
Apple Tree Responses to Deficit Irrigation Combined with Periodic Applications of Particle Film or Abscisic Acid


Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Main Effects
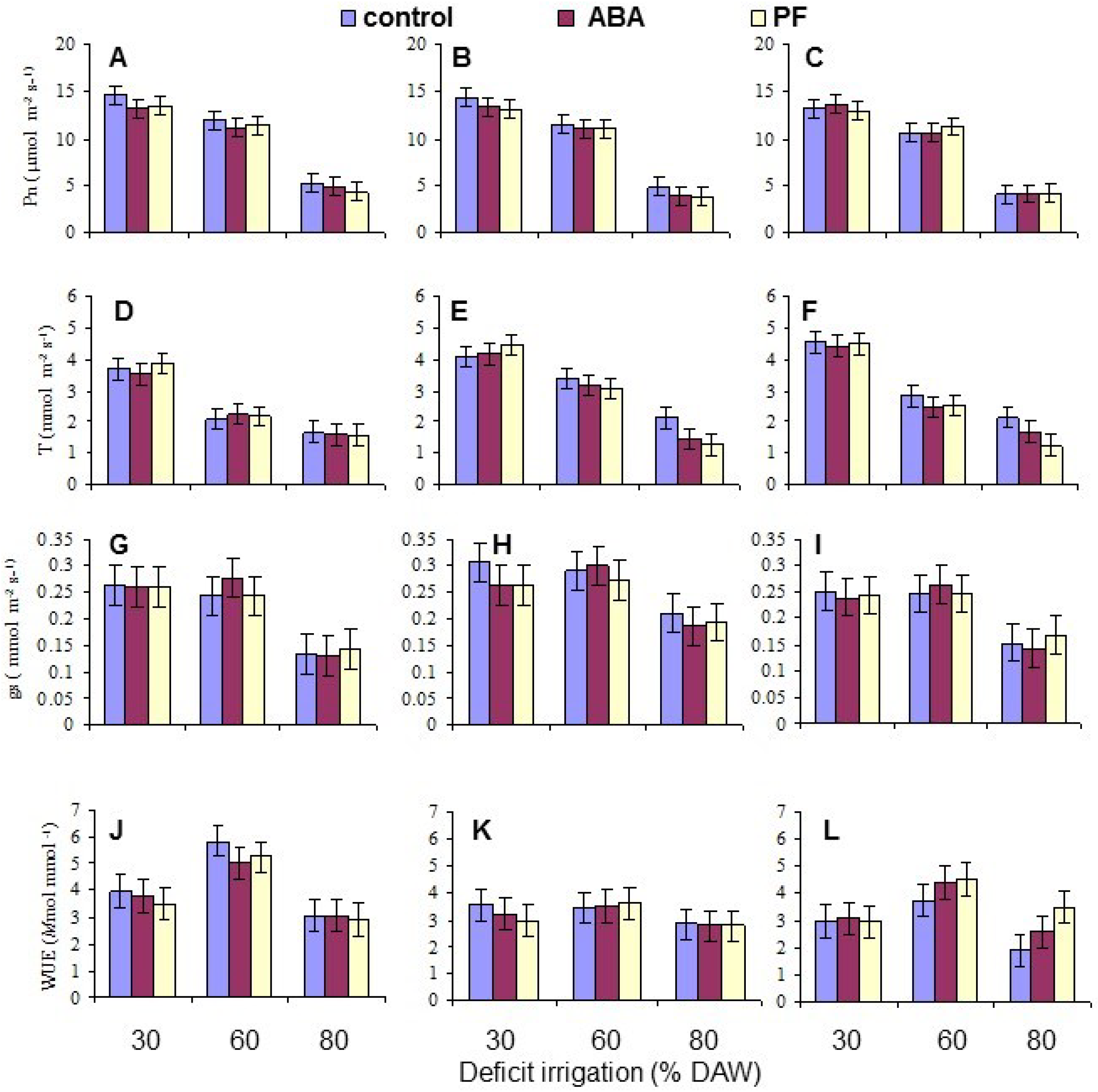
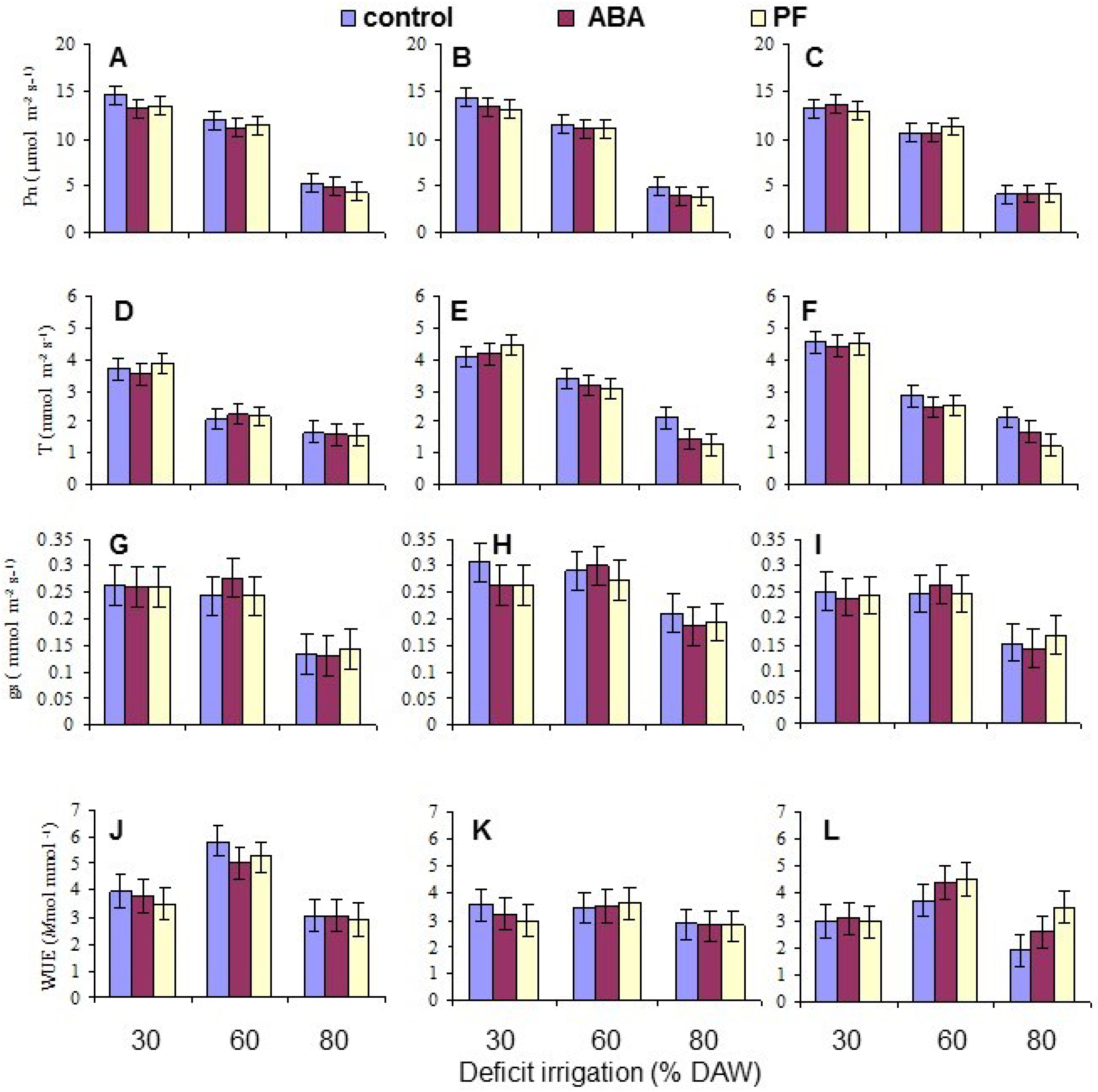
3.2. Leaf Gas Exchange
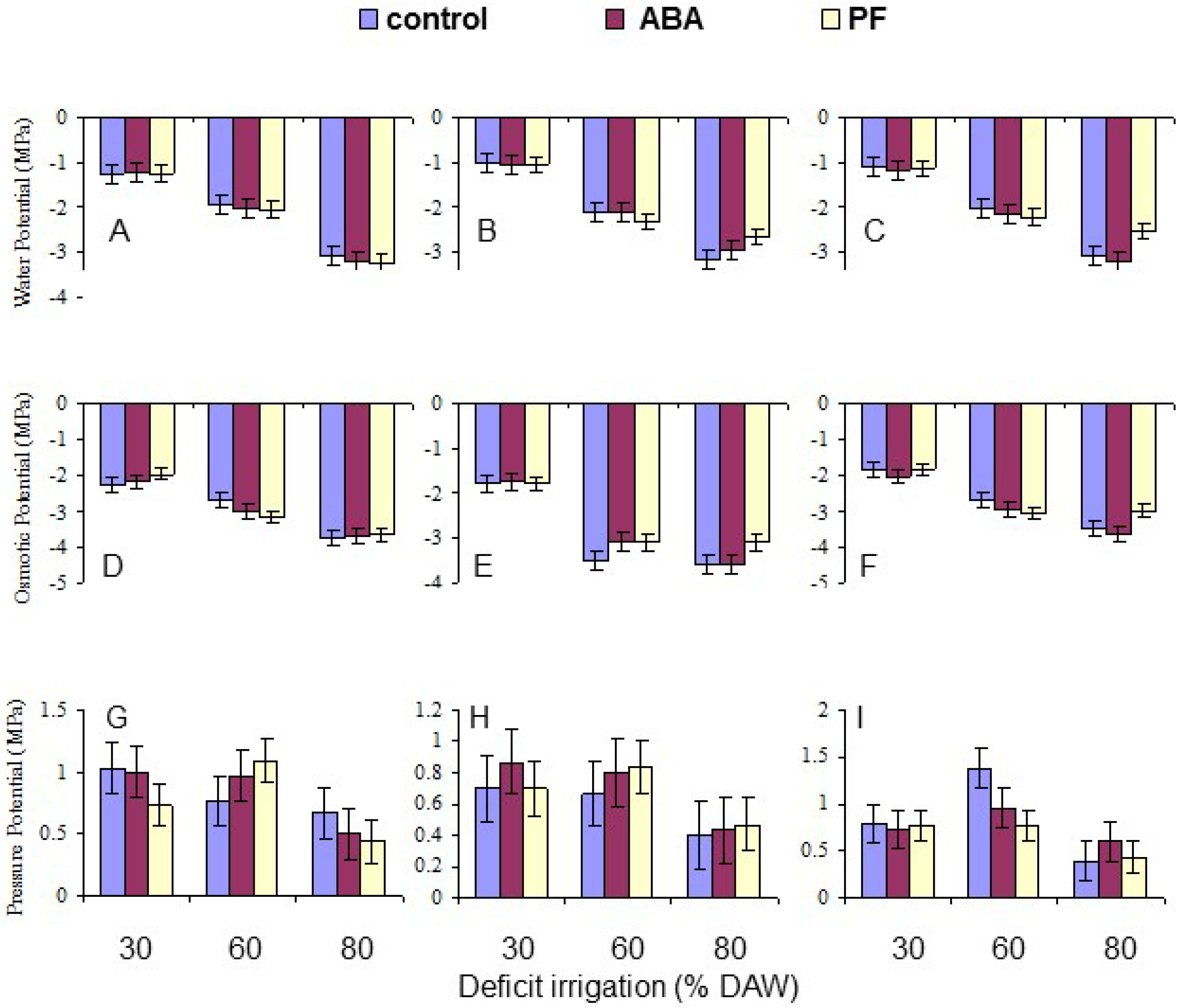
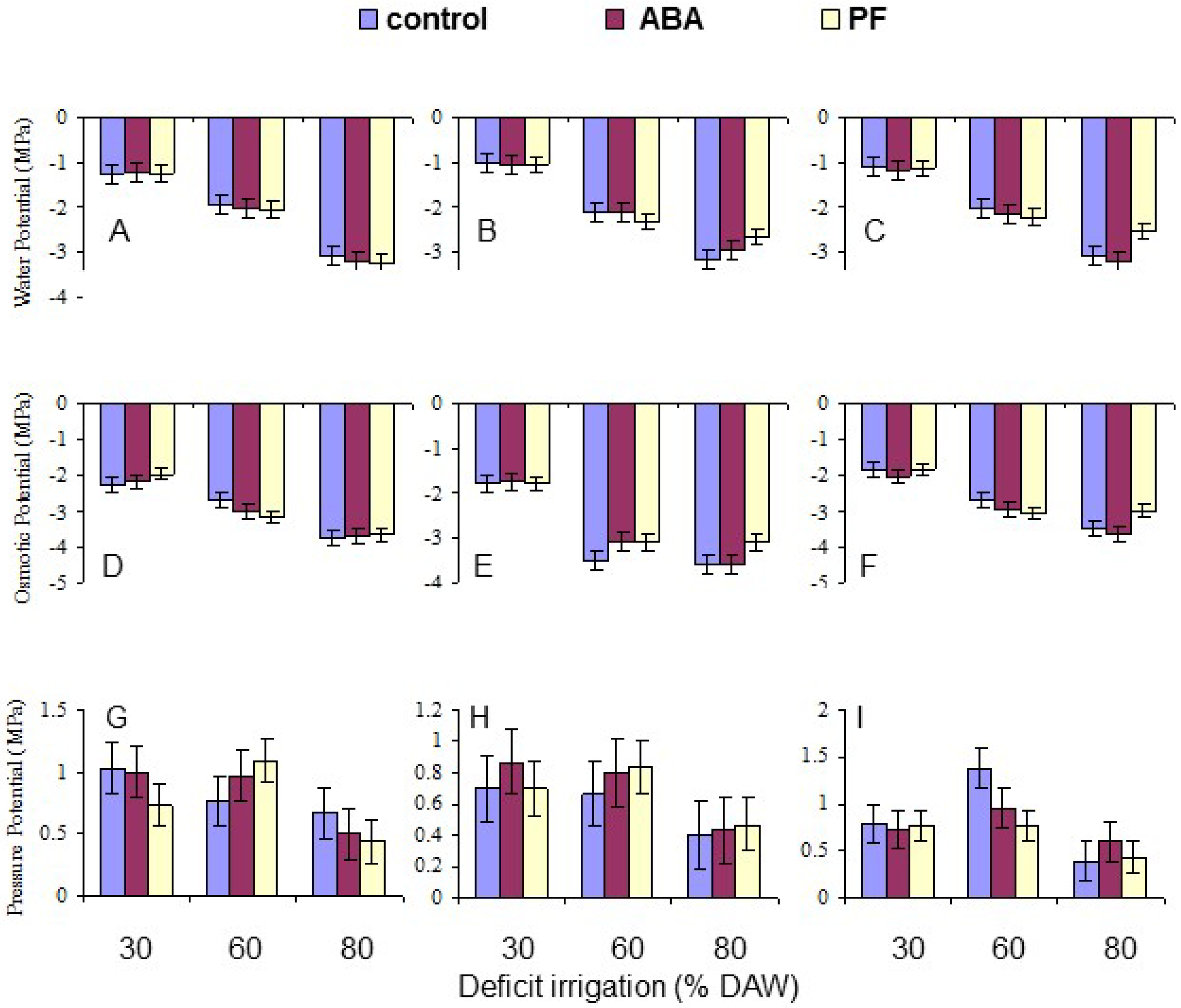
3.3. Leaf Water Relations
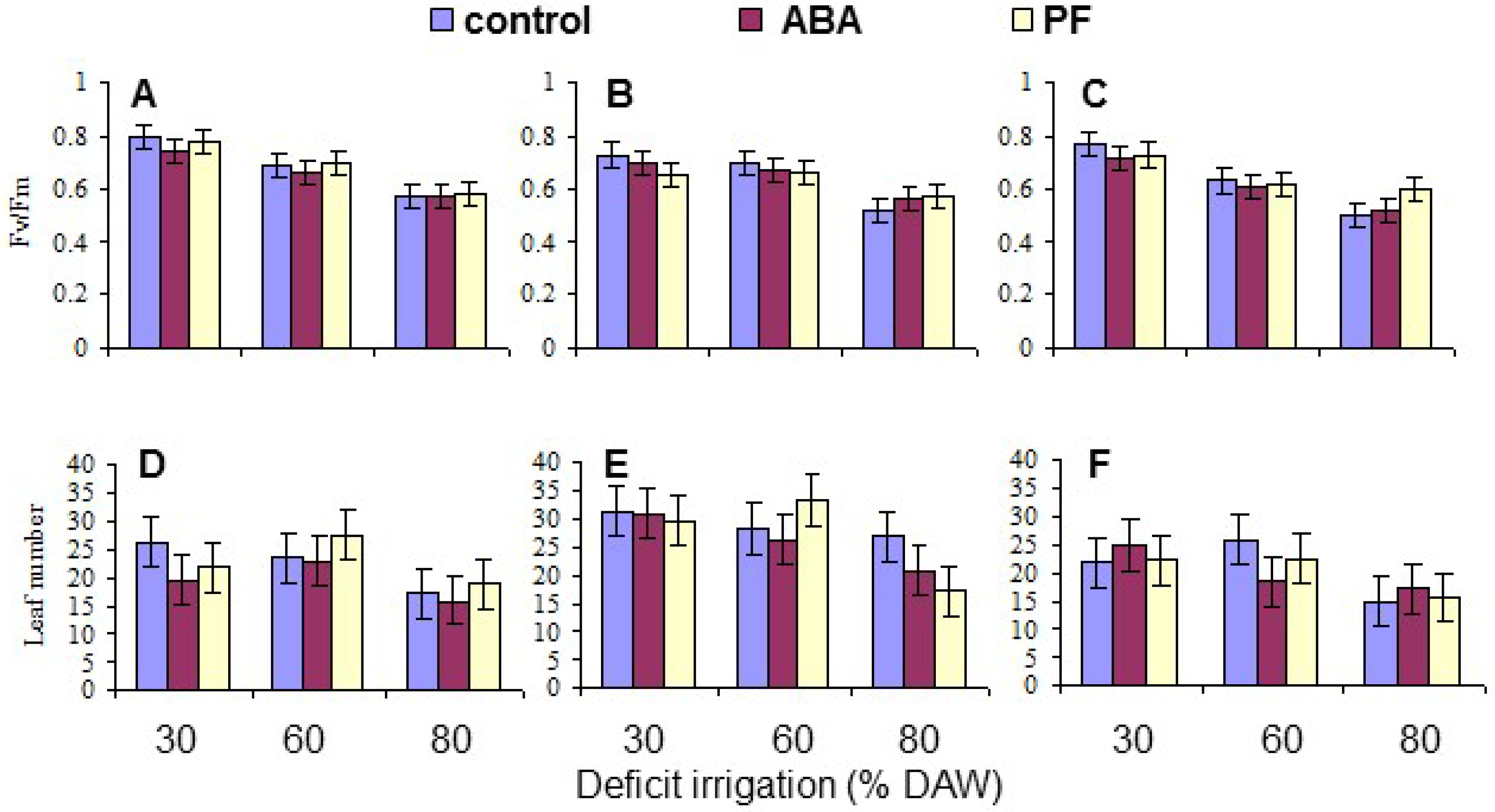
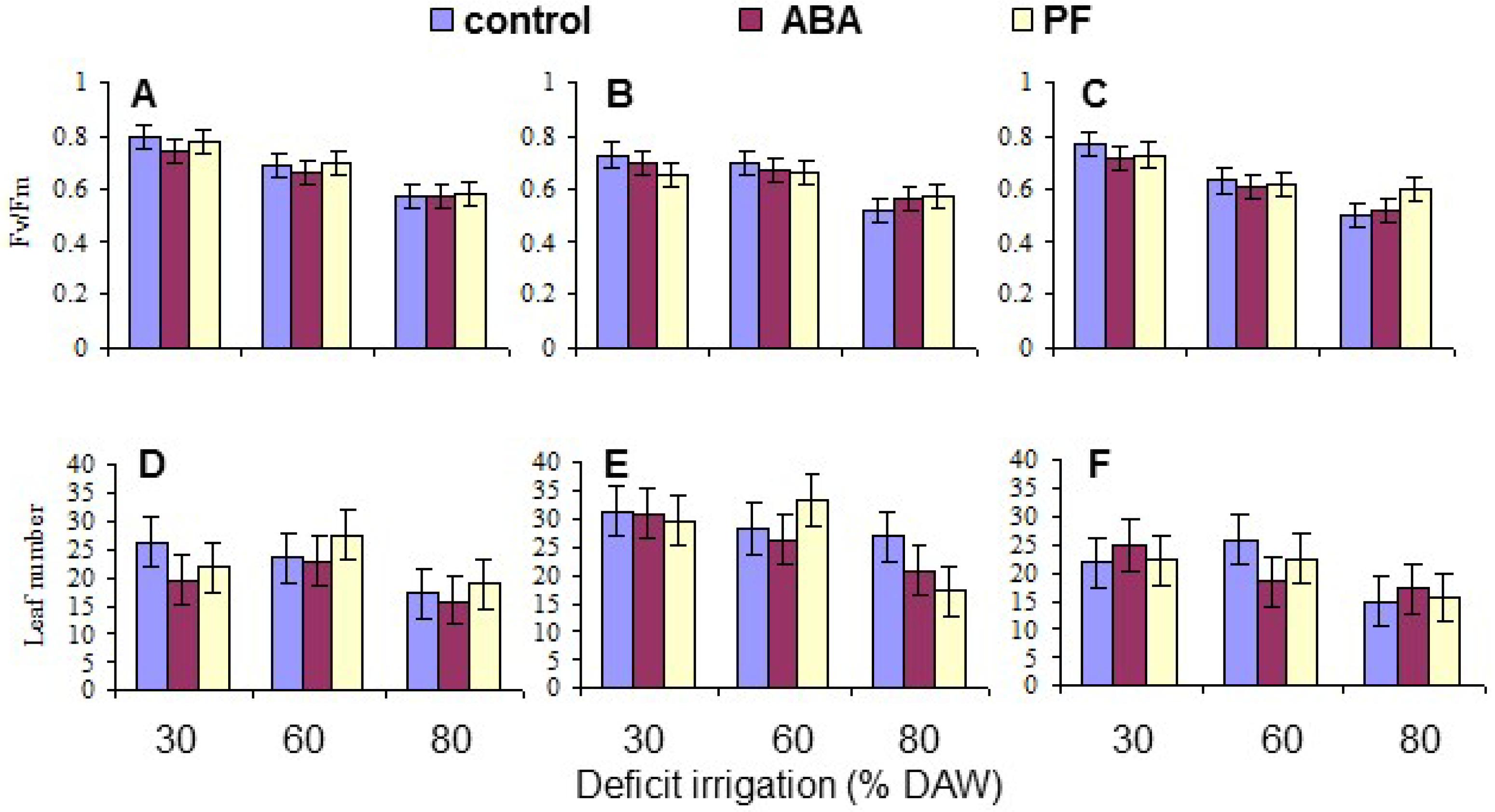
3.4. Leaf Fv/Fm and Number
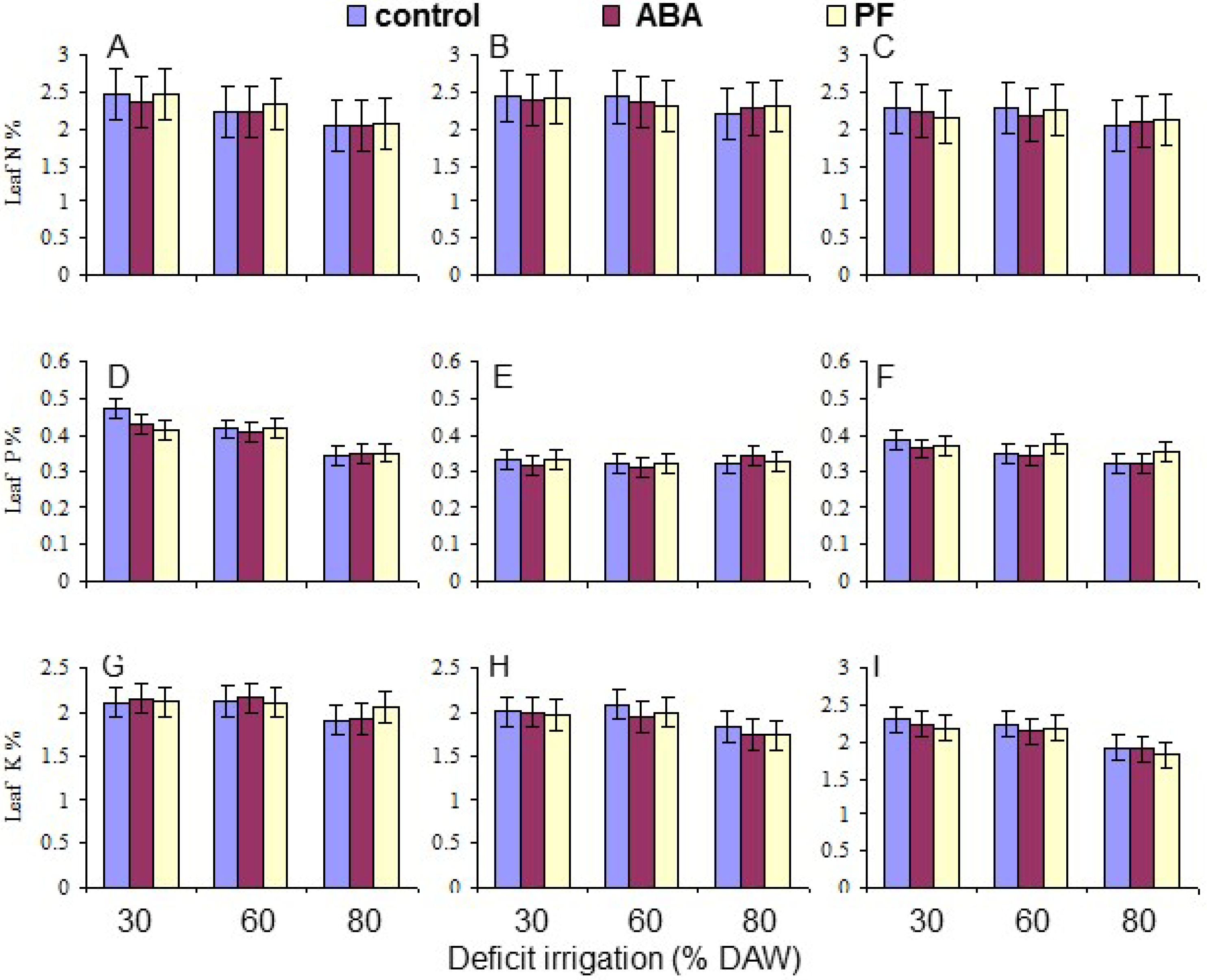
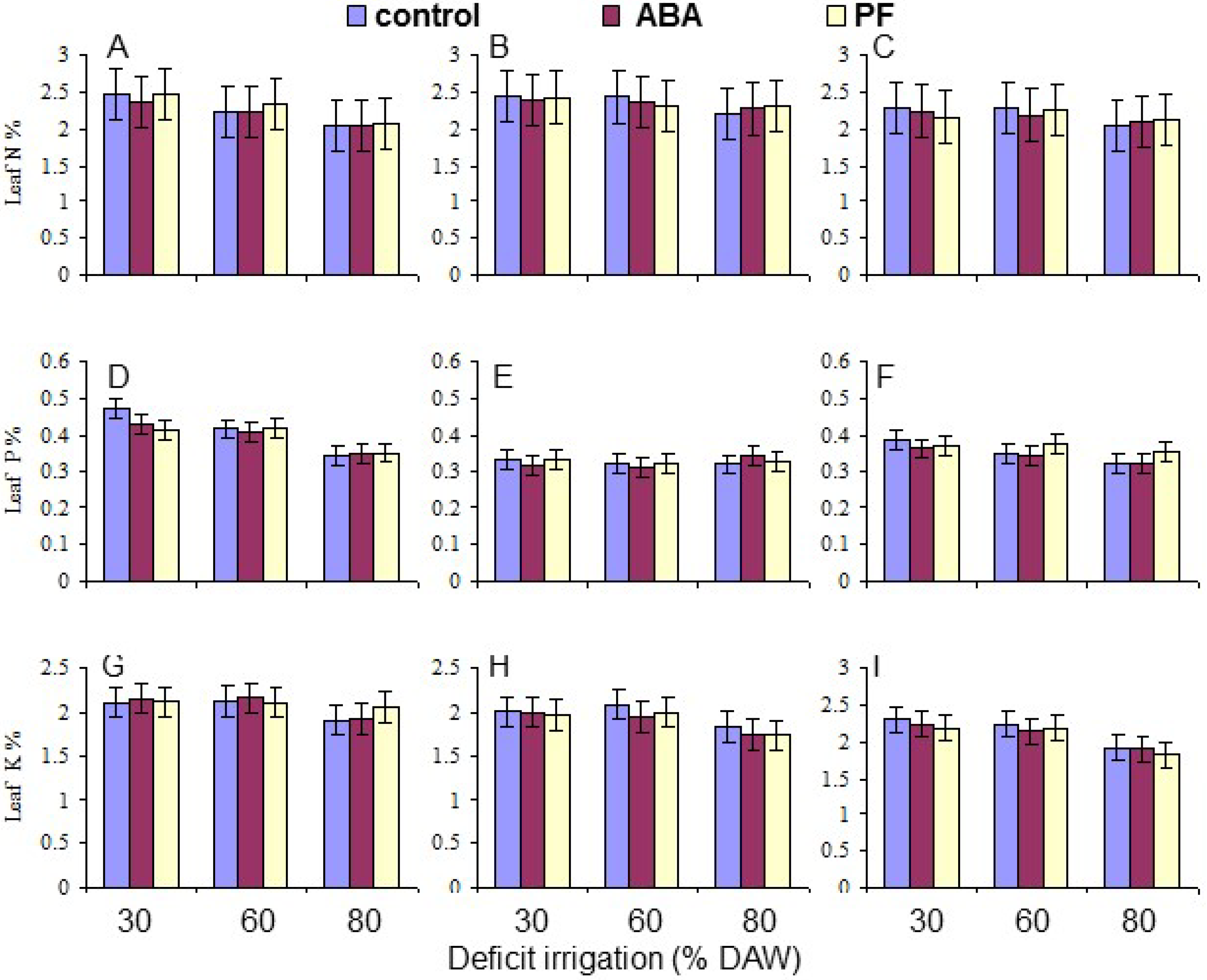
3.5. Leaf N, P, and K Content
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lobell, D.B.; Burke, M.B.; Tebaldi, C.; Mastrandrea, M.D.; Falcon, W.P.; Naylor, R.L. Prioritizing climate change adaptation needs for food security in 2030. Science 2008, 319, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Arar, A. Wastewater reuse for irrigation in the near east region. Water Sci. Technol. 1991, 23, 2127–2134. [Google Scholar]
- Geerts, S.; Raes, D. Deficit irrigation as an on-farm strategy to maximize crop water productivity in dry areas. Agric. Water Manag. 2009, 96, 1275–1284. [Google Scholar] [CrossRef]
- Aroca, R. Plant Responses to Drought Stress: From Morphological to Molecular Features; Springer: Berlin, Germany, 2012. [Google Scholar]
- Ludlow, M.M. Strategies of response to water stress. In Structural and Functional Responses to Environmental Stress; Kree, K.H., Richter, H., Minckley, T.M., Eds.; SPB Academic: The Hague, The Netherlands, 1989; pp. 269–281. [Google Scholar]
- Hanson, A.D.; Hitz, W.D. Metabolic responses of mesophytes to plant water deficits. Ann. Rev. Plant Physiol. 1982, 33, 163–203. [Google Scholar] [CrossRef]
