
Transcriptomic Analysis of Hardened ‘Suli’ Pear (Pyrus bretschneideri Rehd) for Identification of Key Genes for Lignin Biosynthesis and Accumulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fruit Materials
2.2. Observation of Stone Cells
2.3. Lignin Level Measurement
2.4. Extraction of Total RNA and Analysis of RNA-Seq Data
2.5. Quantitative Real-Time PCR Analysis (qRT-PCR)
2.6. Processing of Data
3. Results
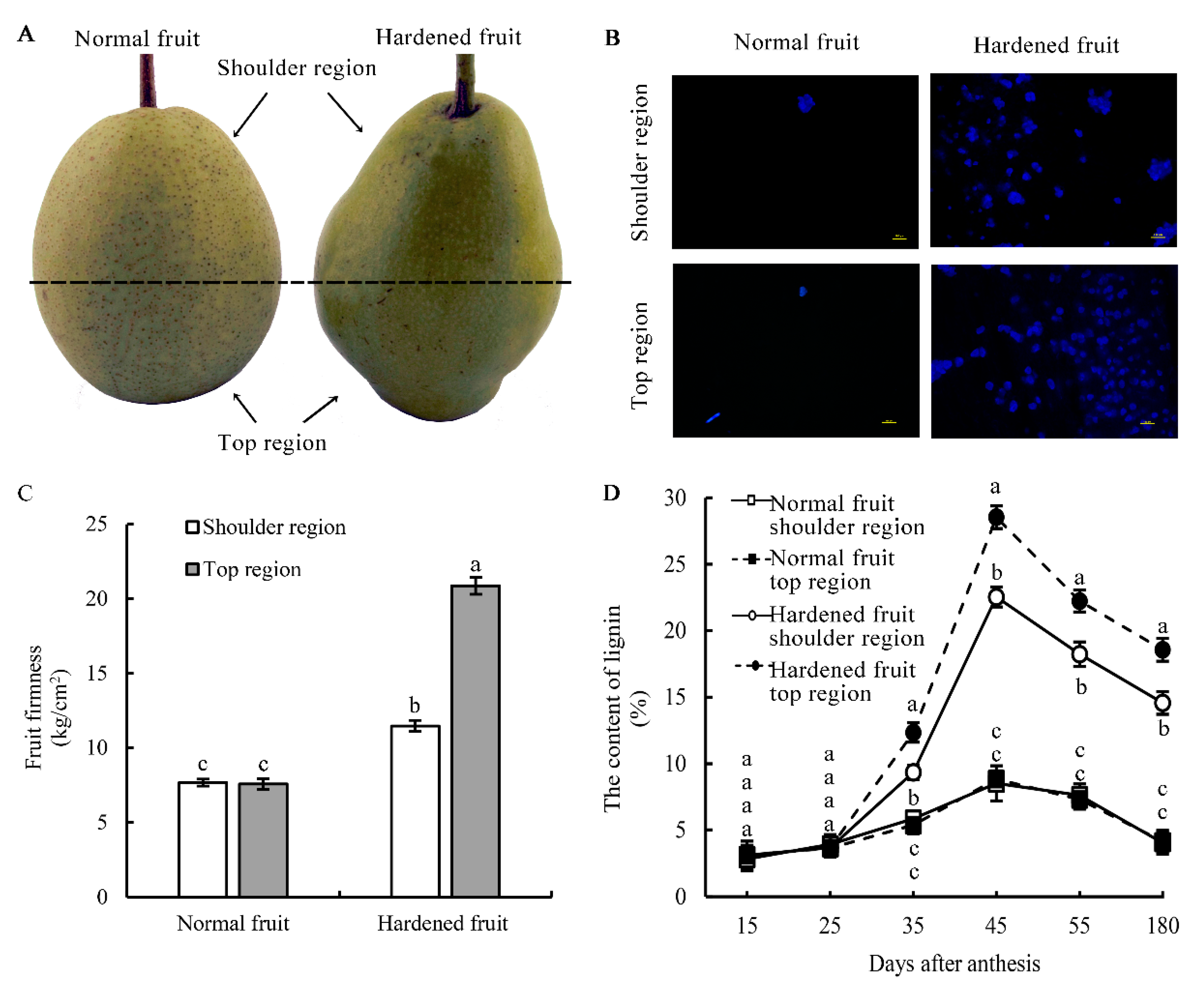
3.1. Lignin Level in HFs at Varying Developmental Stages
3.2. Library Establishement and Transcriptome Sequencing
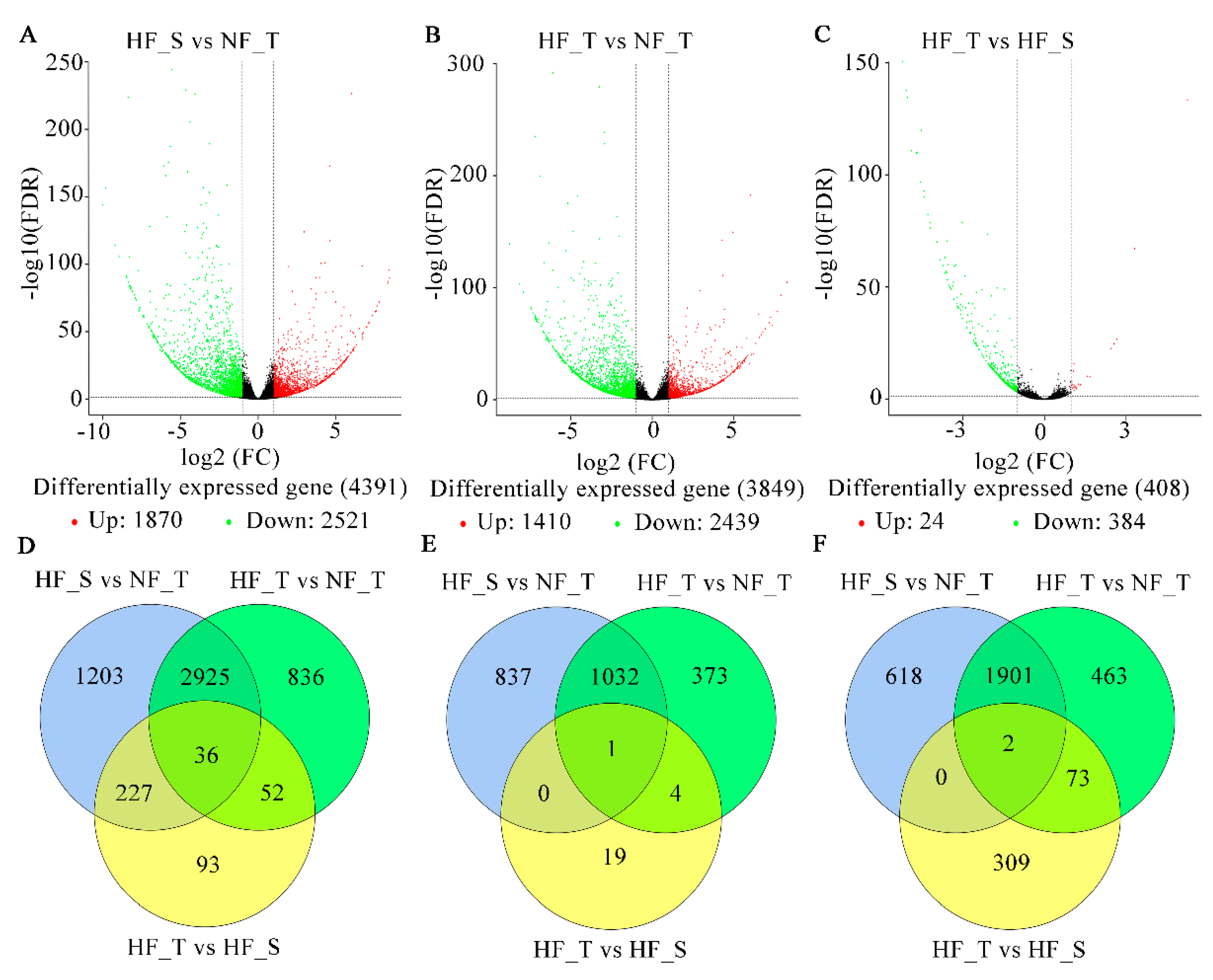
3.3. Identification of DEGs
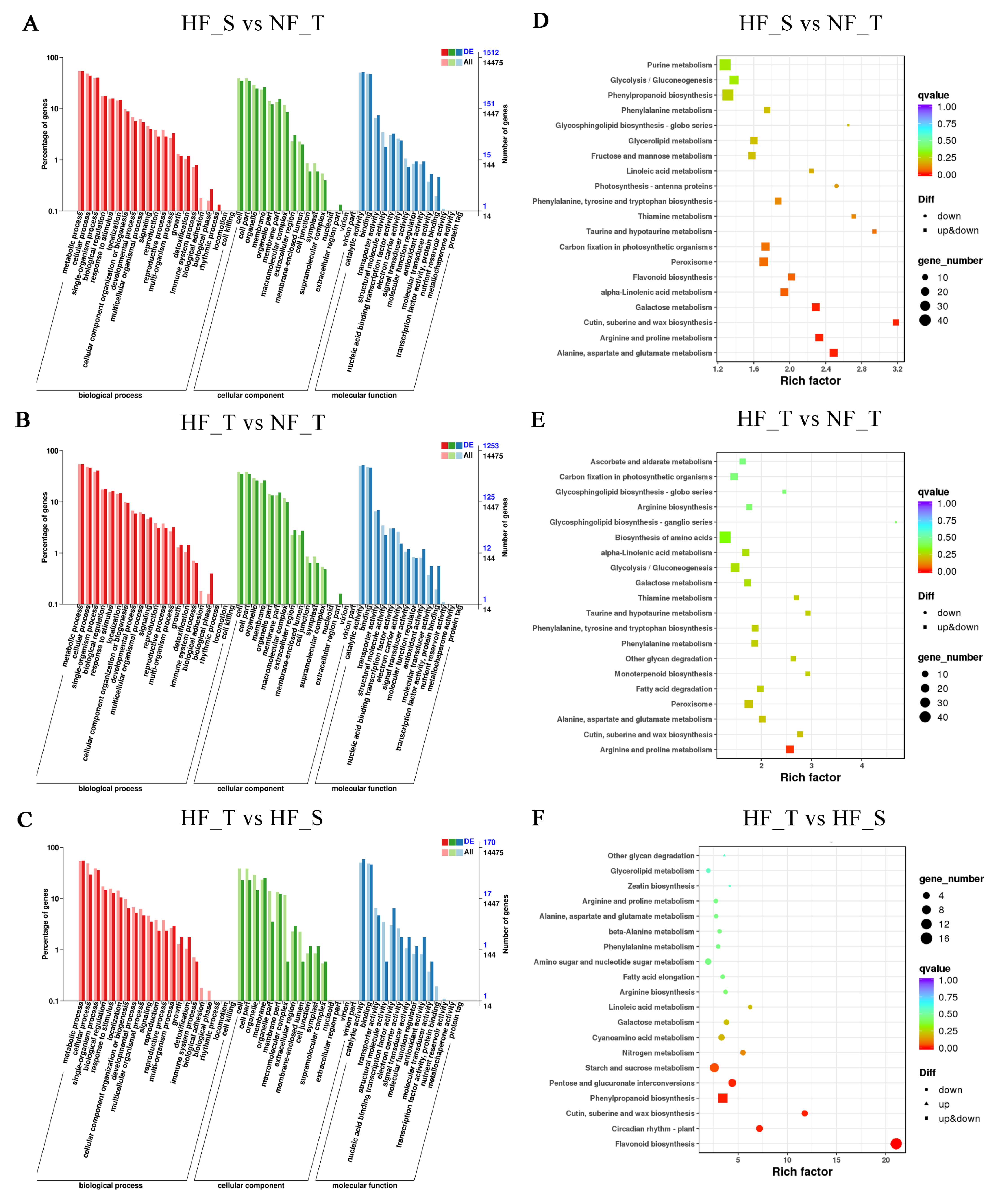
3.4. Gene Ontology (GO) Enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis of the DEGs
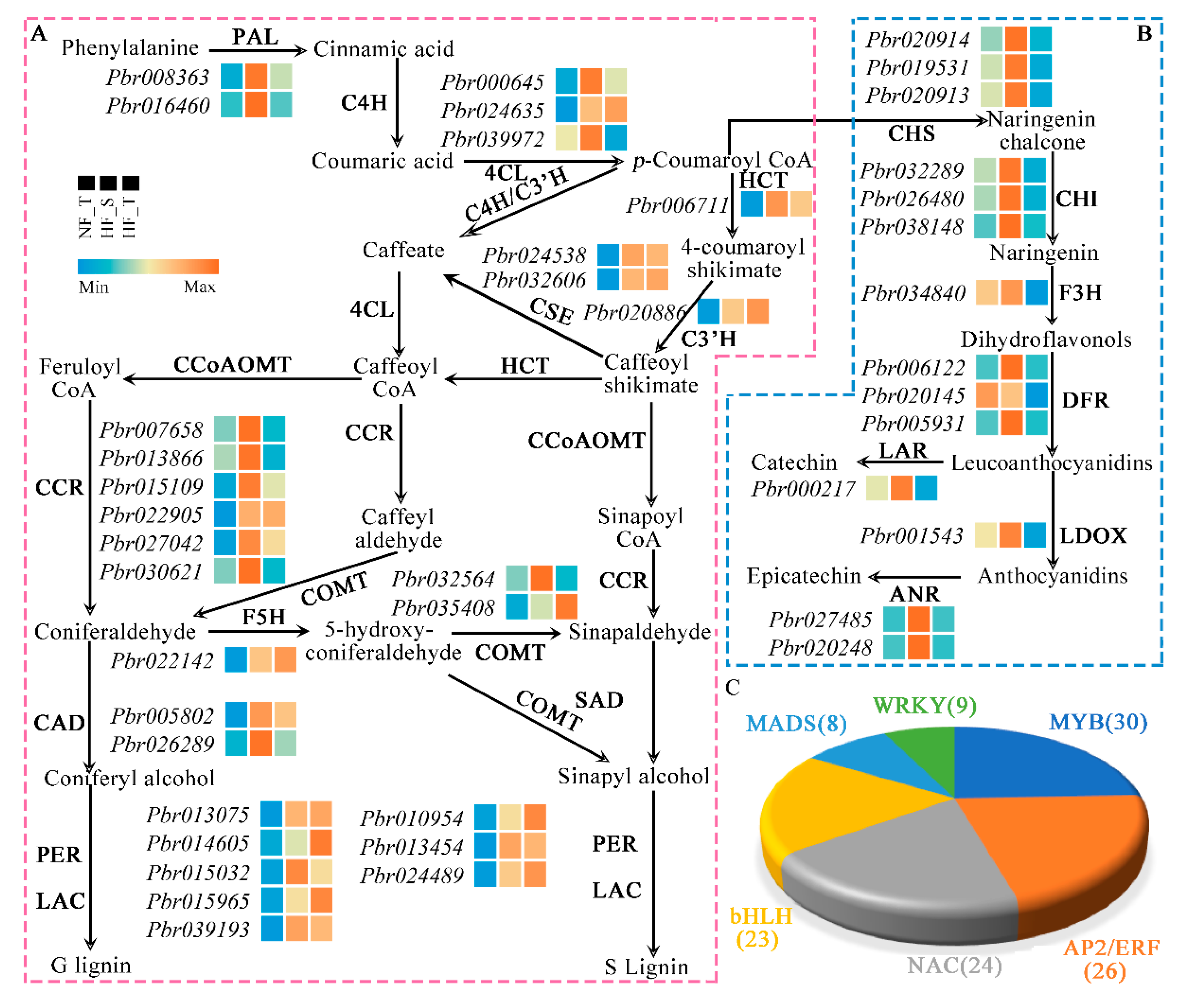
3.5. DEGs Related to Lignin Biosynthesis between NFs and HFs
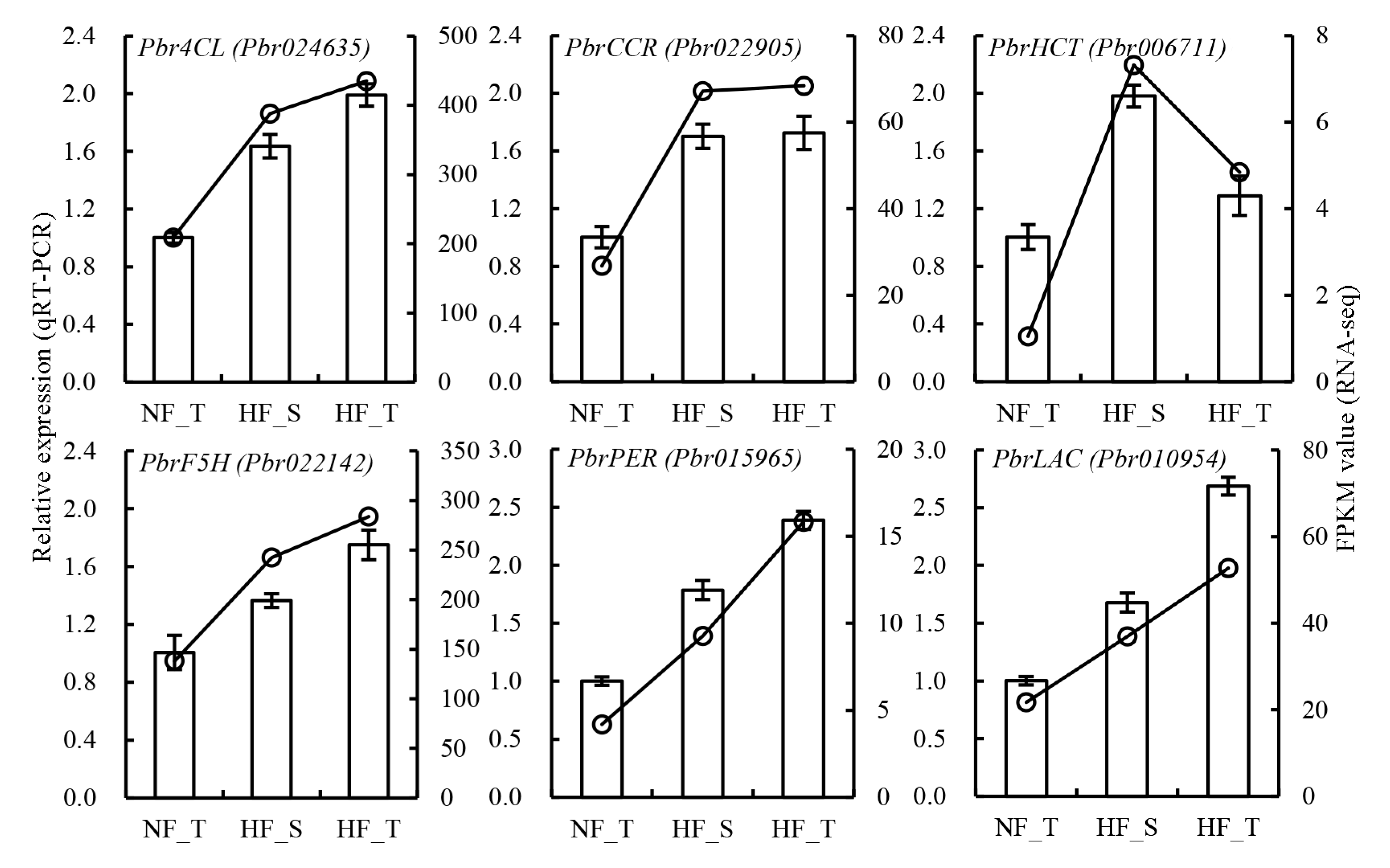
3.6. qRT-PCR Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, G.Q.; Li, W.S.; Zheng, P.H.; Xu, T.; Chen, L.J.; Liu, D.F.; Hussain, S.; Teng, Y.W. Transcriptomic analysis of ‘Suli’ pear (Pyrus pyrifolia white pear group) buds during the dormancy by RNA-Seq. BMC Genom. 2012, 13, 700. [Google Scholar] [CrossRef]
- Liu, X.F.; Li, S.R.; Feng, X.X.; Li, L.L. Study on cell wall composition, fruit quality and tissue structure of hardened ‘Suli’ pears (Pyrus bretschneideri Rehd). J. Plant Growth Regul. 2020, 40, 2007–2016. [Google Scholar] [CrossRef]
- Yamamoto, T.; Watanabe, S. Initial time of development of hard end disorder in ‘Bartlett’ pear. J. Jpn. Soc. Hort. Sci. 1982, 51, 42–151. [Google Scholar] [CrossRef][Green Version]
- Pierson, C.F.; Ceponis, M.J.; Mccolloch, L.P. Market Diseases of Apples, Pears, and Quinces; Agriculture Handbook; USDA: Washington, DC, USA, 1971; No. 376. [Google Scholar]
- Raese, J.T.; Drake, S.R. Calcium foliar sprays for control of alfalfa greening, cork spot, and hard end in ‘Anjou’ pears. J. Plant Nutr. 2006, 29, 543–552. [Google Scholar] [CrossRef]
