20.7 a | 5.9 a | 0.4 c | 0 | 30 a |
| Sampling Stages and Code | Sequences | Total OTUs | ACE | Shannon (Shannon–Weaver Diversity Index) | ||
|---|---|---|---|---|---|---|
| Stage | Type of Stage | Code 1 | ||||
| Pre-harvest | Inflorescence | FP | 73,734 | 183 | 126.9 | 1.76 |
| Small fruit | FP | 98,043 | 250 | 114.3 | 1.32 | |
| S | 53,907 | 354 | 83.8 | 1.32 | ||
| SEP | 80,913 | 323 | 91.5 | 1.48 | ||
| Medium fruit | FP | 71,619 | 241 | 115.7 | 1.92 | |
| S | 46,462 | 364 | 77.2 | 0.83 | ||
| SEP | 128,211 | 268 | 105.8 | 1.80 | ||
| At harvest | Mature fruit | FP | 88,454 | 299 | 109.9 | 1.79 |
| S | 37,464 | 315 | 102.7 | 1.83 | ||
| SEP | 90,168 | 289 | 110.8 | 2.39 | ||
| Post-harvest | After prochloraz packhouse treatment | FP | 60,126 | 245 | 105.5 | 1.19 |
| SEP | 92,011 | 332 | 79.7 | 0.89 | ||
| Two weeks cold storage (8 °C) | FP | 75,921 | 204 | 118.1 | 1.97 | |
| SEP | 205,514 | 304 | 76.7 | 0.52 | ||
| Ready-to-eat stage | FP | 66,709 | 198 | 122.8 | 2.40 | |
| SEP | 189,201 | 316 | 81.8 | 0.86 | ||
| Sampling Stages and Code | Sequences | Total OTUs | ACE | Shannon–Weaver Diversity Index | ||
|---|---|---|---|---|---|---|
| Stage | Type of Stage | Code 1 | ||||
| Pre-harvest | Inflorescence | FP | 64,745 | 239 | 102.6 | 1.19 |
| Small fruit | FP | 81,349 | 352 | 90.8 | 1.17 | |
| S | 54,412 | 425 | 100.4 | 1.16 | ||
| SEP | 105,422 | 421 | 97.2 | 2.62 | ||
| Medium fruit | FP | 76,837 | 313 | 101.4 | 2.32 | |
| S | 52,145 | 408 | 83.4 | 1.14 | ||
| SEP | 57,495 | 414 | 83.9 | 1.13 | ||
| At harvest | Mature fruit | FP | 75,359 | 324 | 98.0 | 1.73 |
| S | 58,768 | 418 | 91.3 | 1.24 | ||
| SEP | 60,271 | 416 | 87.6 | 1.44 | ||
| Post-harvest | After prochloraz packhouse treatment | FP | 93,785 | 339 | 94.1 | 1.52 |
| SEP | 4893 | 392 | 83.9 | 1.11 | ||
| Two weeks cold storage (8 °C) | FP | 99,451 | 348 | 92.1 | 1.70 | |
| SEP | 114,287 | 376 | 88.4 | 1.978 | ||
| Ready-to-eat stage | FP | 51,933 | 314 | 90.6 | 0.76. | |
| SEP | 68,745 | 382 | 92.4 | 2.27 | ||
| Sampling Stage | Pre-Harvest | At Harvest | Post-Harvest | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| InfloRescence | Small Fruit | Medium Fruit | Mature Fruit: Point of Harvest | After Prochloraz Dip | Two Weeks Cold in Storage (8 °C) | Ripe and Ready-to-Eat | ||||||||||
| FP 1 | FP | S | SEP | FP | S | SEP | FP | S | SEP | FP | SEP | FP | SEP | FP | SEP | |
| Stem-end rot causing fungi (%) | ||||||||||||||||
| Botryosphaeria | 0.43 | 0.63 | - | 0.01 | 0.01 | - | 0.01 | 0.16 | 0.16 | 0.02 | 0.02 | 0.74 | 0.10 | 21.37 | 3.13 | 21.57 |
| Cladosporium | 1.04 | 0.83 | 0.01 | 0.04 | 0.11 | - | 0.02 | 1.13 | 0.08 | 0.10 | 0.04 | 0.13 | 0.34 | - | 0.12 | - |
| Aspergillus | - | 0.10 | - | - | 0.01 | - | 0.02 | - | - | - | - | 2.22 | 0.02 | - | 0.01 | - |
| Penicillium | 6.53 | 47.65 | 1.87 | 72.61 | 10.56 | 46.24 | 98.61 | 17.85 | 1.87 | 12.78 | 8.17 | 30.55 | 17.16 | 28.21 | 24.04 | 29.59 |
| Alternaria | 1.13 | 5.53 | - | 0.01 | 0.17 | 0.09 | 0.06 | 4.34 | 0.08 | 0.07 | 0.74 | 0.13 | - | 0.12 | 0.10 | - |
| Other pathogenic fungi (%) | ||||||||||||||||
| Colletotrichum | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| Phoma | 7.79 | 2.29 | 0.01 | - | 0.02 | 0.05 | 0.05 | 0.48 | 0.11 | 0.02 | 0.04 | 0.20 | 0.29 | 0.49 | 0.29 | 0.15 |
| Mucor | 0.39 | 3.55 | 0.09 | 0.08 | 0.10 | 0.09 | 0.09 | 0.48 | 0.27 | 0.43 | 0.33 | 0.74 | 24.25 | 2.44 | 15.67 | 2.04 |
| Fusarium | 0.43 | 1.25 | 0.05 | 0.04 | 1.18 | 0.05 | 0.01 | 0.80 | 1.37 | 0.48 | 0.02 | 0.20 | 1.22 | 0.37 | 2.61 | 1.17 |
| Beneficial bacteria (%) | ||||||||||||||||
| Bacillus | 1.23 | 66.49 | 0.47 | 0.79 | 1.11 | 0.98 | 0.93 | 1.87 | 0.31 | 1.78 | 0.76 | 0.37 | 0.20 | 0.75 | 2.33 | 0.66 |
| Bacillus subtilis | 0.06 | 0.04 | 0.22 | 0.13 | - | 0.09 | 0.25 | 0.15 | 0.07 | 0.15 | 0.04 | 0.03 | 0.01 | 0.12 | 0.02 | 0.13 |
| Lactobacillus | 0.62 | 0.01 | - | - | 0.08 | - | - | - | - | 0.02 | - | - | - | - | - | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bill, M.; Chidamba, L.; Gokul, J.K.; Korsten, L. Mango Endophyte and Epiphyte Microbiome Composition during Fruit Development and Post-Harvest Stages. Horticulturae 2021, 7, 495. https://doi.org/10.3390/horticulturae7110495
Bill M, Chidamba L, Gokul JK, Korsten L. Mango Endophyte and Epiphyte Microbiome Composition during Fruit Development and Post-Harvest Stages. Horticulturae. 2021; 7(11):495. https://doi.org/10.3390/horticulturae7110495
Chicago/Turabian StyleBill, Malick, Lizyben Chidamba, Jarishma Keriuscia Gokul, and Lise Korsten. 2021. "Mango Endophyte and Epiphyte Microbiome Composition during Fruit Development and Post-Harvest Stages" Horticulturae 7, no. 11: 495. https://doi.org/10.3390/horticulturae7110495
APA StyleBill, M., Chidamba, L., Gokul, J. K., & Korsten, L. (2021). Mango Endophyte and Epiphyte Microbiome Composition during Fruit Development and Post-Harvest Stages. Horticulturae, 7(11), 495. https://doi.org/10.3390/horticulturae7110495