Improvement of Growth and Morphology of Vegetable Seedlings with Supplemental Far-Red Enriched LED Lights in a Plant Factory




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation Conditions
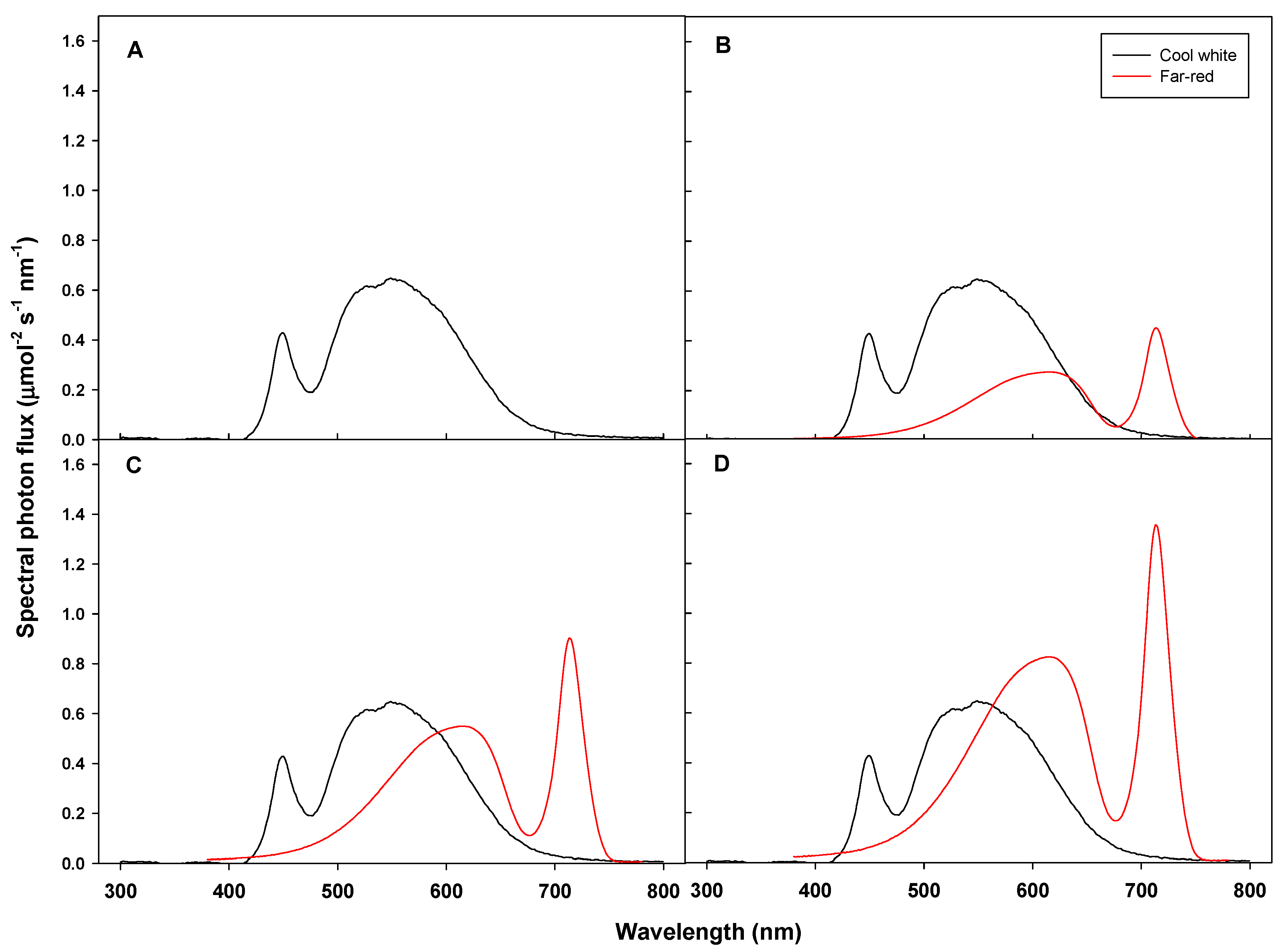
2.2. Supplemental Far-Red Lighting Treatments
2.3. Plant Growth Analysis
2.4. Statistical Analysis
3. Results
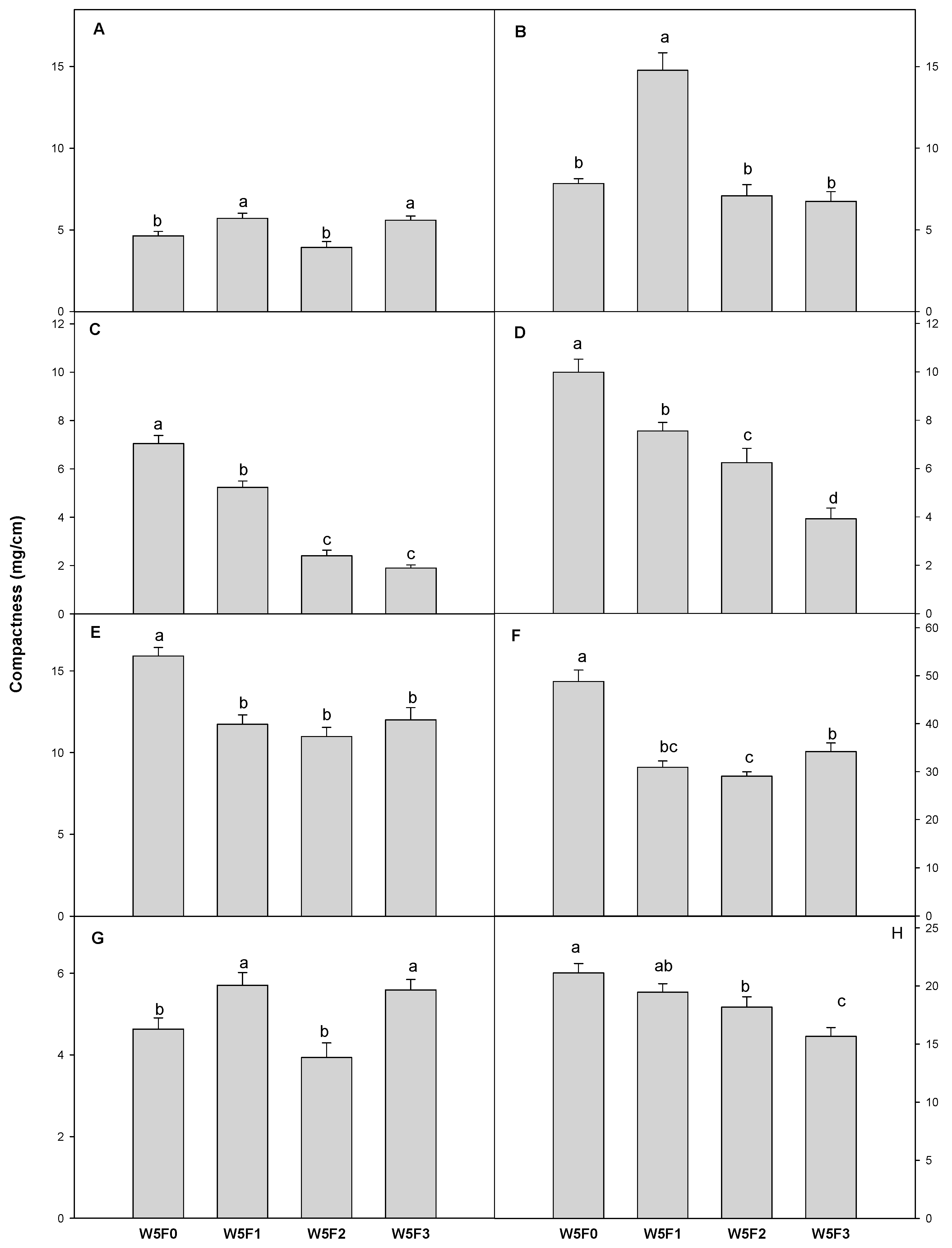
3.1. Plant Growth
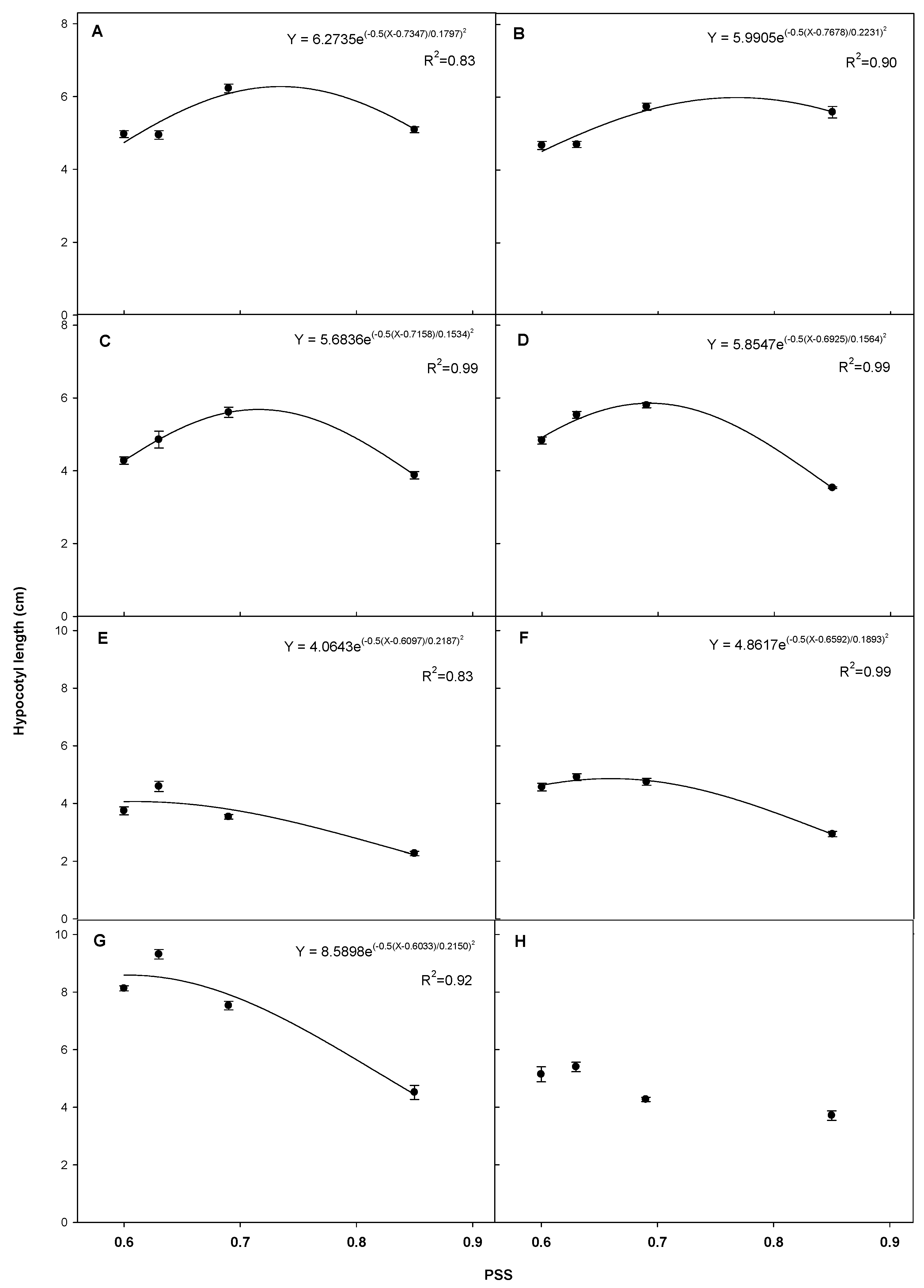
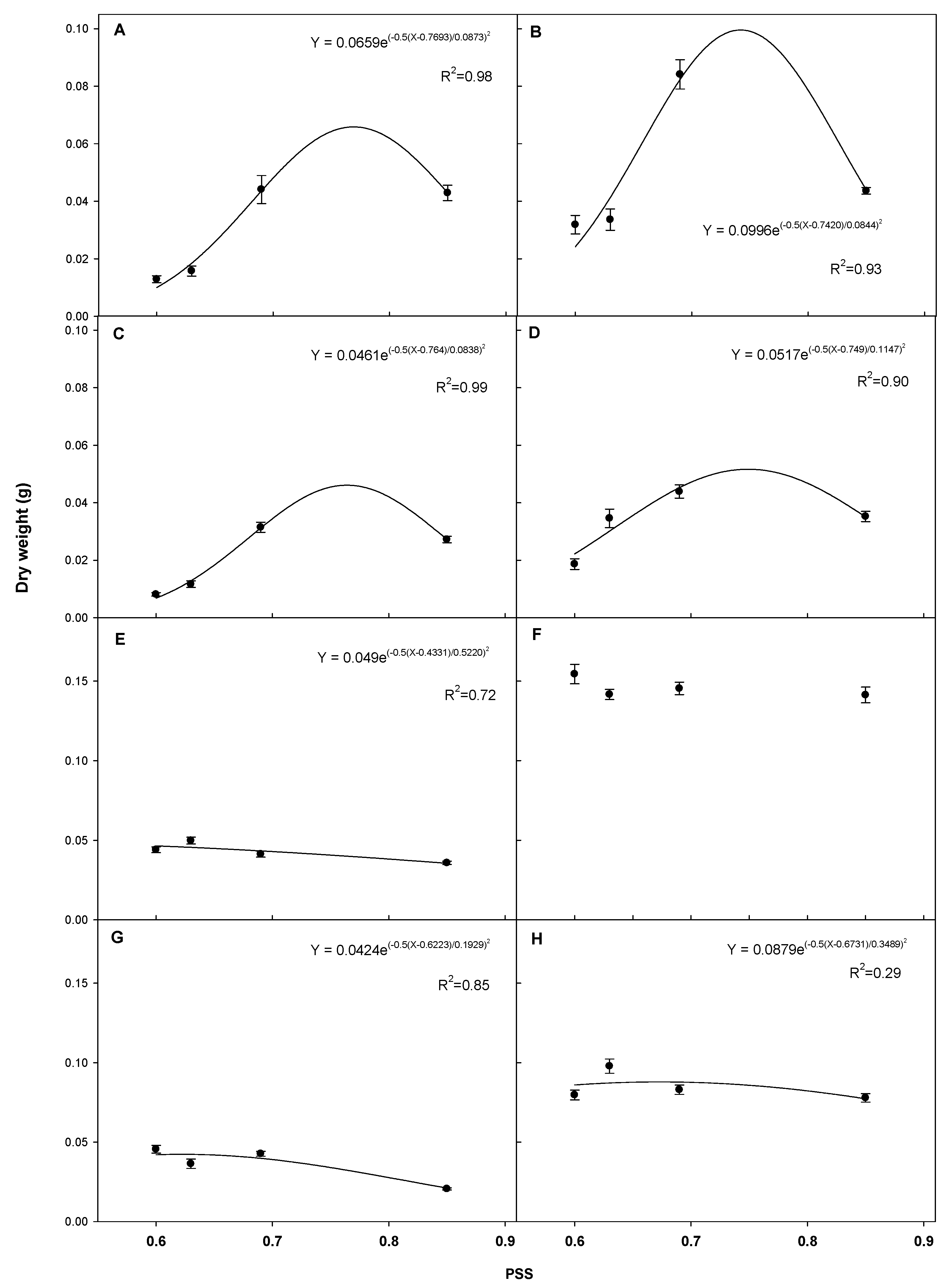
3.2. Plant Growth Analysis for Phytochrome Stationary State (PSS)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Carvalho, S.D.; Schwieterman, M.L.; Abrahan, C.E.; Colquhoun, T.A.; Folta, K.M. Light quality dependent changes in morphology, antioxidant capacity, and volatile production in sweet basil (Ocimum basilicum). Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef]
- Stutte, G.W.; Edney, S.; Skerritt, T.J.H. Photoregulation of bioprotectant content of red leaf lettuce with light-emitting diodes. HortScience 2009, 44, 79–82. [Google Scholar] [CrossRef]
- Lee, M.-J.; Park, S.-Y.; Oh, M.-M. Growth and cell division of lettuce plants under various ratios of red to far-red light-emitting diodes. Hortic. Environ. Biotechnol. 2015, 56, 186–194. [Google Scholar] [CrossRef]
- Lee, M.-J.; Son, K.-H.; Oh, M.-M. Increase in biomass and bioactive compounds in lettuce under various ratios of red to far-red LED light supplemented with blue LED light. Hortic. Environ. Biotechnol. 2016, 57, 139–147. [Google Scholar] [CrossRef]
- Chia, P.-L.; Kubota, C. End-of-day far-red light quality and dose requirements for tomato rootstock hypocotyl elongation. HortScience 2010, 45, 1501. [Google Scholar] [CrossRef]
- Yang, Z.-C.; Kubota, C.; Chia, P.-L.; Kacira, M. Effect of end-of-day far-red light from a movable LED fixture on squash rootstock hypocotyl elongation. Sci. Hortic. 2012, 136, 81–86. [Google Scholar] [CrossRef]