- Fumio, T. Recent advances in research on Japanese pear rootstocks. J. Jpn. Soc. Hort. Sci. 2012, 81, 1–10. [Google Scholar]
- Raese, J.T. Fruit disorders, mineral composition and tree performance influenced by rootstocks of ‘Anjou’ pears. Acta Hortic. 1994, 367, 372–379. [Google Scholar] [CrossRef]
- Wang, Y.L.; Zhang, X.F.; Wang, Y.Z.; Yang, S.L.; Qu, H.Y. The changes of intracellular calcium concentration and distribution in the hard end pear (Pyrus pyrifolia cv. ‘Whangkeumbae’) fruit. Cell Calcium 2018, 71, 15–23. [Google Scholar] [CrossRef]
- Lu, G.L.; Li, Z.J.; Zhang, X.F.; Wang, R.; Yang, S.L. Expression analysis of lignin-associated genes in hard end pear (Pyrus pyrifolia Whangkeumbae) and its response to calcium chloride treatment conditions. J. Plant Growth Regul. 2015, 34, 251–262. [Google Scholar] [CrossRef]
- Chen, H.J.; Cao, S.F.; Fang, X.J.; Mu, H.L.; Yang, H.L.; Wang, X.; Xu, Q.Q.; Gao, H.Y. Changes in fruit firmness, cell wall composition and cell wall degrading enzymes in postharvest blueberries during storage. Sci. Hortic. 2015, 188, 44–48. [Google Scholar] [CrossRef]
- Singh, R.; Rastogi, S.; Dwivedi, U.N. Phenylpropanoid metabolism in ripening fruits. Compr. Rev. Food Sci. Food Saf. 2010, 9, 398–416. [Google Scholar] [CrossRef]
- Dixon, R.A.; Barros, J. Lignin biosynthesis: Old roads revisited and new roads explored. Open Biol. 2019, 9, 190215. [Google Scholar] [CrossRef]
- Yoon, J.; Choi, H.; An, G. Roles of lignin biosynthesis and regulatory genes in plant development. J. Integr. Plant Biol. 2015, 57, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.P.; Li, G.Q.; Nie, J.Q.; Lin, Y.; Nie, F.; Zhang, J.Y.; Xu, Y.L. Study of the structure and biosynthetic pathway of lignin in stone cells of pear. Sci. Hortic. 2010, 125, 374–379. [Google Scholar] [CrossRef]