- Li, F.; Cohen, S.; Naor, A.; Shaozong, K.; Erez, A. Studies of canopy structure and water use of apple trees on three rootstocks. Agric. Water Manag. 2002, 55, 1–14. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Policarpo, M.; Webster, A.D.; Kingswell, G. Drought tolerance of clonal Malus determined from measurements of stomatal conductance and leaf water potential. Tree Physiol. 2000, 20, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, R.T.; Perry, R.L.; Flore, J.A. Drought response of young apple trees on three rootstocks. II. Gas exchange, chlorophyll fluorescence, water relations, and leaf abscisic acid. J. Am. Soc. Hortic. Sci. 1997, 122, 841–848. [Google Scholar]
- Lombardini, L.; Toselli, M.; Flore, J.A. Carbon translocation and root respiration in potted apple trees during conditions of moderate drought. Acta Hortic. 2001, 557, 413–420. [Google Scholar] [CrossRef]
- Mohawesh, O.E.; Al-Absi, K.M. Physiological response of two apple genotypes to different water regimes under semiarid conditions. Adv. Hortic. Sci. 2009, 23, 158–165. [Google Scholar]
- Sircelj, H.; Tausz, M.; Grill, D.; Batic, F. Detecting different levels of drought stress in apple trees (Malus domestica Borkh.) with selected biochemical and physiological parameters. Sci. Hortic. 2007, 113, 362–369. [Google Scholar] [CrossRef]
- Kilili, A.W.; Behboudian, M.H.; Mills, T.M. Composition and quality of “Braeburn” apples under reduced irrigation. Sci. Hortic. 1996, 67, 1–11. [Google Scholar] [CrossRef]
- Behboudian, M.H.; Mills, T.M. Deficit irrigation in deciduous orchards. Hortic. Rev. 1997, 21, 105–130. [Google Scholar]
- Ebel, R.C.; Proebsting, E.L.; Evans, R.G. Deficit irrigation to control vegetative growth in apple and monitoring fruit growth to schedule irrigation. HortScience 1995, 30, 1229–1232. [Google Scholar]
