
Alleviation of Nematode-Mediated Apple Replant Disease by Pre-Cultivation of Tagetes

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pot Experiment on the Effect of Tagetes with or without Incorporation into Soil on Nematode-Mediated ARD
2.2. Effect of Tagetes in Mitigating ARD in Apple Growing Farms and Tree Nurseries
2.3. Biotest on ARD-Induction by Nematodes from Apple Plots with Preceding Tagetes or Grass Cultivation
2.4. Statistical Analysis
3. Results
3.1. Effect of T. patula or T. tenuifolia, with or without Incorporation into Soil, on Nematode-Mediated ARD (Pot Experiment)
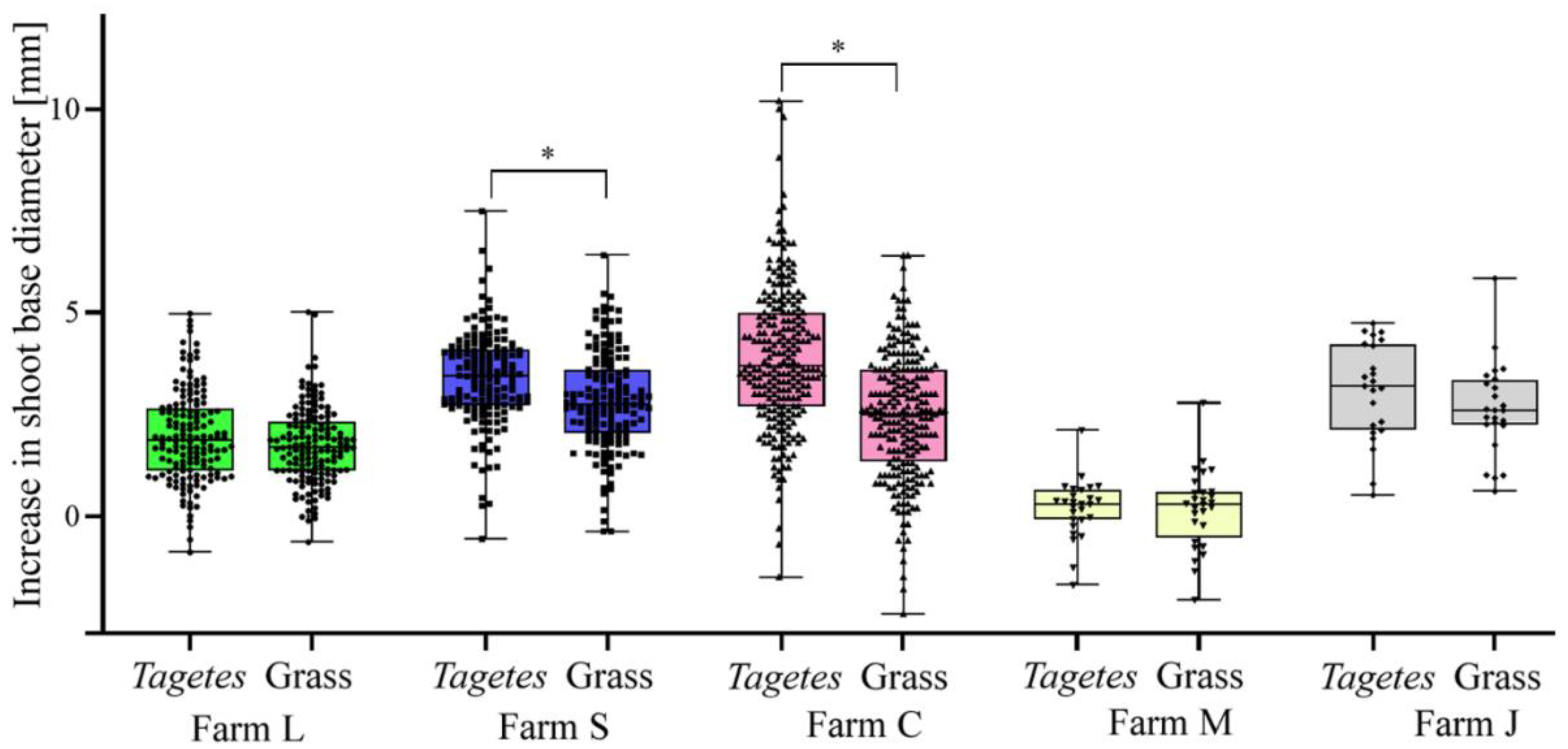
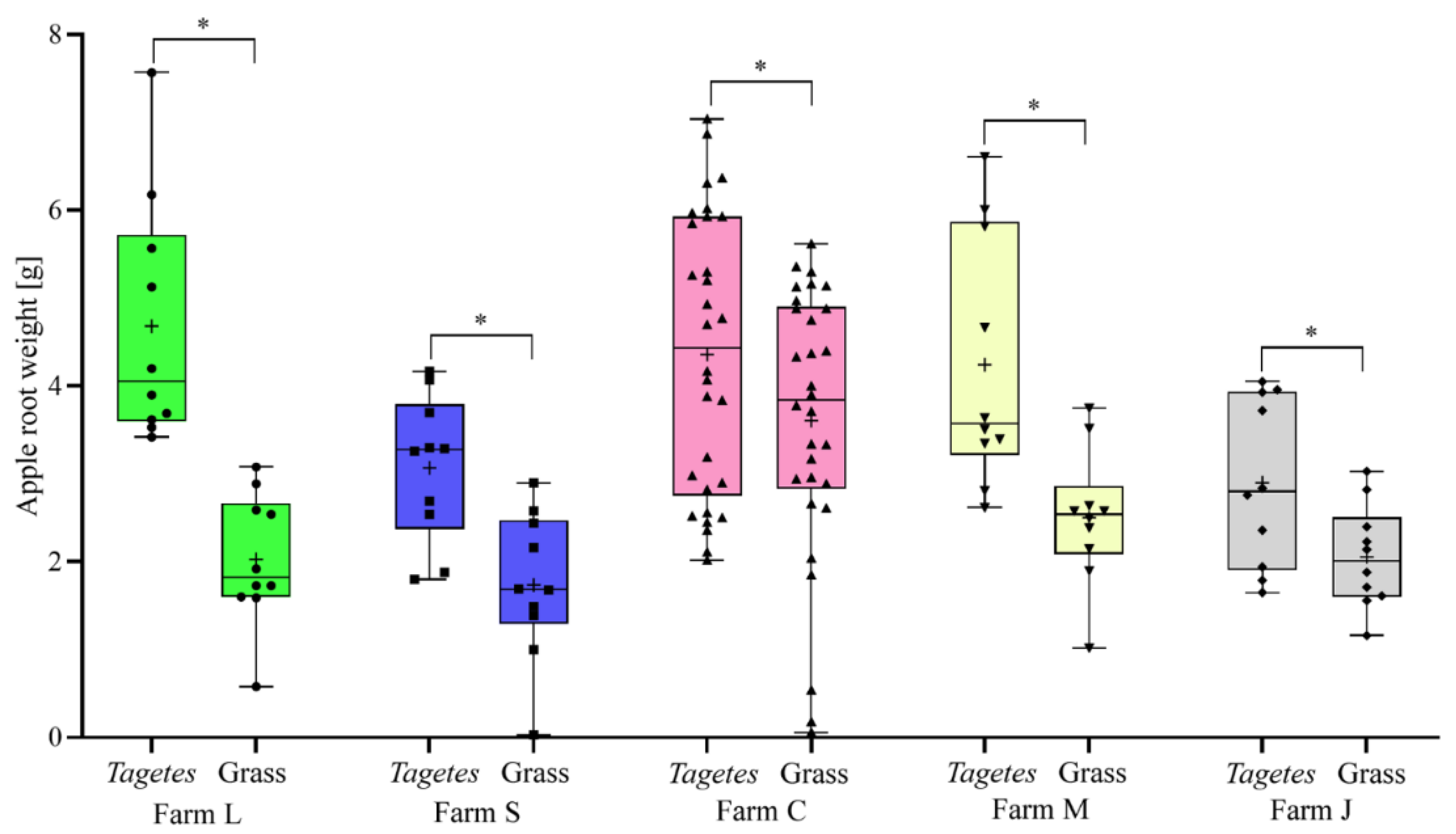
3.2. Effect of Preceding Tagetes Cultivation on Apple Shoot Growth in Apple Orchards and Tree Nurseries
3.3. Effect of Preceding Tagetes Cultivation on Plant-Parasitic Genera and Total Nematodes in Apple Orchards and Tree Nurseries
3.4. Biotest with Inoculation of Nematodes from Apple Plots with Preceding Tagetes or Grass Cultivation
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mai, W.F.; Merwin, I.A.; Abawi, G.S. Diagnosis, etiology and management of replant disorders in New York cherry and apple orchards. Acta Hortic. 1994, 363, 33–42. [Google Scholar] [CrossRef]
- Mai, W.F.; Abawi, G.S. Controlling replant diseases of pome and stone fruits in Northeastern United States by preplant fumigation. Plant Dis. 1981, 65, 859–864. [Google Scholar] [CrossRef]
- Winkelmann, T.; Smalla, K.; Amelung, W.; Baab, G.; Grunewaldt-Stöcker, G.; Kanfra, X.; Meyhöfer, R.; Reim, S.; Schmitz, M.; Vetterlein, D.; et al. Apple replant disease: Causes and mitigation strategies. Curr. Issues Mol. Biol. 2019, 30, 89–106. [Google Scholar] [CrossRef] [Green Version]
- Mazzola, M.; Manici, L.M. Apple replant disease: Role of microbial ecology in cause and control. Annu. Rev. Phytopathol. 2012, 50, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Grunewaldt-Stöcker, G.; Mahnkopp, F.; Popp, C.; Maiss, E.; Winkelmann, T. Diagnosis of apple replant disease (ARD): Microscopic evidence of early symptoms in fine roots of different apple rootstock genotypes. Sci. Hortic. 2019, 243, 583–594. [Google Scholar] [CrossRef]