- Yan, C.C.; Yin, M.; Zhang, N.; Jin, Q.; Fang, Z.; Lin, Y.; Cai, Y.P. Stone cell distribution and lignin structure in various pear varieties. Sci. Hortic. 2014, 174, 142–150. [Google Scholar] [CrossRef]
- Su, X.Q.; Zhao, Y.; Wang, H.; Li, G.H.; Cheng, X.; Jin, Q.; Cai, Y.P. Transcriptomic analysis of early fruit development in Chinese white pear (Pyrus bretschneideri Rehd.) and functional identification of PbCCR1 in lignin biosynthesis. BMC Plant Biol. 2019, 19, 417. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Cheng, X.; Jin, Q.; Su, X.Q.; Li, M.L.; Yan, C.C.; Jiao, X.Y.; Li, D.H.; Lin, Y.; Cai, Y.P. Comparison of the transcriptomic analysis between two Chinese white pear (Pyrus bretschneideri Rehd.) genotypes of different stone cells contents. PLoS ONE 2017, 12, e0187114. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.P.; Liu, T.T.; Hu, C.H.; Song, Y.Q.; Hao, R.J.; Feng, X.X.; Cui, T.T.; Han, Y.Z.; Li, L.L. Comparative analysis of transcriptomic profiling to identify genes involved in the bulged surface of pear fruit (Pyrus bretschneideri Rehd. cv. Yuluxiangli). Physiol. Mol. Biol. Plants 2021, 27, 69–80. [Google Scholar] [CrossRef]
- Wang, Q.; Wu, X.Y.; Liu, L.; Yao, D.Z.; Li, J.C.; Fang, J.C.; Chen, X.N.; Zhu, L.W.; Liu, P.; Ye, Z.F.; et al. Transcriptome and metabolomic analysis to reveal the browning spot formation of ‘Huangguan’ pear. BMC Plant Biol. 2021, 21, 321. [Google Scholar]
- Mamat, A.; Tusong, K.; Xu, J.; Yan, P.; Mei, C.; Wang, J.X. Integrated transcriptomic and proteomic analysis reveals the complex molecular mechanisms underlying stone cell formation in Korla pear. Sci. Rep. 2021, 11, 7688. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, Z.W.; Shi, Z.B.; Zhang, S.; Ming, R.; Zhu, S.L.; Khan, M.A.; Tao, S.T.; Korban, S.S.; Wang, H.; et al. The genome of the pear (Pyrus bretschneideri Rehd). Genome Res. 2013, 23, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Boudet, A.M.; Kajita, S.; Grima-Pettenati, J.; Goffner, D. Lignins and lignocellulosics: A better control of synthesis for new