- Irving, D.E.; Drost, J.H. Effects of water deficit on vegetative growth, fruit growth and fruit quality in Cox’s Orange Pippin apple. J. Hortic. Sci. 1987, 62, 427–432. [Google Scholar] [CrossRef]
- Mpelasoka, B.; Behboudian, M.; Green, S. Water use, yield and fruit quality of lysimeter-grown apple trees: Responses to deficit irrigation and to crop load. Irrig. Sci. 2001, 20, 107–113. [Google Scholar]
- Leib, B.G.; Caspari, H.W.; Redulla, C.A.; Andrews, P.K.; Jabro, J.J. Partial rootzone drying and deficit irrigation of “Fuji” apples in a semi-arid climate. Irrig. Sci. 2006, 24, 85–99. [Google Scholar] [CrossRef]
- O’Connell, M.G.; Goodwin, I. Responses of “Pink Lady” apple to deficit irrigation and partial rootzone drying: Physiology, growth, yield, and fruit quality. Aust. J. Agric. Res. 2005, 58, 1068–1076. [Google Scholar] [CrossRef]
- Glenn, D.M. Particle film mechanisms of action that reduce the effect of environmental stress in “Empire” apple. J. Am. Soc. Hortic. Sci. 2009, 134, 314–321. [Google Scholar]
- Glenn, D.M. Canopy gas exchange and water use efficiency of “Empire” apple in response to particle film, irrigation, and microclimatic factors. J. Am. Soc. Hortic. Sci. 2010, 135, 25–32. [Google Scholar]
- Glenn, D.M.; Erez, A.; Puterka, G.J.; Gundrum, P. Particle films affect carbon assimilation and yield in “Empire” apple. J. Am. Soc. Hortic. Sci. 2003, 128, 356–362. [Google Scholar]
- Glenn, D.M.; Puterka, G.J.; Drake, S.R.; Unruh, T.R.; Knight, A.L.; Baherle, P.; Prado, E.; Baugher, T.A. Particle film application influences apple leaf physiology, fruit yield, and fruit quality. J. Am. Soc. Hortic. Sci. 2001, 126, 175–181. [Google Scholar]
- Davies, W.J.; Zhang, J. Root signals and the regulation of growth and development of plants in drying soil. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 55–76. [Google Scholar] [CrossRef]