- Brown, C.S.; Schuerger, A.C.; Sager, J.C. Growth and photomorphogenesis of pepper plants under red light-emitting diodes with supplemental blue or far-red lighting. J. Am. Soc. Hortic. Sci. 1995, 120, 808. [Google Scholar] [CrossRef]
- Park, Y.; Runkle, E. Investigating the merit of including far-red radiation in the production of ornamental seedlings grown under sole-source lighting. In Proceedings of the VIII International Symposium on Light in Horticulture 1134, East Lansing, MI, USA, 22–26 May 2016; pp. 259–266. [Google Scholar]
- Graham, H.A.H.; Decoteau, D.R.J.S.h. Young watermelon plant growth responses to end-of-day red and far-red light are affected by direction of exposure and plant part exposed. Sci. Hortic. 1997, 69, 41–49. [Google Scholar] [CrossRef]
- Yang, F.; Feng, L.; Liu, Q.; Wu, X.; Fan, Y.; Raza, M.A.; Cheng, Y.; Chen, J.; Wang, X.; Yong, T.; et al. Effect of interactions between light intensity and red-to- far-red ratio on the photosynthesis of soybean leaves under shade condition. Environ. Exp. Bot. 2018, 150, 79–87. [Google Scholar] [CrossRef]
- Shibuya, T.; Endo, R.; Kitaya, Y.; Hayashi, S.J.H. Growth analysis and photosynthesis measurements of cucumber seedlings grown under light with different red to far-red ratios. HortScience 2016, 51, 843–846. [Google Scholar] [CrossRef]
- Kang, J.-H.; Jeon, B.-S. Effect of blue, red and far-red light on seedling growth and cotyledon chlorophyll content of Lagenaria siceraria Standl. In Proceedings of the Korean Society of Life Science Conference; 2001; pp. 26–27. [Google Scholar]
- Barreiro, R.; Guiamét, J.J.; Beltrano, J.; Montaldi, E.R. Regulation of the photosynthetic capacity of primary bean leaves by the red:far-red ratio and photosynthetic photon flux density of incident light. Physiol Plant. 1992, 85, 97–101. [Google Scholar] [CrossRef]