and improved uses. Trends Plant Sci. 2003, 8, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.K.; Li, X.; Zhang, J.; Ge, H.; Yin, X.R.; Chen, K.S. Regulation of loquat fruit low temperature response and lignification involves interaction of heat shock factors and genes associated with lignin biosynthesis. Plant Cell Environ. 2016, 39, 1780–1789. [Google Scholar] [CrossRef] [PubMed]
- Ketsa, S.; Koolpluksee, M. Some physical and biochemical characteristics of damaged pericarp of mangosteen fruit after impact. Postharvest Biol. Technol. 1993, 2, 209–215. [Google Scholar] [CrossRef]
- Bunsiri, A.; Ketsa, S.; Paull, R.E. Phenolic metabolism and lignin synthesis in damaged pericarp of mangosteen fruit after impact. Postharvest Biol. Technol. 2003, 29, 61–71. [Google Scholar] [CrossRef]
- Deng, L.L.; Cui, W.J.; Yao, S.X.; Zeng, K.F. Effect of mechanical damage on organizational structure of postharvest navel orange rind. Food Sci. 2019, 40, 118–126. [Google Scholar]
- Zhou, X.Y.; Yue, J.Q.; Yang, H.B.; Zhu, C.H.; Zhu, F.; Li, J.X.; Xu, R.W.; Gao, J.Y.; Zhou, D.G.; Deng, X.X.; et al. Integration of metabolome, histochemistry and transcriptome analysis provides insights into lignin accumulation in oleocellosis-damaged flavedo of citrus fruit. Postharvest Biol. Technol. 2021, 172, 111362. [Google Scholar] [CrossRef]
- Zheng, J.; Liu, L.B.; Tao, H.H.; An, Y.Y.; Wang, L.J. Transcriptomic profiling of apple calli with a focus on the key genes for ALA-induced anthocyanin accumulation. Front. Plant Sci. 2021, 12, 640606. [Google Scholar] [CrossRef] [PubMed]
- Syros, T.; Yupsanis, T.; Zafiriadis, H.; Economou, A. Activity and isoforms of peroxidases, lignin and anatomy, during adventitious rooting in cuttings of Ebenus cretica L. J. Plant Physiol. 2004, 161, 69–77. [Google Scholar] [CrossRef]