- Mazzola, M. Elucidation of the microbial complex having a causal role in the development of apple replant disease in Washington. Phytopathology 1998, 88, 930–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, B.; Smalla, K.; Winkelmann, T. Evaluation of apple replant problems based on different soil disinfection treatments—Links to soil microbial community structure? Plant Soil 2013, 366, 617–631. [Google Scholar] [CrossRef]
- Klaus, H. Das Problem der Bodenmüdigkeit unter Berücksichtigung des Obstbaus. Landw. Jahrb. 1939, 89, 413–459. [Google Scholar]
- Von Bronsart, H. Der heutige Stand unseres Wissens von der Bodenmüdigkeit. J. Plant Nutr. Soil Sci. 1949, 45, 166–193. [Google Scholar] [CrossRef]
- Manici, L.M.; Caputo, F.; Saccà, M.L. Secondary metabolites released into the rhizosphere by Fusarium oxysporum and Fusarium spp. as underestimated component of nonspecific replant disease. Plant Soil 2017, 415, 85–98. [Google Scholar] [CrossRef]
- Otto, G. Investigations on the cause of soil sickness in fruit trees VII. An actinomycete isolated from rootlets of apple seedlings, the probable cause of specific apple replant disease. J. Cultiv. Plants 2017, 69, 175–179. [Google Scholar] [CrossRef]
- Tewoldemedhin, Y.T.; Mazzola, M.; Labuschagne, I.; McLeod, A. A multi-phasic approach reveals that apple replant disease is caused by multiple biological agents, with some agents acting synergistically. Soil Biol. Biochem. 2011, 43, 1917–1927. [Google Scholar] [CrossRef]
- Tewoldemedhin, Y.T.; Mazzola, M.; Botha, W.J.; Spies, C.F.J.; McLeod, A. Characterization of fungi (Fusarium and Rhizoctonia) and oomycetes (Phytophthora and Pythium) associated with apple orchards in South Africa. Eur. J. Plant Pathol. 2011, 130, 215–229. [Google Scholar] [CrossRef]
- Nyoni, M.; Mazzola, M.; Wessels, J.P.B.; McLeod, A. The efficacy of semiselective chemicals and chloropicrin/1,3-dichloropropene-containing fumigants in managing apple replant disease in South Africa. Plant Dis. 2019, 103, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Popp, C.; Wamhoff, D.; Winkelmann, T.; Maiss, E.; Grunewaldt-Stöcker, G. Molecular identification of Nectriaceae in infections of apple replant disease affected roots collected by Harris Uni-Core punching or laser microdissection. J. Plant Dis. Prot. 2020, 127, 571–582. [Google Scholar] [CrossRef]
- Manici, L.M.; Kelderer, M.; Caputo, F.; Saccà, M.L.; Nicoletti, F.; Topp, A.R.; Mazzola, M. Involvement of Dactylonectria and Ilyonectria spp. in tree decline affecting multi-generation apple orchards. Plant Soil 2018, 425, 217–230. [Google Scholar] [CrossRef]