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi-Travier, S.; Crespel, L.; Morel, P.; Huché-Thélier, L.; Boumaza, R.; et al. Plant responses to red and far-red lights, applications in horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Zhen, S.; van Iersel, M.W. Far-red light is needed for efficient photochemistry and photosynthesis. J. Plant Physiol. 2017, 209, 115–122. [Google Scholar] [CrossRef]
- Ji, Y.; Ouzounis, T.; Courbier, S.; Kaiser, E.; Nguyen, P.T.; Schouten, H.J.; Visser, R.G.F.; Pierik, R.; Marcelis, L.F.M.; Heuvelink, E. Far-red radiation increases dry mass partitioning to fruits but reduces Botrytis cinerea resistance in tomato. Environ. Exp. Bot. 2019, 168, 103889. [Google Scholar] [CrossRef]
- Cole, B.; Kay, S.A.; Chory, J. Automated analysis of hypocotyl growth dynamics during shade avoidance in Arabidopsis. Plant J. 2011, 65, 991–1000. [Google Scholar] [CrossRef]
- Shinomura, T.; Uchida, K.; Furuya, M. Elementary processes of photoperception by phytochrome a for high-irradiance response of hypocotyl elongation in Arabidopsis. Plant Physiol. 2000, 122, 147–156. [Google Scholar] [CrossRef]
- Trupkin, S.A.; Legris, M.; Buchovsky, A.S.; Tolava Rivero, M.B.; Casal, J.J. Phytochrome B Nuclear bodies respond to the low red to far-red ratio and to the reduced irradiance of canopy shade in Arabidopsis. Plant Physiol. 2014, 165, 1698–1708. [Google Scholar] [CrossRef]
- Briggs, W.R.; Mandoli, D.F.; Shinkle, J.R.; Kaufman, L.S.; Watson, J.C.; Thompson, W.F. Phytochrome regulation of plant development at the whole plant, physiological, and molecular levels. In Sensory Perception and Transduction in Aneural Organisms; Springer: Boston, MA, USA, 1985; pp. 265–280. [Google Scholar]