- Al-Absi, K.M. Gas exchange, chlorophyll and growth response of three orange genotypes (Citrus sinensis [L.] Osbeck) to abscisic acid under progressive water deficit. Jordan J. Agric. Sci. 2009, 5, 421–433. [Google Scholar]
- Du, Y.-L.; Wang, Z.-Y.; Fan, J.-W.; Turner, N.C.; He, J.; Wang, T.; Li, F.-M. Exogenous abscisic acid reduces water loss and improves antioxidant defence, desiccation tolerance and transpiration efficiency in two spring wheat genotypes subjected to a soil water deficit. Func. Plant Biol. 2013, 40, 494–506. [Google Scholar] [CrossRef]
- Ma, X.; Ma, F.; Mi, Y.; Ma, Y.; Shu, H. Morphological and physiological responses of two contrasting Malus species to exogenous abscisic acid application. Plant Growth Regul. 2008, 56, 77–87. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, B.; Xu, Q. Effects of abscisic acid on drought responses of Kentucky bluegrass. J. Am. Soc. Hortic. Sci. 2003, 128, 36–41. [Google Scholar]
- Waterland, N.L.; Campbell, C.A.; Finer, J.J.; Jones, M.L. Abscisic acid application enhances drought stress tolerance in bedding plants. HortScience 2010, 45, 409–413. [Google Scholar]
- Kondo, S.; Sugaya, S.; Sugawa, M.; Ninomiya, S.; Kittikorn, M.; Okawa, K.; Ohara, H.; Ueno, K.; Todoroki, Y.; Mizutani, M.; et al. Dehydration tolerance in apple seedlings is affected by an inhibitor of ABA 8-hydroxylase CYP707A. J. Plant Physiol. 2012, 169, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Jie, Z.; Yuncong, Y.; Streeter, J.G.; Ferree, D.C. Influence of soil drought stress on photosynthesis, carbohydrates and the nitrogen and phosphorus absorb in different section of leaves and stem of Fugi/M.9EML, a young apple seedling. Afr. J. Biotechnol. 2010, 9, 5320–5325. [Google Scholar]
- Wang, Z.; Quebedeaux, B.; Stutte, G.W. Osmotic adjustment: Effect of water stress on carbohydrates in leaves, stems and roots of apple. Aust. J. Plant Physiol. 1995, 22, 747–754. [Google Scholar] [CrossRef]