- Tewoldemedhin, Y.T.; Mazzola, M.; Mostert, L.; McLeod, A. Cylindrocarpon species associated with apple tree roots in South Africa and their quantification using real-time PCR. Eur. J. Plant Pathol. 2011, 129, 637–651. [Google Scholar] [CrossRef]
- Lucas, M.; Balbín-Suárez, A.; Smalla, K.; Vetterlein, D. Root growth, function and rhizosphere microbiome analyses show local rather than systemic effects in apple plant response to replant disease soil. PLoS ONE 2018, 13, e0204922. [Google Scholar] [CrossRef] [Green Version]
- Weiß, S.; Liu, B.; Reckwell, D.; Beerhues, L.; Winkelmann, T. Impaired defense reactions in apple replant disease-affected roots of Malus domestica ‘M26’. Tree Physiol. 2017, 37, 1–14. [Google Scholar] [CrossRef]
- Reim, S.; Rohr, A.-D.; Winkelmann, T.; Weiß, S.; Liu, B.; Beerhues, L.; Schmitz, M.; Hanke, M.-V.; Flachowsky, H. Genes involved in stress response and especially in phytoalexin biosynthesis are upregulated in four Malus genotypes in response to apple replant disease. Front. Plant Sci. 2020, 10, 1724. [Google Scholar] [CrossRef]
- Weiß, S.; Bartsch, M.; Winkelmann, T. Transcriptomic analysis of molecular responses in Malus domestica ‘M26’ roots affected by apple replant disease. Plant Mol. Biol. 2017, 94, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Yeates, G.W.; Bongers, T.; de Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Elling, A.A. Major emerging problems with minor Meloidogyne species. Phytopathology 2013, 103, 1092–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef]
- Singh, S.; Singh, B.; Singh, A.P. Nematodes: A threat to sustainability of agriculture. Procedia Environ. Sci. 2015, 29, 215–216. [Google Scholar] [CrossRef] [Green Version]
- Stirling, G.R. Biological Control of Plant Parasitic Nematodes, 2nd ed.; CABI: Wallingford, UK, 2014; ISBN 1780644159. [Google Scholar]
- Back, M.A.; Haydock, P.P.J.; Jenkinson, P. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathol. 2002, 51, 683–697. [Google Scholar] [CrossRef]
- Morris, K.A.; Langston, D.B.; Dutta, B.; Davis, R.F.; Timper, P.; Noe, J.P.; Dickson, D.W. Evidence for a disease complex between Pythium aphanidermatum and root-knot nematodes in cucumber. Plant Health Progr. 2016, 17, 200–201. [Google Scholar] [CrossRef]