- Otto, V.; Mousinger, E.; Sauter, M.; Schäfer, E. Phytochrome control of its own synthesis in Sorghum vulgarae and Avena sativa. Photochem. Photobiol. 1983, 38, 693–700. [Google Scholar] [CrossRef]
- VanDerWoude, W.J. A dimeric mechanism for the action of phytochrome; evidance from photothermal interactions in lettuce seed germination. Photochem. Photobiol. 1985, 42, 655–661. [Google Scholar] [CrossRef]
- Hennig, L.; Büche, C.; Schäfer, E. Degradation of phytochrome A and the high irradiance response in Arabidopsis: A kinetic analysis. Plant Cell Environ. 2000, 23, 727–734. [Google Scholar] [CrossRef]
- Nagy, F.; Schäfer, E. Phytochromes control photomorphogenesis by differentially regulated, interacting signaling pathways in higher plants. Plant Physiol. 2002, 53, 329–355. [Google Scholar] [CrossRef]
- Sager, J.C.; Smith, W.O.; Edwards, J.L.; Cyr, K.L. Photosynthetic efficiency and phytochrome photoequilibria determination using spectral data. Trans. ASAE 1988, 31, 1882–1889. [Google Scholar] [CrossRef]
- Smith, H. Sensing the light environment: The functions of the phytochrome family. In Photomorphogenesis in Plants; Springer: Dordrecht, The Netherlnds, 1994; pp. 377–416. [Google Scholar]
- Park, Y.; Runkle, E.S. Far-red radiation promotes growth of seedlings by increasing leaf expansion and whole-plant net assimilation. Environ. Exp. Bot. 2017, 136, 41–49. [Google Scholar] [CrossRef]
- Morgan, D.C.; Smith, H. Linear relationship between phytochrome photoequilibrium and growth in plants under simulated natural radiation. Nature 1976, 262, 210–212. [Google Scholar] [CrossRef]
- Smith, H.; Whitelam, G.; McCormac, A. Do the members of the phytochrome family have different roles? Physiological evidence from wild-type, mutant and transgenic plants. In Phytochrome Properties and Biological Action; Springer: Berlin/Heidelberg, Germany, 1991; pp. 217–236. [Google Scholar]