- Jaakola, L.; Pirttila, A.M.; Halonen, M.; Hohtola, A. Isolation of high quality RNA from bilberry (Vaccinium myrtillus L.) fruit. Mol. Biotechnol. 2001, 19, 201–203. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Goujon, T.; Sibout, R.; Eudes, A.; Mackay, J.; Jouanin, L. Genes involved in the biosynthesis of lignin precursors in Arabidopsis thaliana. Plant Physiol. Biochem. 2003, 41, 677–687. [Google Scholar] [CrossRef]
- Lu, J.Y.; Shi, Y.Z.; Li, W.J.; Chen, S.; Wang, Y.F.; He, X.L.; Yin, X.G. RcPAL, a key gene in lignin biosynthesis in Ricinus communis L. BMC Plant Biol. 2019, 19, 181. [Google Scholar]
- Li, X.; Bonawitz, N.D.; Weng, J.K.; Chapple, C. The growth reduction associated with repressed lignin biosynthesis in Arabidopsis thaliana is independent of flavonoids. Plant Cell 2010, 22, 1620–1632. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Cesarino, I.; Rataj, K.; Xiao, Y.G.; Sundin, L.; Goeminne, G.; Kim, H.; Cross, J.; Morreel, K.; Araujo, P.; et al. Caffeoyl shikimate esterase (CSE) is an enzyme in the lignin biosynthetic pathway in Arabidopsis. Science 2013, 341, 1103–1106. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Li, M.L.; Li, D.H.; Zhang, J.Y.; Jin, Q.; Sheng, L.L.; Cai, Y.P.; Lin, Y. Characterization and analysis of CCR and CAD gene families at the whole-genome level for lignin synthesis of stone cells in pear (Pyrus bretschneideri) fruit. Biol. Open 2017, 6, 1602–1613. [Google Scholar] [CrossRef]
- Rahantamalala, A.; Rech, P.; Martinez, Y.; Chaubet-Gigot, N.; Grima-Pettenati, J.; Pacquit, V. Coordinated transcriptional regulation of two key genes in the lignin branch pathway—CAD and CCR—is mediated through MYB-binding sites. BMC Plant Biol. 2010, 10, 130. [Google Scholar] [CrossRef] [PubMed]
- Sirisha, V.L.; Prashant, S.; Kumar, D.R.; Pramod, S.; Jalaja, N.; Kumari, P.H.; Rao, P.M.; Rao, S.N.; Mishra, P.; Karumanchi, S.R.; et al. Cloning, characterization and impact of up- and down-regulating subabul cinnamyl alcohol dehydrogenase (CAD) gene on plant growth and lignin profiles in transgenic tobacco. Plant Growth Regul. 2012, 66, 239–253. [Google Scholar] [CrossRef]