- Dullahide, S.R.; Stirling, G.R.; Nikulin, A.; Stirling, A.M. The role of nematodes, fungi, bacteria, and abiotic factors in the etiology of apple replant problems in the Granite Belt of Queensland. Aust. J. Exp. Agric. 1994, 34, 1177–1182. [Google Scholar] [CrossRef]
- Jaffee, B.A.; Abawi, G.S.; Mai, W.F. Role of soil microflora and Pratylenchus penetrans in an apple replant disease. Phytopathology 1982, 72, 247–251. [Google Scholar] [CrossRef]
- Kanfra, X.; Liu, B.; Beerhues, L.; Sørensen, S.J.; Heuer, H. Free-living nematodes together with associated microbes play an essential role in apple replant disease. Front. Plant Sci. 2018, 9, 1666. [Google Scholar] [CrossRef]
- Covey, R.P., Jr.; Benson, N.R.; Haglund, W.A. Effect of soil fumigation on the apple replant disease in Washington. Phytopathology 1979, 69, 684–686. [Google Scholar] [CrossRef]
- Auvil, T.D.; Schmidt, T.R.; Hanrahan, I.; Castillo, F.; McFerson, J.R.; Fazio, G. Evaluation of dwarfing rootstocks in washington apple replant sites. Acta Hortic. 2011, 265–271. [Google Scholar] [CrossRef]
- Onkendi, E.M.; Kariuki, G.M.; Marais, M.; Moleleki, L.N. The threat of root-knot nematodes (Meloidogyne spp.) in Africa: A review. Plant Pathol. 2014, 63, 727–737. [Google Scholar] [CrossRef] [Green Version]
- Zasada, I.A.; Halbrendt, J.M.; Kokalis-Burelle, N.; LaMondia, J.; McKenry, M.V.; Noling, J.W. Managing nematodes without methyl bromide. Annu. Rev. Phytopathol. 2010, 48, 311–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzola, M.; Hewavitharana, S.S.; Strauss, S.L. Brassica seed meal soil amendments transform the rhizosphere microbiome and improve apple production through resistance to pathogen reinfestation. Phytopathology 2015, 105, 460–469. [Google Scholar] [CrossRef] [Green Version]
- Yim, B.; Hanschen, F.S.; Wrede, A.; Nitt, H.; Schreiner, M.; Smalla, K.; Winkelmann, T. Effects of biofumigation using Brassica juncea and Raphanus sativus in comparison to disinfection using Basamid on apple plant growth and soil microbial communities at three field sites with replant disease. Plant Soil 2016, 406, 389–408. [Google Scholar] [CrossRef]
- Agbenin, N.O. Biological control of plant parasitic nematodes: Prospects and challenges for the poor Africa farmer. Plant Protect. Sci. 2012, 47, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Fourie, H.; van Aardt, W.J.; Venter, C.; Tiedt, L.R. The effects of CropGuard® on the motility, ultrastructure, and respiration of two Meloidogyne species. Nematropica 2014, 44, 85–92. [Google Scholar]