- Yamazaki, K. Nutrient Solution Culture; Pak-kyo Co.: Tokyo, Japan, 1982; Volume 41. [Google Scholar]
- Jeong, H.W.; Lee, H.R.; Kim, H.M.; Kim, H.M.; Hwang, H.S.; Hwang, S.J. Using light quality for growth control of cucumber seedlings in closed-type plant production system. Plants 2020, 9, 639. [Google Scholar] [CrossRef]
- Bae, J.-H.; Park, S.-Y.; Oh, M.-M. Supplemental irradiation with far-red light-emitting diodes improves growth and phenolic contents in Crepidiastrum denticulatum in a plant factory with artificial lighting. Hortic. Environ. Biotechnol. 2017, 58, 357–366. [Google Scholar] [CrossRef]
- Emerson, R. Dependence of yield of photosynthesis in long-wave red on wavelength and intensity of supplementary light. Proc. Sci. 1957, 125, 746. [Google Scholar]
- Ballaré, C.L.; Sánchez, R.A.; Scopel, A.L.; Casal, J.J.; Ghersa, C.M. Early detection of neighbour plants by phytochrome perception of spectral changes in reflected sunlight. Plant Cell Environ. 1987, 10, 551–557. [Google Scholar] [CrossRef]
- de Wit, M.; Spoel, S.H.; Sanchez-Perez, G.F.; Gommers, C.M.M.; Pieterse, C.M.J.; Voesenek, L.A.C.J.; Pierik, R. Perception of low red:far-red ratio compromises both salicylic acid- and jasmonic acid-dependent pathogen defences in Arabidopsis. Plant J. 2013, 75, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Gaba, V.; Black, M. Photocontrol of hypocotyl elongation in light-grown Cucumis sativus L. Planta 1985, 164, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Kalaitzoglou, P.; van Ieperen, W.; Harbinson, J.; van der Meer, M.; Martinakos, S.; Weerheim, K.; Nicole, C.C.S.; Marcelis, L.F.M. Effects of continuous or end-of-day far-red light on tomato plant growth, morphology, light absorption, and fruit production. Front. Plant Sci. 2019, 10, 322. [Google Scholar] [CrossRef] [PubMed]