- Weng, J.K.; Mo, H.; Chapple, C. Over-expression of F5H in COMT-deficient Arabidopsis leads to enrichment of an unusual lignin and disruption of pollen wall formation. Plant J. 2010, 64, 898–911. [Google Scholar] [CrossRef] [PubMed]
- Berthet, S.; Demont-Caulet, N.; Pollet, B.; Bidzinski, P.; Cézard, L.; Le Bris, P.; Borrega, N.; Hervé, J.; Blondet, E.; Balzergue, S.; et al. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems. Plant Cell 2011, 23, 1124–1137. [Google Scholar] [CrossRef]
- Cosio, C.; Ranocha, P.; Francoz, E.; Burlat, V.; Zheng, Y.; Perry, S.E.; Ripoll, J.J.; Yanofsky, M.; Dunand, C. The class III peroxidase PRX17 is a direct target of the MADS-box transcription factor AGAMOUS-LIKE15 (AGL15) and participates in lignified tissue formation. New Phytol. 2017, 213, 250–263. [Google Scholar] [CrossRef]
- Cao, Y.P.; Han, Y.H.; Meng, D.D.; Li, D.H.; Jin, Q.; Lin, Y.; Cai, Y.P. Structural, Evolutionary, and Functional Analysis of the Class III Peroxidase Gene Family in Chinese Pear (Pyrus bretschneideri). Front. Plant Sci. 2016, 7, 1874. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Dixon, R.A. Transcriptional networks for lignin biosynthesis: More complex than we thought? Trends Plant Sci. 2011, 16, 227–233. [Google Scholar] [CrossRef]
- Zhou, J.L.; Lee, C.H.; Zhong, R.Q.; Ye, Z.H. MYB58 and MYB63 are transcriptional activators of the lignin biosynthetic pathway during secondary cell wall formation in Arabidopsis. Plant Cell 2009, 21, 248–266. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, R.L.; Zhong, R.Q.; Ye, Z.H. MYB83 is a direct target of SND1 and acts redundantly with MYB46 in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell Physiol. 2009, 50, 1950–1964. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.Q.; Richardson, E.A.; Ye, Z.H. The MYB46 Transcription factor is a direct target of SND1 and regulates secondary wall biosynthesis in Arabidopsis. Plant Cell 2007, 19, 2776–2792. [Google Scholar] [CrossRef] [PubMed]