- Reim, S.; Cestaro, A.; Siewert, C.; Wöhner, T.; Mahnkopp-Dirks, F.; Winkelmann, T.; Hanke, M.-V.; Flachowsky, H. Evaluation of tolerance to apple replant disease (ARD) in Malus germplasm. Acta Hortic. 2021, 327–334. [Google Scholar] [CrossRef]
- Karakas, M.; Bolukbasi, E. A review: Using marigolds (Tagetes spp.) as an alternative to chemical nematicides for nematode management. Int. J. Adv. Eng. Manag. Sci. 2019, 5, 556–560. [Google Scholar] [CrossRef]
- Xu, L.; Chen, J.; Qi, H.; Shi, Y. Phytochemicals and their biological activities of plants in Tagetes L. Chin. Herb. Med. 2012, 4, 103–117. [Google Scholar] [CrossRef]
- Supradip, S.; Suresh, W.; Kundu, A.; Kumar, B.; Decksha, J. Antifungal acetylinic thiophenes from Tagetes minuta: Potential biopesticide. J. Appl. Bot. Food Qual. 2012, 85, 207–211. [Google Scholar]
- Hooks, C.R.R.; Wang, K.-H.; Ploeg, A.; McSorley, R. Using marigold (Tagetes spp.) as a cover crop to protect crops from plant-parasitic nematodes. Appl. Soil Ecol. 2010, 46, 307–320. [Google Scholar] [CrossRef]
- Yim, B.; Nitt, H.; Wrede, A.; Jacquiod, S.; Sørensen, S.J.; Winkelmann, T.; Smalla, K. Effects of soil pre-treatment with Basamid® granules, Brassica juncea, Raphanus sativus, and Tagetes patula on bacterial and fungal communities at two apple replant disease sites. Front. Microbiol. 2017, 8, 1604. [Google Scholar] [CrossRef] [PubMed]
- Mahnkopp, F.; Simon, M.; Lehndorff, E.; Pätzold, S.; Wrede, A.; Winkelmann, T. Induction and diagnosis of apple replant disease (ARD): A matter of heterogeneous soil properties? Sci. Hortic. 2018, 241, 167–177. [Google Scholar] [CrossRef]
- Hooper, D.J.; Hallmann, J.; Subbotin, S.A. Methods for Extraction, Processing and Detection of Plant and Soil Nematodes. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture, 2nd ed.; Luc, M., Sikora, R.A., Bridge, J., Eds.; CABI: Wallingford, UK, 2005; pp. 53–85. ISBN 9780851997278. [Google Scholar]
- European and Mediterranean Plant Protection Organization. PM 7/119 (1) Nematode extraction. EPPO Bull. 2013, 43, 471–495. [Google Scholar] [CrossRef]
- Santo, G.S.; Wilson, J.H. Effects of fenamiphos on Pratylenchus penetrans and growth of apple. J. Nematol. 1990, 22, 779–782. [Google Scholar] [PubMed]