- Mancinelli, A.L.; Rabino, I.J.T.B.R. The “high irradiance responses” of plant photomorphogenesis. Bot. Rev. 1978, 44, 129. [Google Scholar] [CrossRef]
- Schäfer, E. A new approach to explain the “high irradiance responses” of photomorphogenesis on the basis of phytochrome. J. Math. Biol. 1975, 2, 41–56. [Google Scholar] [CrossRef]
- Jose, A.M.; Vince-Frue, D. Light-induced changes in the photoresponses of plant stems the loss of a high irradiance response to far-red light. Planta 1977, 135, 95–100. [Google Scholar] [CrossRef]
- Holmes, M.G.; Schäfer, E. Action spectra for changes in the “high irradiance reaction” in hypocotyls of Sinapis alba L. Planta 1981, 153, 267–272. [Google Scholar] [CrossRef]
- Heim, B.; Schäfer, E. The effect of red and far-red light in the high irradiance reaction of phytochrome (hypocotyl growth in dark-grown Sinapis alba L.). Plant Cell Environ. 1984, 7, 39–44. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose–responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Fan, X.-X.; Xu, Z.-G.; Liu, X.-Y.; Tang, C.-M.; Wang, L.-W.; Han, X.-l. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Scientia Horticulturae 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Walton, L.J.; Reid, D.M. Interaction of red to far red light ratio and ethylene in regulating stem elongation of Helianthus annuus. Plant. Growth Regul. 2007, 51, 53–61. [Google Scholar] [CrossRef]
- Su, H.; Abernathy, S.D.; White, R.H.; Finlayson, S.A. Photosynthetic photon flux density and phytochrome B interact to regulate branching in Arabidopsis. Plant. Cell Environ. 2011, 34, 1986–1998. [Google Scholar] [CrossRef] [PubMed]
- Kajihara, S.; Katsutani, N. Effect of mother-stem length on cut flower stem yield and characteristic form during the high-rack training system for rose plants. Hort. Res. 2008, 7, 47–50. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Light Intensity (μmol∙m−2∙s−1) | PPF z | TPF y | PSS x | |||