- Bomal, C.; Bedon, F.; Caron, S.; Mansfield, S.D.; Levasseur, C.; Cooke, J.E.; Blais, S.; Tremblay, L.; Morency, M.J.; Pavy, N.; et al. Involvement of Pinus taeda MYB1 and MYB8 in phenylpropanoid metabolism and secondary cell wall biogenesis: A comparative in planta analysis. J. Exp. Bot. 2008, 59, 3925–3939. [Google Scholar] [CrossRef] [PubMed]
- Patzlaff, A.; Newman, L.J.; Dubos, C.; Whetten, R.W.; Smith, C.; McInnis, S.; Bevan, M.W.; Sederoff, R.R.; Campbell, M.M. Characterisation of PtMYB1, an R2R3-MYB from pine xylem. Plant Mol. Biol. 2003, 53, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Goicoechea, M.; Lacombe, E.; Legay, S.; Mihaljevic, S.; Grima-Pettenati, J. EgMYB2, a new transcriptional activator from Eucalyptus xylem, regulates secondary cell wall formation and lignin biosynthesis. Plant J. 2010, 43, 553–567. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, R.L.; Zhong, R.; Fowler, S.; Lyskowski, D.; Piyasena, H.; Carleton, K.; Spicer, C.; Ye, Z.H. The poplar MYB transcription factors, PtrMYB3 and PtrMYB20, are involved in the regulation of secondary wall biosynthesis. Plant Cell Physiol. 2010, 51, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Lee, C.; Zhou, J.; McCarthy, R.L.; Ye, Z.H. A battery of transcription factors involved in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell 2008, 20, 2763–2782. [Google Scholar] [CrossRef]
- Li, M.T.; Cheng, C.X.; Zhang, X.F.; Zhou, S.P.; Wang, C.H.; Ma, C.H.; Yang, S.L. PpNAC187 enhances lignin synthesis in ‘Whangkeumbae’ pear (Pyrus pyrifolia) ‘hard-end’ fruit. Molecules 2019, 24, 4338. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Li, S.J.; Liu, X.F.; Yin, X.R.; Grierson, D.; Chen, K.S. Ternary complex EjbHLH1-EjMYB2-EjAP2-1 retards low temperature-induced flesh lignification in loquat fruit. Plant Physiol. Biochem. 