- Shellie, K.; Glenn, D.M. Wine grape response to foliar particle film under differing levels of preveraison water stress. HortScience 2008, 43, 1392–1397. [Google Scholar]
- Shellie, K.C.; King, B.A. Kaolin-based foliar reflectant and water deficit influence Malbec leaf and berry temperature, pigments, and photosynthesis. Am. J. Enol. Vit. 2013, 64, 223–230. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Archbold, D.D. Water relations of a Fragaria chiloensis and a F. virginiana selection respond to water deficit stress. J. Am. Soc. Hortic. Sci. 1993, 118, 280–285. [Google Scholar]
- Tandon, H.L.S. Methods of Analysis of Soils, Plants, Waters and Fertilizers; Fertilizer Development and Consultation Organization: New Delhi, India, 1995; pp. 49–82. [Google Scholar]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Misra, A.K.; Das, B.K.; Datta, J.K.; De, G.C. Influence of antitranspirants on photosynthesis, leaf dry wt., nitrate reductase activity and leaf yield of mulberry (Morus alba L.) under water stress condition. Indian J. Agric. Res. 2009, 43, 144–147. [Google Scholar]
- Grange, M.; Wand, S.J.; Theron, K.L. Effect of kaolin applications on apple fruit quality and gas exchange of apple leaves. Acta Hortic. 2004, 636, 545–550. [Google Scholar] [CrossRef]
- Prive, J.P.; Russell, L.; LeBlanc, A. The impact of kaolin clay sprays on leaf gas exchange of Ginger Gold apple trees in New Brunswick. Can. J. Plant Sci. 2006, 86, 1377–1381. [Google Scholar] [CrossRef]
- Lombardini, L.; Glenn, D.M.; Harris, M.K. Application of kaolin-based particle film on pecan trees: Consequences on leaf gas exchange, stem water potential, nut quality, and insect populations. HortScience 2004, 39, 857–858. [Google Scholar]
