
Potential Bacterial Antagonists for the Control of Charcoal Rot (Macrophomina phaseolina) in Strawberry

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Collection
2.2. Fungal Culture
2.3. Antifungal Activity of Bacterial Strains against M. phaseolina
2.4. Molecular Characterization of Bacteria
2.5. Biocontrol Related Enzymatic Activity and Hydrogen Cyanide (HCN) Production
2.6. Biocontrol Efficacy against Charcoal Rot Disease on Strawberry Plants
2.7. Growth Promotion in Greenhouse Trials
2.8. Field Trials
2.8.1. Fruit Weight
2.8.2. Plant Mortality and Charcoal Rot Incidence in Dead Plants
2.8.3. Fruit Quality
2.8.4. Fruit Rots
2.8.5. Fruits Firmness
2.8.6. Soluble Solid Content
2.8.7. Anthocyanins
2.8.8. Ascorbic Acid (Vitamin C)
2.9. Statistical Analyses
3. Results
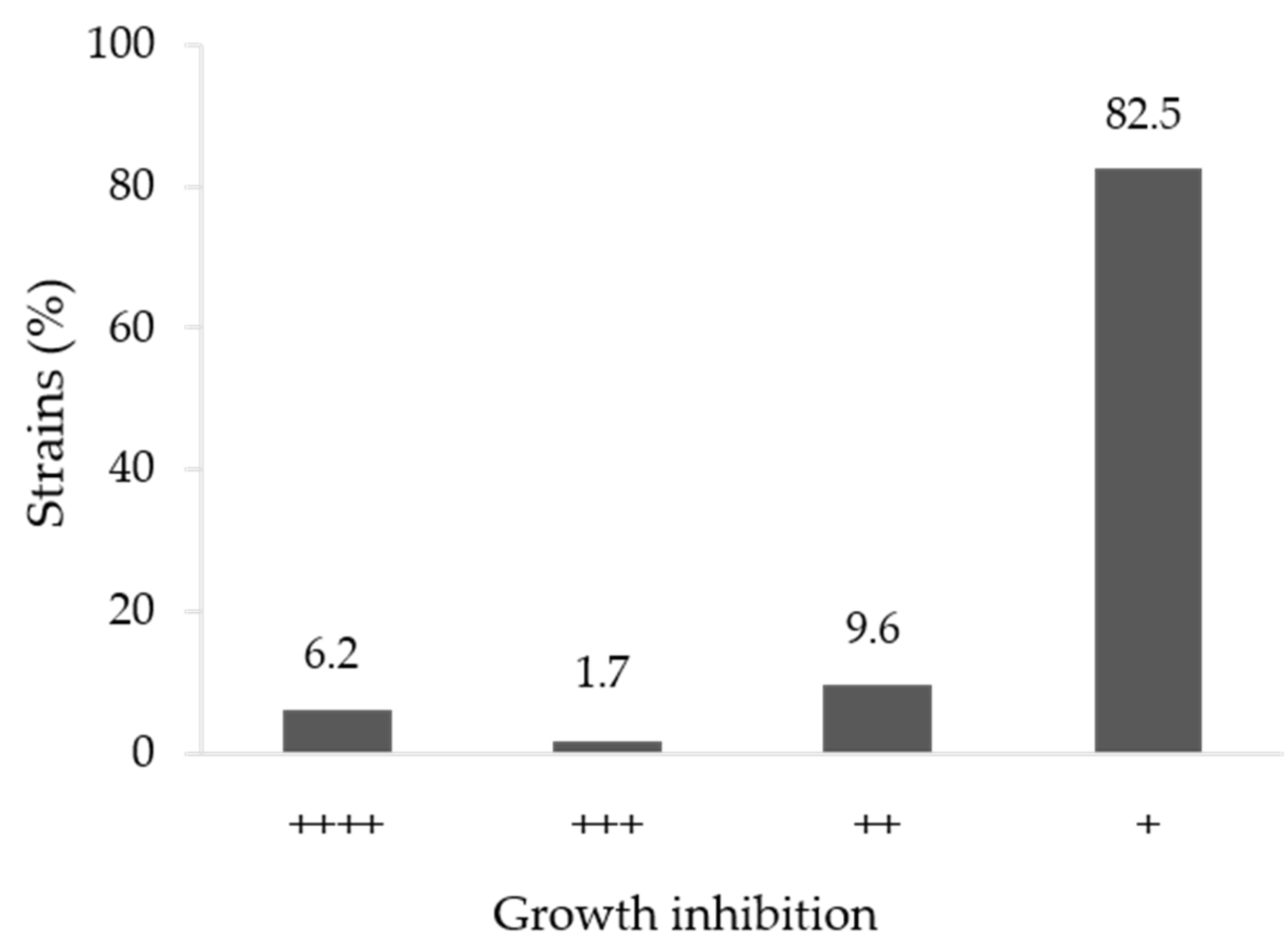
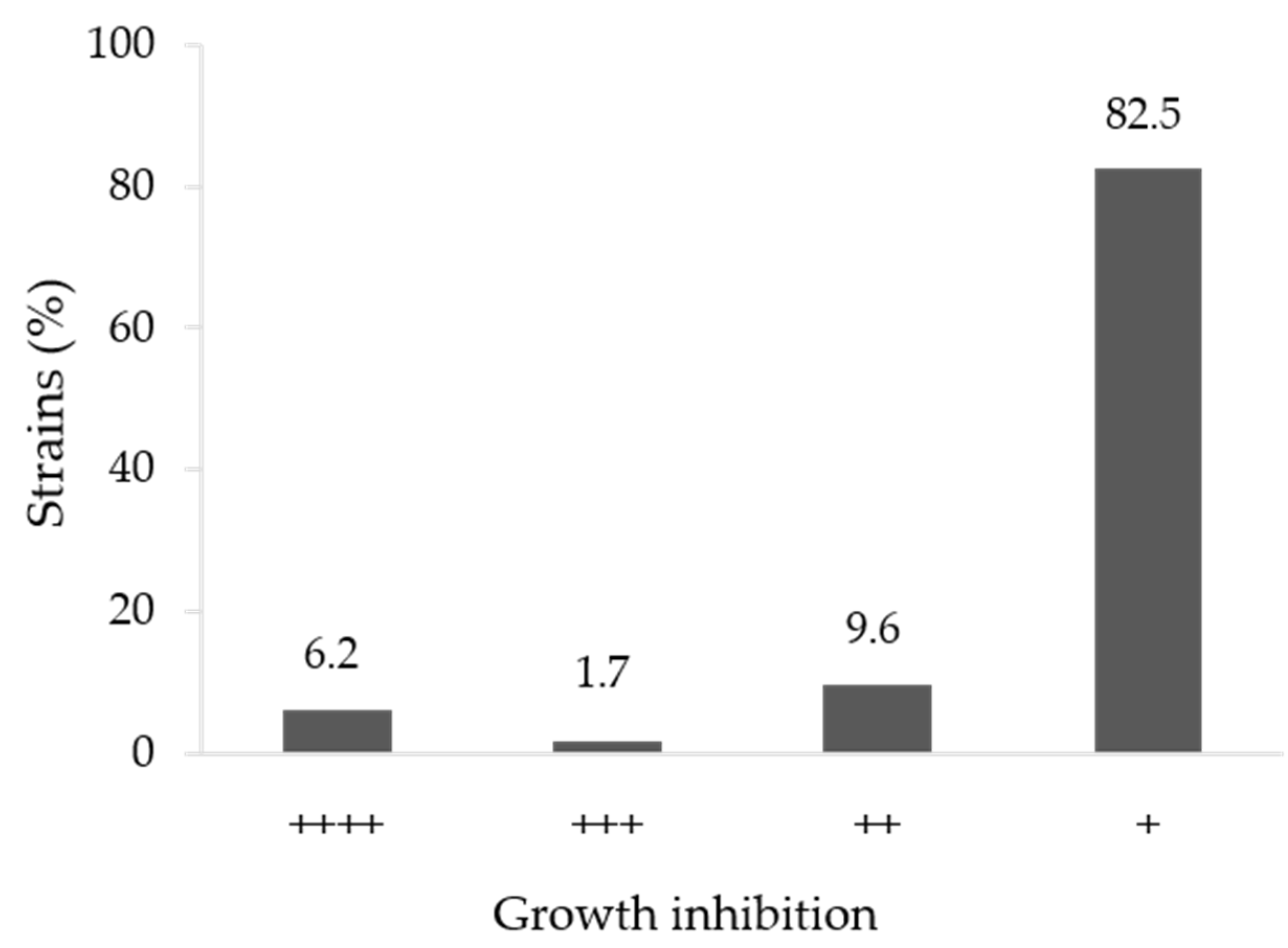