|---|---|---|---|---|---|---|---|
| 400–500 nm | 500–600 nm | 600–700 nm | 700–800 nm | ||||
| W5F0 | 42.4 | 122.6 | 33.2 | 1.6 | 198.2 | 199.8 | 0.85 |
| W5F1 | 46.0 | 146.8 | 58.2 | 18.9 | 251.0 | 269.9 | 0.69 |
| W5F2 | 49.1 | 168.1 | 80.2 | 34.1 | 297.4 | 297.4 | 0.63 |
| W5F3 | 53.2 | 195.3 | 108.3 | 53.4 | 356.8 | 356.8 | 0.60 |
| Plant | Cultivar | Treatment | Hypocotyl Length (cm) | Stem Diameter (mm) | Shoot | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fresh Weight (g) | Dry Weight (g) | |||||||||
| Tomato | Dotaerangdia | W5F0 | 5.09 | b z | 16.1 | a | 0.947 | a | 0.049 | a |
| W5F1 | 6.22 | a | 16.7 | a | 0.678 | b | 0.044 | a | ||
| W5F2 | 4.94 | b | 10.3 | b | 0.245 | c | 0.015 | b | ||
| W5F3 | 4.97 | b | 11.3 | b | 0.210 | c | 0.012 | b | ||
| B-blocking | W5F0 | 5.74 | a | 17.7 | b | 0.725 | b | 0.043 | b | |
| W5F1 | 5.59 | a | 22.1 | a | 1.250 | a | 0.084 | a | ||
| W5F2 | 4.70 | b | 15.3 | c | 0.544 | c | 0.033 | bc | ||
| W5F3 | 4.68 | b | 15.3 | c | 0.478 | c | 0.031 | c | ||
| Pepper | Shinhong | W5F0 | 3.88 | c | 7.9 | b | 0.339 | b | 0.027 | b |
| W5F1 | 5.61 | a | 10.4 | a | 0.398 | a | 0.031 | a | ||
| W5F2 | 4.86 | ab | 7.1 | b | 0.156 | c | 0.011 | c | ||
| W5F3 | 4.28 | bc | 5.9 | c | 0.098 | d | 0.008 | d | ||
| Tantan | W5F0 | 3.53 | d | 9.1 | b | 0.389 | b | 0.035 | b | |
| W5F1 | 5.80 | a | 11.5 | a | 0.582 | a | 0.043 | a | ||
| W5F2 | 5.53 | b | 11.0 | a | 0.402 | b | 0.034 | b | ||
| W5F3 | 4.83 | c | 9.3 | b | 0.261 | c | 0.018 | c | ||
| Cucumber | Joeunbaegdadagi | W5F0 | 2.27 | c | 16.7 | b | 0.402 | c | 0.035 | c |
| W5F1 | 3.53 | b | 18.5 | b | 0.501 | b | 0.041 | b | ||
| W5F2 | 4.59 | a | 23.0 | a | 0.598 | a | 0.049 | a | ||
| W5F3 | 3.74 | b | 19.1 | b | 0.494 | b | 0.044 | b | ||