- Rosati, A.; Metcalf, S.G.; Buchner, R.P.; Fulton, A.E.; Lampinen, B.D. Physiological effects of kaolin applications in well-irrigated and water-stressed walnut and almond trees. Ann. Bot. 2006, 98, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Wunsche, J.N.; Lombardini, L.; Greer, D.H. Surround particle film applications—Effects on whole canopy physiology of apple. Acta Hortic. 2004, 636, 565–571. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W.; Grace, S.C. Physiology of light tolerance in plants. Hortic. Rev. 1997, 18, 215–246. [Google Scholar]
- Johnson, J.D.; Tognetti, R.; Paris, P. Water relations and gas exchange in poplar and willow under water stress and elevated atmospheric CO2. Physiol. Plant. 2002, 115, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Chu, L.; Jaleel, C.A.; Zhao, C. Water-deficit stress-induced anatomical changes in higher plants. Plant Biol. Path. 2008, 331, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Cabuslay, G.S.; Ito, O.; Alejar, A.A. Physiological evaluation of responses of rice (Oryza sativa L.) to water deficit. Plant Sci. 2002, 63, 815–827. [Google Scholar] [CrossRef]
- Nilsen, E.T.; Orcutt, D.M. The Physiology of Plant under Stress; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Zeevaart, J.A.D.; Creelman, R.A. Metabolism and physiology of abscisic acid. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1998, 39, 439–473. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Trait | Main Effects and Interactions | ||||||
|---|---|---|---|---|---|---|---|
| G | I | A | G × I | G × A | I × A | G × I × A | |
| Pn | NS | <10−4 | NS | NS | NS | 0.0082 | NS |
| T | <10−4 | <10−4 | NS | 0.0002 | NS | NS | NS |
| gs | 0.0037 | <10−4 | NS | NS | NS | NS | NS |
| WUE | 0.0001 | <10−4 | NS | 0.0180 | NS | NS | NS |
| Ψw | 0.0091 | <10−4 | NS | NS | NS | NS | NS |
| Ψπ | 0.0321 | <10−4 | NS | 0.0189 | NS | NS | NS |