- Moein, S.; Mazzola, M.; Ntushelo, N.S.; McLeod, A. Apple nursery trees and irrigation water as potential external inoculum sources of apple replant disease in South Africa. Eur. J. Plant Pathol. 2019, 153, 1131–1147. [Google Scholar] [CrossRef]
- Isutsa, D.K.; Merwin, I.A. Nematodes and fungi associated with apple replant disorder in sampled New York State orchards. Glob. J. Biosci. Biotechnol. 2014, 3, 174–180. [Google Scholar]
- Hoestra, H. Replant disease of apple in the Netherlands. Meded. Landbouwhogeschool Wageningen 1968, 68, 1–105. [Google Scholar]
- Oostenbrink, M.; Hoestra, H. Nematode damage and “specific sickness” in Rosa, Malus and Laburnum. Tijdschr. Over Plantenziekten 1961, 67, 264–272. [Google Scholar]
- Wang, L.; Mazzola, M. Field evaluation of reduced rate Brassicaceae seed meal amendment and rootstock genotype on the microbiome and control of apple replant disease. Phytopathology 2019, 109, 1378–1391. [Google Scholar] [CrossRef]
- Nježić, B.; de Sutter, N.; Moens, M. Interaction of Tagetes patula cv. Single Gold with the life cycle of the plant-parasitic nematodes Meloidogyne chitwoodi and Pratylenchus penetrans. Russ. J. Nematol. 2014, 22, 101–108. [Google Scholar]
- Ploeg, A.T. Effects of selected marigold varieties on root-knot nematodes and tomato and melon yields. Plant Dis. 2002, 86, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Kimpinski, J.; Arsenault, W.J.; Gallant, C.E.; Sanderson, J.B. The effect of marigolds (Tagetes spp.) and other cover crops on Pratylenchus penetrans and on following potato crops. J. Nematol. 2000, 32, 531–536. [Google Scholar] [PubMed]
- Hamaguchi, T.; Sato, K.; Vicente, C.S.L.; Hasegawa, K. Nematicidal actions of the marigold exudate α-terthienyl: Oxidative stress-inducing compound penetrates nematode hypodermis. Biol. Open 2019, 8, bio038646. [Google Scholar] [CrossRef] [Green Version]
- Marotti, I.; Marotti, M.; Piccaglia, R.; Nastri, A.; Grandi, S.; Dinelli, G. Thiophene occurrence in different Tagetes species: Agricultural biomasses as sources of biocidal substances. J. Sci. Food Agric. 2010, 90, 1210–1217. [Google Scholar] [CrossRef]
- Uhlenbroek, J.H.; Bijloo, J.D. Investigations on nematicides: I. Isolation and structure of a nematicidal principe occurring in Tagetes roots. Recl. Trav. Chim. Pays-Bas 1958, 77, 1004–1009. [Google Scholar] [CrossRef]