| Gourd | Heukjong | W5F0 | 2.93 | b | 26.7 | a | 1.742 | a | 0.014 | a |
| W5F1 | 4.75 | a | 29.0 | a | 1.910 | a | 0.014 | a | ||
| W5F2 | 4.91 | a | 28.0 | a | 1.743 | a | 0.014 | a | ||
| W5F3 | 4.57 | a | 28.6 | a | 1.875 | a | 0.015 | a | ||
| Watermelon | Sambokkul | W5F0 | 4.51 | d | 19.7 | b | 0.447 | c | 0.020 | c |
| W5F1 | 7.53 | c | 25.2 | a | 0.876 | b | 0.042 | a | ||
| W5F2 | 9.31 | a | 24.5 | a | 0.869 | b | 0.036 | b | ||
| W5F3 | 8.13 | b | 27.5 | a | 1.016 | a | 0.045 | a | ||
| Bottle gourd | Bulrojangsaeng | W5F0 | 3.71 | c | 30.9 | ab | 1.167 | c | 0.077 | b |
| W5F1 | 4.27 | b | 28.3 | b | 1.265 | bc | 0.083 | b | ||
| W5F2 | 5.40 | a | 35.9 | a | 1.565 | a | 0.097 | a | ||
| W5F3 | 5.14 | a | 31.3 | ab | 1.374 | b | 0.079 | b | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, H.; An, S.; Lee, B.; Chun, C. Improvement of Growth and Morphology of Vegetable Seedlings with Supplemental Far-Red Enriched LED Lights in a Plant Factory. Horticulturae 2020, 6, 109. https://doi.org/10.3390/horticulturae6040109
Hwang H, An S, Lee B, Chun C. Improvement of Growth and Morphology of Vegetable Seedlings with Supplemental Far-Red Enriched LED Lights in a Plant Factory. Horticulturae. 2020; 6(4):109. https://doi.org/10.3390/horticulturae6040109
Chicago/Turabian StyleHwang, Hyunseung, Sewoong An, Byungkwan Lee, and Changhoo Chun. 2020. "Improvement of Growth and Morphology of Vegetable Seedlings with Supplemental Far-Red Enriched LED Lights in a Plant Factory" Horticulturae 6, no. 4: 109. https://doi.org/10.3390/horticulturae6040109
APA StyleHwang, H., An, S., Lee, B., & Chun, C. (2020). Improvement of Growth and Morphology of Vegetable Seedlings with Supplemental Far-Red Enriched LED Lights in a Plant Factory. Horticulturae, 6(4), 109. https://doi.org/10.3390/horticulturae6040109