| Ψp | 0.0346 | <10−4 | NS | NS | NS | NS | 0.0255 |
| Fv/Fm | 0.0091 | <10−4 | NS | NS | NS | NS | NS |
| Leaf Number | <10−4 | <10−4 | NS | NS | NS | NS | NS |
| N | 0.0002 | <10−4 | NS | NS | NS | NS | NS |
| P | <10−4 | 0.0001 | NS | 0.0025 | NS | NS | NS |
| K | 0.0051 | <10−4 | NS | NS | NS | NS | NS |
| Ca | NS | NS | NS | NS | NS | NS | NS |
| Mg | NS | NS | NS | NS | NS | NS | NS |
| Leaf Trait | Genotype | Deficit Irrigation (% DAW) | ||||
|---|---|---|---|---|---|---|
| GS | RG | GD | 30 | 60 | 80 | |
| Photosynthesis (Pn) (μmol·CO2·m−2·s−1) | 10.0 NS | 9.7 | 9.4 | 13.6 a | 11.2 b | 4.3 c |
| Transpiration (T) (mmol·H2O·m−2·s−1) | 2.50 c | 3.06 a | 2.92 b | 4.14 a | 2.67 b | 1.67 c |
| Stomatal conductance (gs) (mmol·H2O·m−2·s−1) | 0.217 b | 0.254 a | 0.217 b | 0.261 a | 0.265 a | 0.162 b |
| Water use efficiency (Pn/T) | 4.04 a | 3.20 b | 3.29 b | 3.34 b | 4.37 a | 2.81 b |
| Genotype | Deficit Irrigation (% DAW) | ||
|---|---|---|---|
| 30 | 60 | 80 | |
| Transpiration (mmol·H2O·m−2·s−1) | |||
| Granny Smith | 3.70 b | 2.17 e | 1.63 f |
| Royal Gala | 4.23 a | 3.23 c | 1.70 f |
| Golden Delicious | 4.50 a | 2.60 d | 1.67 f |
| Water use efficiency (net photosynthetic rate/transpiration rate) | |||
| Granny Smith | 3.74 c | 5.37 a | 3.01 ef |
| Royal Gala | 3.26 de | 3.54 cd | 2.78 fg |
| Golden Delicious | 3.00 ef | 4.22 b | 2.66 g |
| Leaf osmotic potential (Ψπ, MPa) | |||
| Granny Smith | −2.14 d | −2.97 c | −3.71 a |
| Royal Gala | −1.79 e | −3.23 b | −3.41 b |
| Golden Delicious | −1.89 e | −2.91 c | −3.38 b |
| Antitranspirant | Deficit Irrigation (% DAW) | ||
|---|---|---|---|
| 30 | 60 | 80 | |
| Photosynthetic rate (μmol·CO2·m−2·s−1) | |||
| None | 14.1 a | 11.4 c | 4.6 d |
| Abscisic acid | 13.4 b | 11.0 c | 4.3 d |
| Particle film | 13.2 b | 11.3 c | 4.1 e |