- Wang, K.-H.; Hooks, C.R.; Ploeg, A. Protecting crops from nematode pests: Using marigold as an alternative to chemical nematicides. Plant Dis. 2007, PD-35, 1–6. [Google Scholar]
- Natarajan, N.; Cork, A.; Boomathi, N.; Pandi, R.; Velavan, S.; Dhakshnamoorthy, G. Cold aqueous extracts of African marigold, Tagetes erecta for control tomato root knot nematode, Meloidogyne incognita. Crop Prot. 2006, 25, 1210–1213. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Farm | Tagetes Treatment (2019) | Apple Cultivation (2020) | ||||
|---|---|---|---|---|---|---|
| Species | Sowing | Incorporation | Apple Plants | Measured Plants Per Treatment | Number of Blocks | |
| L, tree nursery | T. erecta | Jun | Sep | Rootstock A2 | 160 | 4 |
| S, tree nursery | T. patula ‘Nemamix‘ | May | Apr (2020) | Rootstock M9 | 160 | 4 |
| C, tree nursery | T. patula ‘Nemamix‘ | May | Nov | Rootstock ‘Bittenfelder’ | 235 | 6 |
| M, organic orchard | T. patula ‘Nemamix‘ | May | Nov | ‘Red Prince’ on M9 | 28 | 1 |
| J, orchard | T. patula ‘Nemamix‘ | May | Nov | ‘Sweet Tango’ on M9 | 24 | 2 |
| Farm | Location | Soil Type | Clay [%] | Silt [%] | Sand [%] | pH (CaCl2) | C [%] | N [%] |
|---|---|---|---|---|---|---|---|---|
| L | Northrhine-Westfalia 51.83254, 7.42113 | Braunerde-Podzol | 7.4 | 16.0 | 76.6 | 5.9 | 2.95 | 0.18 |
| S | Schleswig Holstein 53.67823, 9.73118 | Podzol-Parabraunerde | 5.9 | 41.9 | 52.2 | 4.7 | 1.59 | 0.11 |
| C | Schleswig Holstein 53.63304, 9.70630 | Podzol | 5.1 | 5.5 | 89.4 | 5.3 | 2.45 | 0.14 |
| M | Lower Saxony 53.50748, 9.68593 | Kleimarsch | 30.5 | 64.0 | 5.5 | 5.9 | 3.37 | 0.32 |
| J | Lower Saxony 53.47639, 9.59167 | Pseudogley-Braunerde | 7.2 | 32.6 | 60.2 | 5.5 | 2.00 | 0.16 |
| Source of Nematodes Inoculated to Apple Plantlets | Plant Growth Parameter 1 | |||
|---|---|---|---|---|
| Shoot Length (cm) | Shoot Fresh Mass (g) | Leaf Fresh Mass (g) | Root Fresh Mass (g) | |
| Untreated ARD soil | 13.7 ± 3.8c | 3.7 ± 1.4c | 2.3 ± 0.9c | 1.9 ± 0.6b |
| T. patula treated ARD soil | 18.4 ± 1.4ab | 4.9 ± 1.2a | 3.0 ± 0.9ab | 3.7 ± 0.6a |
| T. patula treated ARD soil (with incorporation) | 18.4 ± 3.5ab | 4.9 ± 0.9ab | 3.0 ± 0.6ab | 3.8 ± 0.8a |
| T. tenuifolia treated ARD soil | 11.9 ± 5.0c | 3.5 ± 0.9c | 2.0 ± 0.6c | 3.4 ± 1.1a |
| T. tenuifolia treated ARD soil (with incorporation) | 14.9 ± 3.3bc | 3.9 ± 1.3bc | 2.4 ± 0.8bc | 3.4 ± 1.1a |
| Control without nematodes | 21.2 ± 2.2a | 5.3 ± 0.5a | 3.3 ± 0.5a | 3.4 ± 1.0a |