2019, 139, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.K.; Li, X.; Xu, Q.; Chen, J.Y.; Yin, X.R.; Ferguson, I.B.; Chen, K.S. EjAP2-1, an AP2/ERF gene, is a novel regulator of fruit lignification induced by chilling injury, via interaction with EjMYB transcription factors. Plant Biotechnol. J. 2015, 13, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Besseau, S.; Hoffmann, L.; Geoffroy, P.; Lapierre, C.; Legrand, M. Flavonoid accumulation in Arabidopsis repressed in lignin synthesis affects auxin transport and plant growth. Plant Cell 2007, 19, 148–162. [Google Scholar] [CrossRef]
- Hoffmann, L.; Besseau, S.; Geoffroy, P.; Ritzenthaler, C.; Meyer, D.; Lapierre, C.; Pollet, B.; Legrand, M. Silencing of hydroxycinnamoyl-coenzyme a shikimate/quinate hydroxycinnamoyltransferase affects phenylpropanoid biosynthesis. Plant Cell 2004, 16, 1446–1465. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; Liu, X.; Jia, Y.; Liu, H.; Li, L. Transcriptomic Analysis of Hardened ‘Suli’ Pear (Pyrus bretschneideri Rehd) for Identification of Key Genes for Lignin Biosynthesis and Accumulation. Horticulturae 2021, 7, 467. https://doi.org/10.3390/horticulturae7110467
Feng X, Liu X, Jia Y, Liu H, Li L. Transcriptomic Analysis of Hardened ‘Suli’ Pear (Pyrus bretschneideri Rehd) for Identification of Key Genes for Lignin Biosynthesis and Accumulation. Horticulturae. 2021; 7(11):467. https://doi.org/10.3390/horticulturae7110467
Chicago/Turabian StyleFeng, Xinxin, Xiaofeng Liu, Yuxiu Jia, Huifang Liu, and Liulin Li. 2021. "Transcriptomic Analysis of Hardened ‘Suli’ Pear (Pyrus bretschneideri Rehd) for Identification of Key Genes for Lignin Biosynthesis and Accumulation" Horticulturae 7, no. 11: 467. https://doi.org/10.3390/horticulturae7110467
APA StyleFeng, X., Liu, X., Jia, Y., Liu, H., & Li, L. (2021). Transcriptomic Analysis of Hardened ‘Suli’ Pear (Pyrus bretschneideri Rehd) for Identification of Key Genes for Lignin Biosynthesis and Accumulation. Horticulturae, 7(11), 467. https://doi.org/10.3390/horticulturae7110467