| Leaf Water Relations | Genotype | Deficit Irrigation (% DAW) | ||||
|---|---|---|---|---|---|---|
| Component (MPa) | GS | RG | GD | 30 | 60 | 80 |
| Water potential (Ψw) | −2.14 a | −2.05 b | −2.07 b | −1.12 c | −2.12 b | −3.02 a |
| Osmotic potential (Ψπ) | −2.94 a | −2.81 b | −2.72 c | −1.94 b | −3.04 a | −3.50 a |
| Pressure potential (Ψp) | 0.80 a | 0.76 a | 0.65 b | 0.81 a | 0.92 a | 0.48 b |
| Leaf Trait | Genotype | Deficit Irrigation (% DAW) | ||||
|---|---|---|---|---|---|---|
| GS | RG | GD | 30 | 60 | 80 | |
| Fv/Fm | 0.674 a | 0.643 b | 0.634 c | 0.737 a | 0.661 b | 0.558 c |
| Leaf number | 21.6 b | 27.2 a | 20.5 b | 25.5 a | 25.5 a | 18.4 b |
| N content | 2.24 bc | 2.34 ab | 2.18 c | 2.37 a | 2.28 a | 2.12 b |
| P content | 0.40 a | 0.33 c | 0.36 b | 0.38 a | 0.37 b | 0.34 c |
| K content | 2.09 a | 1.91 b | 2.10 a | 2.13 a | 2.11 a | 1.86 b |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Absi, K.M.; Archbold, D.D. Apple Tree Responses to Deficit Irrigation Combined with Periodic Applications of Particle Film or Abscisic Acid. Horticulturae 2016, 2, 16. https://doi.org/10.3390/horticulturae2040016
Al-Absi KM, Archbold DD. Apple Tree Responses to Deficit Irrigation Combined with Periodic Applications of Particle Film or Abscisic Acid. Horticulturae. 2016; 2(4):16. https://doi.org/10.3390/horticulturae2040016
Chicago/Turabian StyleAl-Absi, Khalid M., and Douglas D. Archbold. 2016. "Apple Tree Responses to Deficit Irrigation Combined with Periodic Applications of Particle Film or Abscisic Acid" Horticulturae 2, no. 4: 16. https://doi.org/10.3390/horticulturae2040016
APA StyleAl-Absi, K. M., & Archbold, D. D. (2016). Apple Tree Responses to Deficit Irrigation Combined with Periodic Applications of Particle Film or Abscisic Acid. Horticulturae, 2(4), 16. https://doi.org/10.3390/horticulturae2040016