| M26 directly grown in ARD soil | 5.8 ± 5.3d | 1.6 ± 0.3d | 0.9 ± 0.2d | 1.1 ± 0.6b |
| Tree Nursery L | Tree Nursery S | Tree Nursery C | Apple Orchard M | Apple Orchard J | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Genus/Type | G | Tag | G | Tag | G | Tag | G | Tag | G | Tag |
| Rotylenchus | 4 | 112 | 80 | |||||||
| Meloidogyne | 8 | 44 | 16 | |||||||
| Paratylenchus | 4 | 80 | 128 | 200 | ||||||
| Pratylenchus | 8 | 20 | ||||||||
| Trichodorus | 56 | 4 | 4 | |||||||
| Non-parasitic | 2744 | 3176 | 1656 | 1472 | 256 | 1104 | 384 | 594 | 1024 | 1464 |
| Plant Growth Parameter 1 | ||||
|---|---|---|---|---|
| Farm | Treatment | Shoot Length (cm) | Shoot Fresh Mass (g) | Root Fresh Mass (g) |
| L | Tagetes | 6.2 ± 1.1 a | 3.5 ± 2.9 a | 4.9 ± 1.4 a |
| Grass | 5.1 ± 0.8 b | 1.8 ± 0.5 b | 2.0 ± 0.8 b | |
| S | Tagetes | 7.1 ± 1.1 a | 3.0 ± 0.6 a | 3.1 ± 0.8 a |
| Grass | 4.7 ± 1.3 b | 1.6 ± 0.8 b | 1.7 ± 0.8 b | |
| C | Tagetes | 6.5 ± 1.4 a | 2.3 ± 0.7 a | 4.3 ± 1.5 a |
| Grass | 4.9 ± 1.1 b | 1.5 ± 0.6 b | 3.6 ± 1.5 b | |
| M | Tagetes | 7.3 ± 0.8 a | 3.1 ± 0.3 a | 4.2 ± 1.4 a |
| Grass | 5.3 ± 0.8 b | 2.1 ± 0.3 b | 2.5 ± 0.8 b | |
| J | Tagetes | 6.6 ± 0.7 a | 2.3 ± 1.0 a | 2.9 ± 1.0 a |
| Grass | 5.3 ± 0.9 b | 1.3 ± 0.6 b | 2.1 ± 0.6 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanfra, X.; Obawolu, T.; Wrede, A.; Strolka, B.; Winkelmann, T.; Hardeweg, B.; Heuer, H. Alleviation of Nematode-Mediated Apple Replant Disease by Pre-Cultivation of Tagetes. Horticulturae 2021, 7, 433. https://doi.org/10.3390/horticulturae7110433
Kanfra X, Obawolu T, Wrede A, Strolka B, Winkelmann T, Hardeweg B, Heuer H. Alleviation of Nematode-Mediated Apple Replant Disease by Pre-Cultivation of Tagetes. Horticulturae. 2021; 7(11):433. https://doi.org/10.3390/horticulturae7110433
Chicago/Turabian StyleKanfra, Xorla, Taye Obawolu, Andreas Wrede, Bernhard Strolka, Traud Winkelmann, Bernd Hardeweg, and Holger Heuer. 2021. "Alleviation of Nematode-Mediated Apple Replant Disease by Pre-Cultivation of Tagetes" Horticulturae 7, no. 11: 433. https://doi.org/10.3390/horticulturae7110433
APA StyleKanfra, X., Obawolu, T., Wrede, A., Strolka, B., Winkelmann, T., Hardeweg, B., & Heuer, H. (2021). Alleviation of Nematode-Mediated Apple Replant Disease by Pre-Cultivation of Tagetes. Horticulturae, 7(11), 433. https://doi.org/10.3390/horticulturae7110433