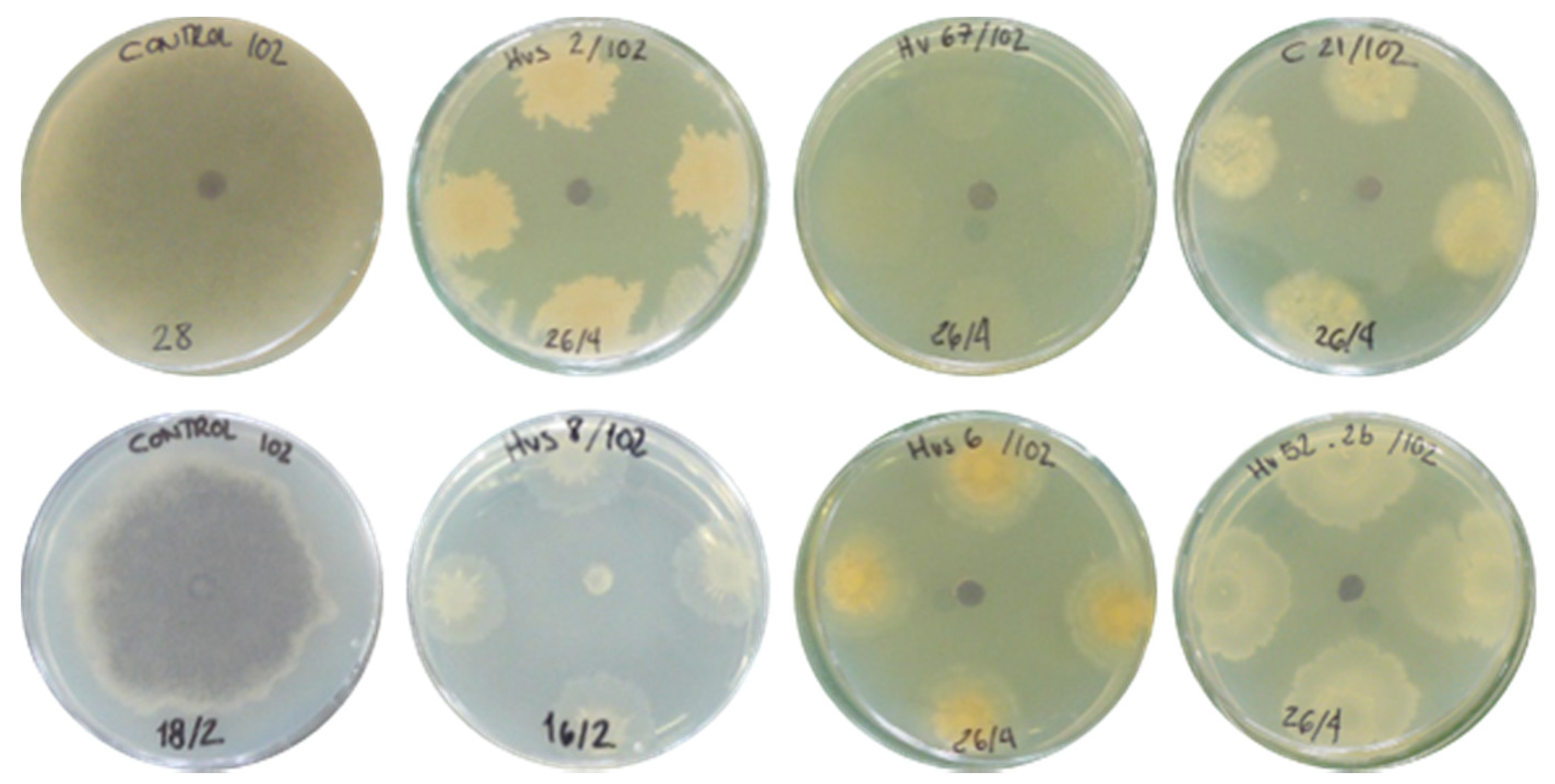
3.1. In Vitro Antifungal Activity
3.2. Molecular Characterization
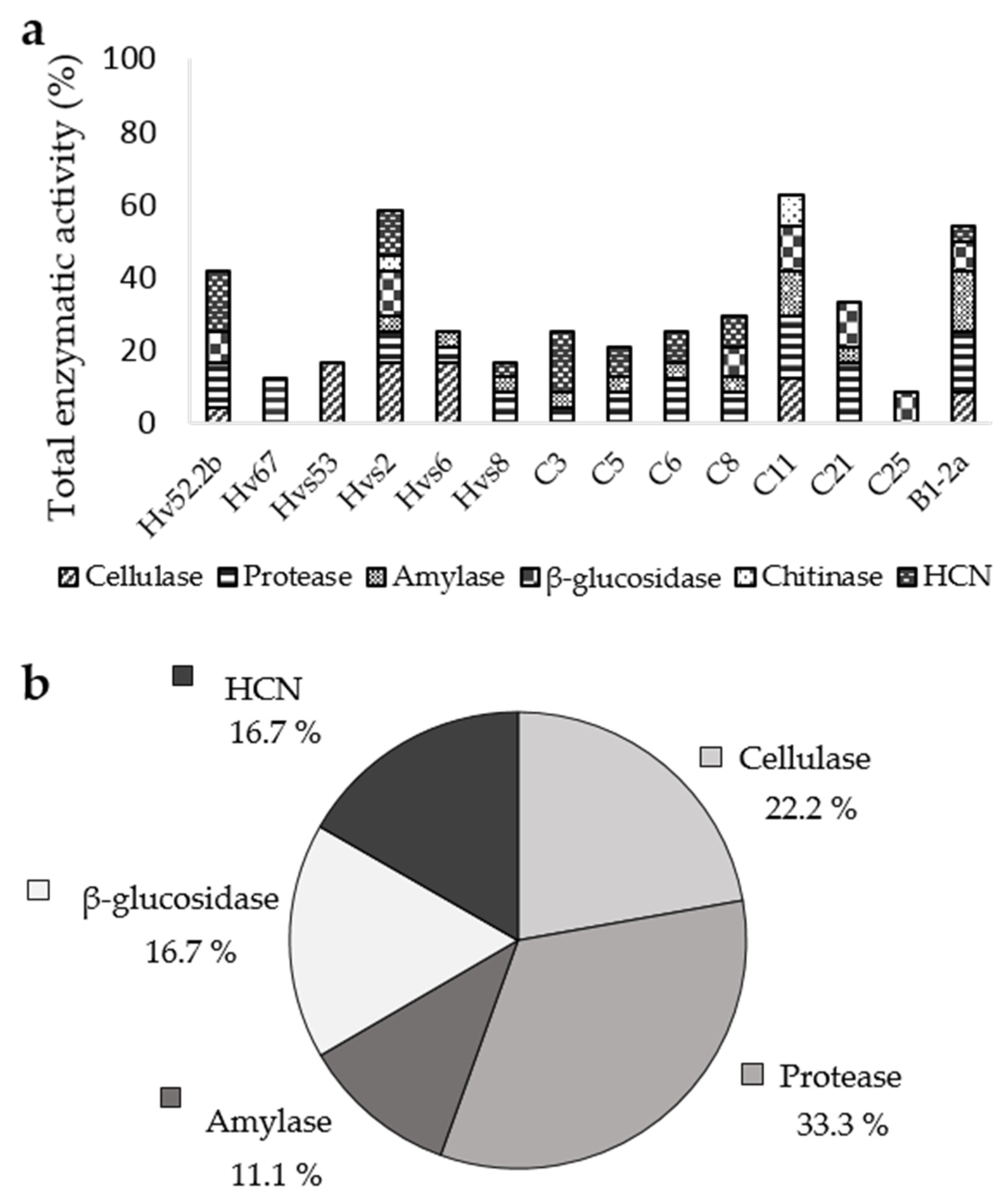
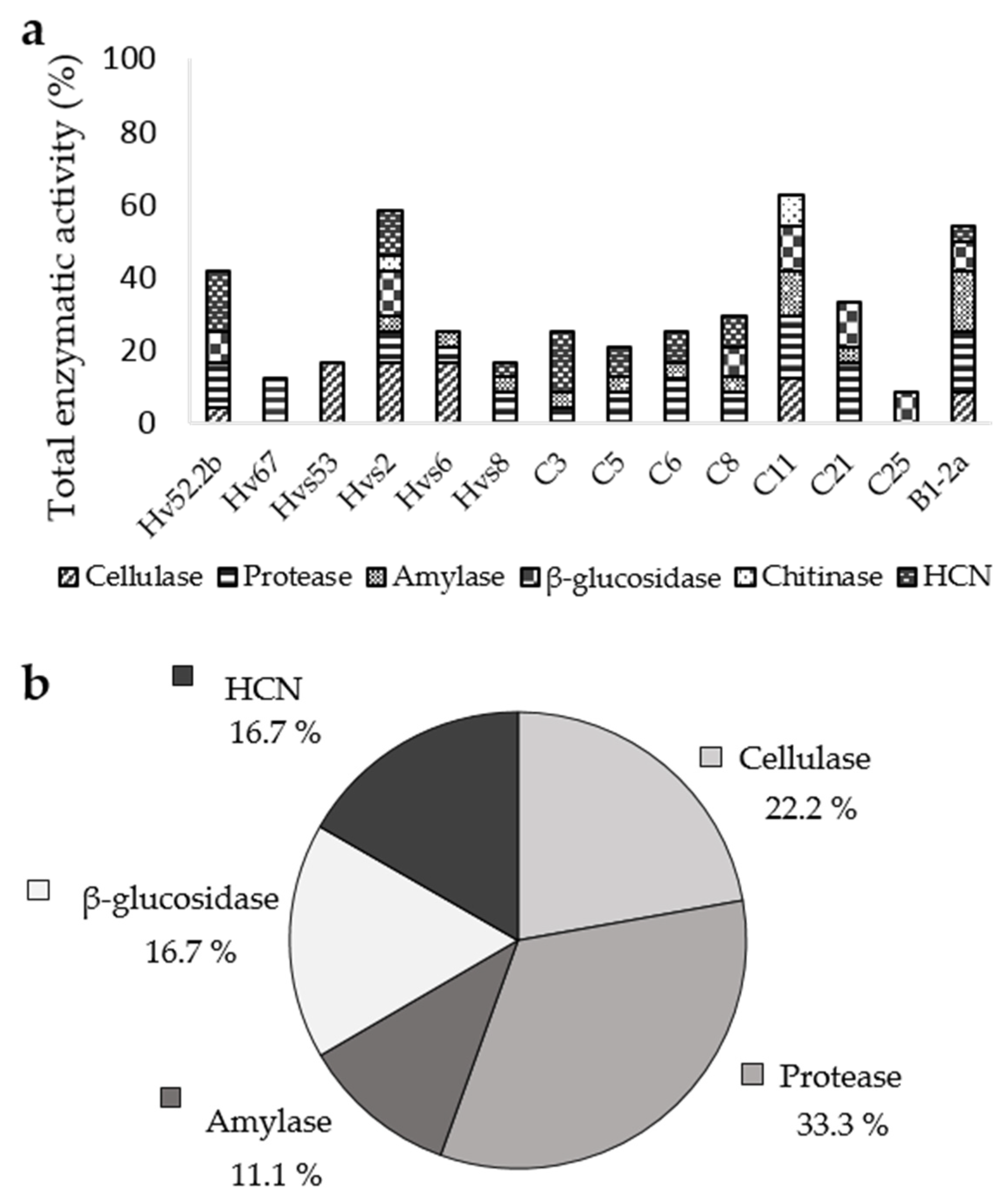
3.3. Enzymatic Activity and HCN Production
3.4. Biocontrol against Charcoal Rot Disease on Strawberry Plants
3.5. Growth Promotion in Greenhouse Trials
3.6. Field Trials
3.6.1. Fruit Weight
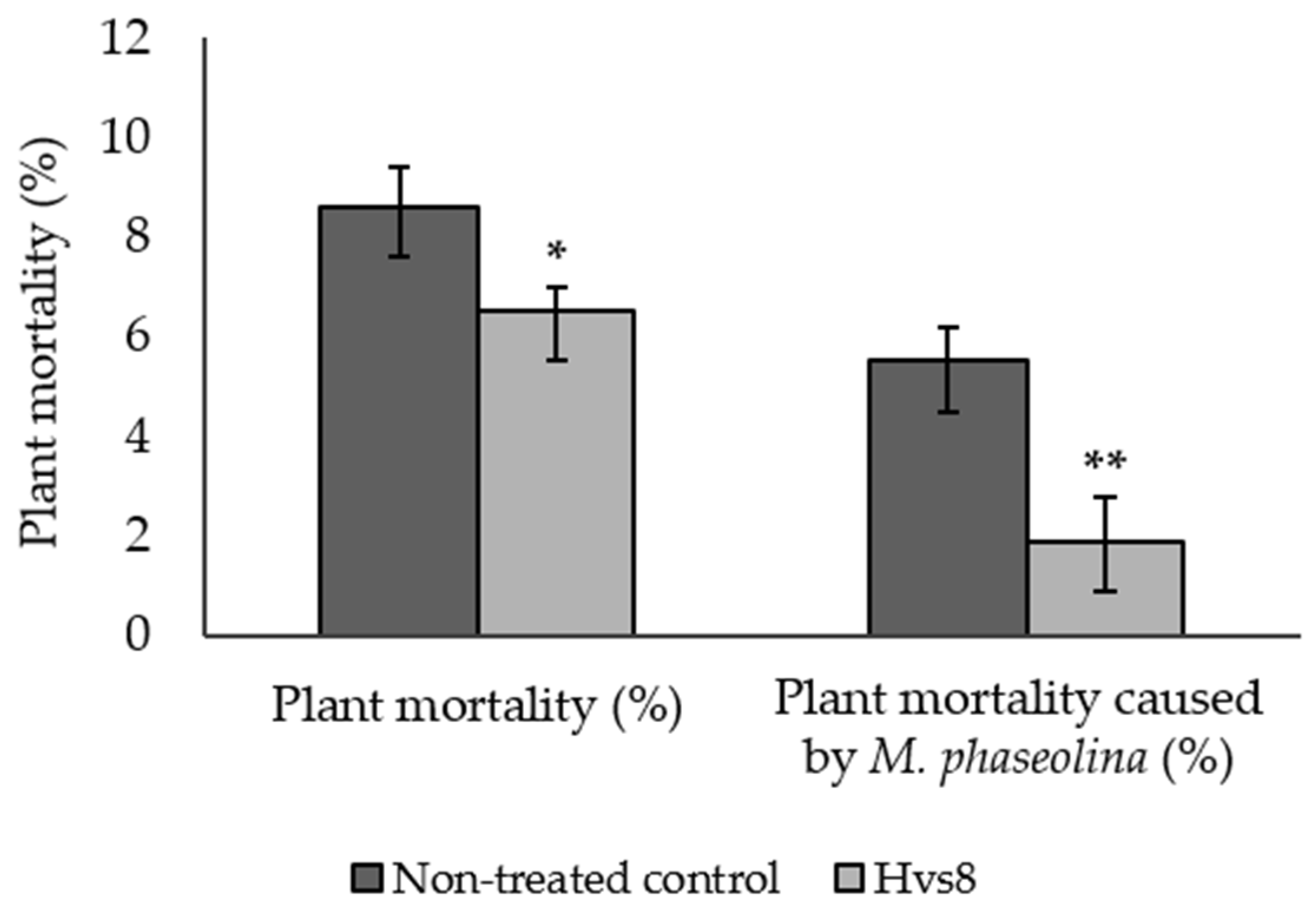
3.6.2. Plant Mortality and Charcoal Rot Incidence in Dead Plants
3.6.3. Fruit Quality
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kaur, S.; Dhillon, G.S.; Brar, S.K.; Vallad, G.E.; Chand, R.; Chauhan, V.B. Emerging phytopathogen Macrophomina phaseolina: Biology, economic importance and current diagnostic trends. Crit. Rev. Microbiol. 2012, 38, 136–151. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.T. Crown rot of strawberry caused by Macrophomina phaseolina in California. Plant Dis. 2008, 92, 1253. [Google Scholar] [CrossRef]
- Avilés, M.; Castillo, S.; Bascon, J.; Zea-Bonilla, T.; Martín-Sánchez, P.M.; Pérez-Jiménez, R.M. First report of Macrophomina phaseolina causing crown and root rot of strawberry in Spain. Plant Pathol. 2008, 57, 382. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#home (accessed on 27 August 2021).
- Talavera, M.; Gómez-Mora, J.A.; de los Santos, B.; Miranda, L.; Vela, M.D.; Fernández-Plaza, M.; Medina-Mínguez, J.J.; Soria, C.; Verdejo-Lucas, S. Problemática fitopatológica del cultivo de la fresa en Huelva. Agricultura 2018, 1022, 6–9. [Google Scholar]
- Lodha, S.; Mawar, R. Population dynamics of Macrophomina phaseolina in relation to disease management: A review. J. Phytopathol. 2019, 168, 1–17. [Google Scholar] [CrossRef]
- Baggio, J.S.; Cordova, L.G.; Peres, N.A. Sources of inoculum and survival of Macrophomina phaseolina in Florida strawberry fields. Plant Dis. 2019, 103, 2417–2424. [Google Scholar] [CrossRef]
- Reino, J.L.; Guerrero, R.F.; Hernández-Galán, R.; Collado, I.G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 2008, 7, 89–123. [Google Scholar] [CrossRef]
- O’Brien, P.A. Biological control of plant diseases. Australas. Plant Pathol. 2017, 46, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Vega, F.E. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Rengasamy, P. Soil salinity and sodicity. In Growing Crops with Reclaimed Wastewater; CSIRO Publishing: Clayton, Australia, 2006; pp. 125–138. [Google Scholar]
- Redondo-Gómez, S.; Mateos-Naranjo, E.; Figueroa, M.E.; Davy, A.J. Salt stimulation of growth and photosynthesis in an extreme halophyte, Arthrocnemum macrostachyum. Plant Biol. 2010, 12, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Chacón-López, A.; Guardado-Valdivia, L.; Bañuelos-González, M.; López-García, U.; Montalvo-González, E.; Arvizu-Gómez, J.; Stoll, A.; Aguilera, S. Effect of metabolites produced by Bacillus atrophaeus and Brevibacterium frigoritolerans strains on postharvest biocontrol of Alternaria alternata in tomato (Solanum lycopersicum L.). Biocontrol Sci. 2021, 26, 67–74. [Google Scholar] [CrossRef]
- Chopra, A.; Kumar Vandana, U.; Rahi, P.; Satpute, S.; Mazumder, P.B. Plant growth promoting potential of Brevibacterium sediminis A6 isolated from the tea rhizosphere of Assam, India. Biocatal. Agric. Biotechnol. 2020, 27, 101610. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Ebrahim, W.; Mikhailovna, P.A.; Henrich, B.; Proksch, P. Extraction and identification of some metabolites produced by antagonistic apple plant bacteria Brevibacterium halotolerans. Int. J. Adv. Res. 2015, 3, 1208–1217. [Google Scholar]
- Mohamad, O.A.A.; Li, L.; Ma, J.-B.; Hatab, S.; Xu, L.; Guo, J.-W.; Rasulov, B.A.; Liu, Y.-H.; Hedlund, B.P.; Li, W.-J. Evaluation of the antimicrobial activity of endophytic bacterial populations from Chinese traditional medicinal plant licorice and characterization of the bioactive secondary metabolites produced by Bacillus atrophaeus against Verticillium dahliae. Front. Microbiol. 2018, 9, 924. [Google Scholar] [CrossRef] [PubMed]
- Ait Bahadou, S.; Ouijja, A.; Karfach, A.; Tahiri, A.; Lahlali, R. New potential bacterial antagonists for the biocontrol of fire blight disease (Erwinia amylovora) in Morocco. Microb. Pathog. 2018, 117, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Meena, T.N.; Saharan, B.S. Plant growth promoting traits shown by bacteria Brevibacterium frigrotolerans SMA23 isolated from Aloe vera rhizosphere. Agric. Sci. Dig. 2017, 37, 226–231. [Google Scholar] [CrossRef]
- Ghosh, T.; Biswas, M.K.; Guin, C.; Roy, P. A review on characterization, therapeutic approaches and pathogenesis of Macrophomina phaseolina. Plant Cell Biotechnol. Mol. Biol. 2018, 19, 72–84. [Google Scholar]
- Pastrana, A.M.; Basallote-Ureba, M.J.; Aguado, A.; Khalid, A.K.D.I.; Capote, N. Biological control of strawberry soil-borne pathogens Macrophomina phaseolina and Fusarium solani, using Trichoderma asperellum and Bacillus spp. Phytopathol. Mediterr. 2016, 55, 109–120. [Google Scholar] [CrossRef]
- Barcia Piedras, J.M. Fitodesalinización Asistida Por Microorganismos Para la Resuperación de Suelos Agrícolas. Ph.D. Thesis, Universidad de Sevilla, Sevilla, Spain, 2020. [Google Scholar]
- De la Lastra, E.; Camacho, M.; Capote, N. Soil bacteria as potential biological control agents of Fusarium species associated to asparagus decline syndrome. Appl. Sci. 2021, 11, 8356. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; John Wiley and Sons: New York, NY, USA, 1991; p. 329. [Google Scholar]
- Camacho, M.; Medina, C.; Rodríguez-Navarro, D.N.; Temprano Vera, F. Biodiversity of rhizobia present in plant nodules of Biserrula pelecinus across Southwest Spain. Syst. Appl. Microbiol. 2019, 42, 415–421. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Teather, R.M.; Wood, P.J. Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl. Environ. Microbiol. 1982, 43, 777–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harley, J.P.; Prescott, L.M. Laboratory Exercises in Microbiology, 6th ed.; McGraw-Hill Science Engineering: New York, NY, USA, 2002. [Google Scholar]
- Cowan, S.T.; Steel, K.J. Manual for the Identification of Medical Bacteria; Cambridge University Press: Cambridge, UK, 1965. [Google Scholar]
- Gong, X.; Gruninger, R.J.; Qi, M.; Paterson, L.; Forster, R.J.; Teather, R.M.; Mcallister, T.A. Cloning and identification of novel hydrolase genes from a dairy cow rumen metagenomic library and characterization of a cellulase gene. BMC Res. Notes 2012, 5, 566. [Google Scholar] [CrossRef] [Green Version]
- Mesa, J.; Mateos-Naranjo, E.; Caviedes, M.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Endophytic cultivable bacteria of the metal bioaccumulator Spartina maritima improve plant growth but not metal uptake in polluted marshes soils. Front. Microbiol. 2015, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lorck, H. Production of hydrocyanic acid by bacteria. Physiol. Plant. 1948, 1, 142–146. [Google Scholar] [CrossRef]
- Avilés, M.; Castillo, S.; Borrero, C.; Castillo, M.L.; Zea-Bonilla, T.; Pérez-Jiménez, R.M. Response of strawberry cultivars: “Camarosa”, “Candonga” and “Ventana” to inoculation with isolates of Macrophomina phaseolina. Acta Hortic. 2009, 842, 291–294. [Google Scholar] [CrossRef]
- Fang, X.; Phillips, D.; Li, H.; Sivasithamparam, K.; Barbetti, M.J. Comparisons of virulence of pathogens associated with crown and root diseases of strawberry in Western Australia with special reference to the effect of temperature. Sci. Hortic. 2011, 131, 39–48. [Google Scholar] [CrossRef]
- López-Aranda, J.M. The cultivation of the strawberry in Huelva. In The Strawberry Crop at Huelva.; de Andalucía, J., Ed.; Ideas Exclusivas y Publicidad S.L.: Sevilla, Spain, 2008; pp. 101–174. [Google Scholar]
- Zveibil, A.; Freeman, S. Methods for detection of soilborne pathogens affecting strawberry in Israel. Acta Hortic. 2009, 842, 191–194. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and measurement of anthocyanins by UV-visible spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F1–F2. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Humayun, P.; Kiran, B.K.; Kannan, I.G.K.; Vidya, M.S.; Deepthi, K.; Rupela, O. Evaluation of bacteria isolated from rice rhizosphere for biological control of charcoal rot of sorghum caused by Macrophomina phaseolina (Tassi) Goid. World J. Microbiol. Biotechnol. 2011, 27, 1313–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castaldi, S.; Petrillo, C.; Donadio, G.; Piaz, F.D.; Cimmino, A.; Masi, M.; Evidente, A.; Isticato, R. Plant growth promotion function of Bacillus sp. strains isolated from salt-pan rhizosphere and their biocontrol potential against Macrophomina phaseolina. Int. J. Mol. Sci. 2021, 22, 3324. [Google Scholar] [CrossRef]
- Kurniawan, O.; Wilson, K.; Mohamed, R.; Avis, T.J. Bacillus and Pseudomonas spp. provide antifungal activity against gray mold and Alternaria rot on blueberry fruit. Biol. Control 2018, 126, 136–141. [Google Scholar] [CrossRef]
- Mavrodi, D.V.; Yang, M.; Mavrodi, O.V.; Wen, S. Management of soilborne plant pathogens with beneficial root-colonizing Pseudomonas. In Advances in PGPR Research; Singh, H.B., Sarma, B.K., Keswani, C., Eds.; CABI: Boston, MA, USA, 2017; ISBN 978-1-78639-032-5. [Google Scholar]
- Motta, A.S.; Brandelli, A. Characterization of an antibacterial peptide produced by Brevibacterium linens. J. Appl. Microbiol. 2002, 92, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, J.R. Bacterial inhibition of fungal growth and pathogenicity. Microb. Ecol. Health Dis. 1999, 11, 129–142. [Google Scholar] [CrossRef]
- Abed, H.; Rouag, N.; Mouatassem, D.; Rouabhi, A. Screening for Pseudomonas and Bacillus antagonistic rhizobacteria strains for the biocontrol of Fusarium wilt of chickpea. Eurasian J. Soil Sci. 2016, 5, 182. [Google Scholar] [CrossRef] [Green Version]
- Karimi, E.; Sadeghi, A.; Peyman, A.D.; Dalvand, Y.; Omidvar, M.; Nezhad, M.K. Biocontrol science and technology biocontrol activity of salt tolerant Streptomyces isolates against phytopathogens causing root rot of sugar beet. Biocontrol Sci. Technol. 2012, 22, 333–349. [Google Scholar] [CrossRef]
- Singh, N.; Pandey, P.; Dubey, R.C.; Maheshwari, D.K. Biological control of root rot fungus Macrophomina phaseolina and growth enhancement of Pinus roxburghii (Sarg.) by rhizosphere competent Bacillus subtilis BN1. World J. Microbiol. Biotechnol. 2008, 24, 1669. [Google Scholar] [CrossRef]
- Ehteshamul-Haque, S.; Sultana, V.; Ara, J.; Athar, M. Cultivar response against root-infecting fungi and efficacy of Pseudomonas aeruginosa in controlling soybean root rot. Plant Biosyst. 2007, 141, 51–55. [Google Scholar] [CrossRef]
- Yasmin, H.; Naz, R.; Nosheen, A.; Hassan, M.N.; Ilyas, N.; Sajjad, M.; Anjum, S.; Gao, X.; Geng, Z. Identification of new biocontrol agent against charcoal rot disease caused by Macrophomina phaseolina in soybean (Glycine max L.). Sustainability 2020, 12, 6856. [Google Scholar] [CrossRef]
- Ghorchiani, M.; Etesami, H.; Alikhani, H.A. Improvement of growth and yield of maize under water stress by co-inoculating an arbuscular mycorrhizal fungus and a plant growth promoting rhizobacterium together with phosphate fertilizers. Agric. Ecosyst. Environ. 2018, 258, 59–70. [Google Scholar] [CrossRef]
- Padmavathi, T.; Dikshit, R.; Seshagiri, S. Effect of Rhizophagus spp. and plant growth-promoting Acinetobacter junii on Solanum lycopersicum and Capsicum annuum. Brazilian J. Bot. 2015, 38, 273–280. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Kim, K.; Subramanian, P.; Senthilkumar, M.; Anandham, R.; Sa, T. Arbuscular mycorrhizal fungi and associated bacteria isolated from salt-affected soil enhances the tolerance of maize to salinity in coastal reclamation soil. Agric. Ecosyst. Environ. 2016, 231, 233–239. [Google Scholar] [CrossRef]
- Singh, S.; Tripathi, A.; Maji, D.; Awasthi, A.; Vajpayee, P.; Kalra, A. Evaluating the potential of combined inoculation of Trichoderma harzianum and Brevibacterium halotolerans for increased growth and oil yield in Mentha arvensis under greenhouse and field conditions. Ind. Crop. Prod. 2019, 131, 173–181. [Google Scholar] [CrossRef]
- On, A.; Wong, F.; Ko, Q.; Tweddell, R.J.; Antoun, H.; Avis, T.J. Antifungal effects of compost tea microorganisms on tomato pathogens. Biol. Control 2015, 80, 63–69. [Google Scholar] [CrossRef]
- Lee, S.M.; Kong, H.G.; Song, G.C.; Ryu, C.M. Disruption of Firmicutes and Actinobacteria abundance in tomato rhizosphere causes the incidence of bacterial wilt disease. ISME J. 2021, 15, 330–347. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Species | Similarity (%) | Growth Inhibition |
|---|---|---|---|
| Hv52.2b | Bacillus altitudinis 41KF2bT | 99.8 | ++++ |
| Hv67 | Pseudomonas rhodesiae CIP 104664T | 99.5 | +++ |
| Hvs53 | Pseudomonas zhaodongensis NEAU-ST5-21T | 98.9 | +++ |
| Hvs2 * | Bacillus paralicheniformis KJ-16 T | 100 | ++++ |
| Hvs6 * | Streptomices fradiae DSM 40063T | 100 | ++++ |
| Hvs8 * | Brevibacterium frigoritolerans DSM 8801T | 99.9 | +++ |
| C3 | Brevibacterium frigoritolerans DSM 8801T | 100 | ++++ |
| C5 | Rossellomorea aquimaris TF-12T | 99.3 | ++++ |
| C6 | Rossellomorea aquimaris TF-12T | 99.3 | ++++ |
| C8 | Priestia aryabhattai B8W22T | 100 | ++++ |
| C11 | Bacillus mobilis 0711P9-1T | 99.8 | ++++ |
| C21 | Bacillus aryabhattai B8W22T | 99.8 | ++++ |
| C25 | Staphylococcus hominis subsp. novobiosepticus GTC 1228T | 99.7 | ++++ |
| B1-2a | Bacillus altitudinis 41KF2bT | 100 | ++++ |
| Treatments | Disease Incidence (%) | DSC |
|---|---|---|
| Non-treated control | 100 a | 2.6 |
| B1-2a | 100 a | 3.8 |
| C3 | 80 ab | 1.8 |
| C5 | 100 ab | 2.2 |
| C6 | 80 ab | 1.6 |
| C8 | 100 a | 3.0 |
| C11 | 100 a | 3.6 |
| C21 | 80 ab | 2.4 |
| C25 | 40 bc | 1.8 |
| Hvs2 | 40 bc | 1.6 |
| Hvs6 | 80 ab | 2.8 |
| Hv67 | 80 ab | 2.8 |
| Hvs53 | 80 ab | 3.6 |
| Hv52-2b | 60 abc | 2.2 |
| Hvs8 | 20 c | 0.4 |
| Significance | * | ns |
| Treatments | Number of Fruits | Weight of Fruits (g) | Shoot Dry Weight (g/Plant) | Root Dry Weight (g/Plant) |
|---|---|---|---|---|
| Non-treated control | 6.6 | 65.6 | 5.3 | 5.8 |
| Hvs8 | 6.9 | 66.3 | 5.6 | 7.8 |
| Significance | ns | ns | ns | ** |
| Treatments | Marketable Fruit Weight (g/Plant) | Second-Class Fruits Weight (g/Plant) |
|---|---|---|
| Non-treated control | 1215 | 129 |
| Hvs8 | 1190 | 119 |
| Significance | ns | ns |
| Treatments | Fruit Rots (%) a | Fruit Firmness (g of Pressure) b | Soluble Solid (Brix Degree) b | Anthocyanins (mg/100 g Fresh Weight) a | Vitamin C (mg/100 g Fresh Weight) a |
|---|---|---|---|---|---|
| Non-treated control | 23.5 | 596.8 | 7.9 | 14.2 | 48.7 |
| Hvs8 | 19.9 | 605.1 | 8.2 | 14.2 | 49.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viejobueno, J.; Rodríguez-Berbel, N.; Miranda, L.; de los Santos, B.; Camacho, M. Potential Bacterial Antagonists for the Control of Charcoal Rot (Macrophomina phaseolina) in Strawberry. Horticulturae 2021, 7, 457. https://doi.org/10.3390/horticulturae7110457
Viejobueno J, Rodríguez-Berbel N, Miranda L, de los Santos B, Camacho M. Potential Bacterial Antagonists for the Control of Charcoal Rot (Macrophomina phaseolina) in Strawberry. Horticulturae. 2021; 7(11):457. https://doi.org/10.3390/horticulturae7110457
Chicago/Turabian StyleViejobueno, Josefina, Natalia Rodríguez-Berbel, Luis Miranda, Berta de los Santos, and María Camacho. 2021. "Potential Bacterial Antagonists for the Control of Charcoal Rot (Macrophomina phaseolina) in Strawberry" Horticulturae 7, no. 11: 457. https://doi.org/10.3390/horticulturae7110457
APA StyleViejobueno, J., Rodríguez-Berbel, N., Miranda, L., de los Santos, B., & Camacho, M. (2021). Potential Bacterial Antagonists for the Control of Charcoal Rot (Macrophomina phaseolina) in Strawberry. Horticulturae, 7(11), 457. https://doi.org/10.3390/horticulturae7110457