Abstract
Insect parasitoids are key biological agents within terrestrial ecosystems, offering a promising avenue for controlling insect pests. Hyperparasitoids are a group of insects that lay their eggs in or on the body of parasitoid hosts, which can greatly hamper the effectiveness of parasitoids. To optimize their reproductive success, adult parasitoids/hyperparasitoids must find sufficient food sources and mate partners (when they do not reproduce parthenogenetically) and locate suitable hosts for their offspring. To complete these tasks, parasitoids largely rely on their ability to detect relevant chemical cues (semiochemicals or infochemicals). In the last three decades, the identities of semiochemicals and their ethological significance have been widely characterized, and the possibility of using these chemical cues in insect pest management has received a lot of attention. Insects have evolved a highly sensitive and sophisticated chemosensory system adept at navigating complex and dynamic chemical environments. In this review, we first summarize the semiochemicals used by insect parasitoids, primarily including semiochemicals involved in food location, host foraging, and mate finding, while also addressing semiochemicals employed by hyperparasitoids. Next, we discuss recent progress in elucidating the chemosensory mechanisms underlying parasitoid responses to semiochemicals, with a focus on olfactory and gustatory pathways. Finally, we evaluate the potential applications of semiochemicals in pest management, highlighting the roles of parasitoids and hyperparasitoids. This paper aims to establish a theoretical framework for the effective employment of parasitoids in biological control of insect pests.
1. Introduction
The control of insect pests relies predominantly on the application of chemical pesticides. However, prolonged use of chemical pesticides has resulted in progressive increases in pest resistance, posing a serious threat to food safety and ecological security [1]. Therefore, there is an urgent need to develop efficient and environmentally sustainable pest management strategies. Using insects’ natural enemies, including parasitoids and predators, is a key strategy in biological pest control, offering a sustainable alternative to chemical pesticides [2]. Parasitoids are insects whose larvae develop in or on the bodies of other arthropods (mostly other insects), whereas the adults are free living. Most parasitoids are quite specialized and only attack a limited number of related host species, which minimizes the risk of harming non-target organisms of the ecosystem. This makes parasitoids key biocontrol agents within terrestrial ecosystems, offering a promising avenue for controlling agricultural and environmental pests [3]. For instance, in the Huang-Huai River Basin in China, the ichneumonid wasp Campoletis chlorideae Uchida (Hymenoptera: Ichneumonidae) parasitizes and kills up to 63.1% of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) larvae [4]. Parasitoids have their own specialized natural enemies, i.e., hyperparasitoids. Hyperparasitoids are a group of insects in the fourth trophic level which lay their eggs in or on the body of other parasitoid hosts, which can greatly hamper the effectiveness of parasitoids [5].
Parasitoids and hyperparasitoids base foraging decisions on information from different trophic levels, and semiochemicals serve as critical cues [6,7,8,9]. In recent decades, numerous semiochemicals associated with the behavior of insect parasitoids/hyperparasitoids have been identified. For example, in the life cycle of the egg parasitoids (Hymenoptera) after emergence, they rely on semiochemicals to locate suitable mates and hosts, a process critical for successful mating and parasitism. In the egg parasitoid Nasonia vitripennis (Walker) (Hymenoptera: Pteromalidae), male-derived pheromones (4R,5R)-5-hydroxy-4-decanolide, (4R,5S)-5-hydroxy-4-decanolide, and 4-methylquinazoline induce mating behaviors of females [10,11,12]. Maize volatiles, such as (3E)-4,8-dimethyl-1,3,7-nonatriene (DMNT), (3E,7E)-4,8,12-trimethyl-1,3,7,11-tridecatetraene (TMTT), and (E)-(1R,9S)-caryophyllene, induced by oviposition of the stemborer Chilo partellus (Swinhoe) (Lepidoptera: Pyralidae), could effectively attract the egg parasitoid Trichogramma bournieri Pintureau & Babault (Hymenoptera: Trichogrammatidae) [13]. Furthermore, some studies have verified that the use of semiochemicals can modify the behavior of parasitoids/hyperparasitoids, opening new perspectives in using semiochemicals to improve/curb the efficiency of parasitoids/hyperparasitoids [5,14,15,16,17,18]. Insects rely on chemosensory perception to locate food, hosts, and mate partners [18,19]. The understanding of semiochemicals used by parasitoids/hyperparasitoids in complex chemical environments and the chemosensory mechanism underlying semiochemical perception could facilitate the exploitation of parasitoids as bio-control agents. In this review, we systematically explore the semiochemicals used by parasitoid/hyperparasitoids wasps for food location, host foraging, and mate finding. We also summarize the chemosensory genes implicated in their detection and discuss the practical applications of these semiochemicals in insect pest management.
2. Semiochemicals for Food Location and Mate Finding
Although a minority of parasitoid species feed on their larval host to obtain protein, most adult parasitoids seek food sources that are enriched with carbohydrates, such as floral nectar and honeydews [20]. Plant volatiles commonly occurring in floral blends are the main triggers of food foraging behaviors in insect parasitoids [21]. One of such commonly occurring compounds is linalool [22]. This compound is an abundantly released component of floral volatiles. In the evergreen shrub Daphne mezereum L. (Myrtales: Thymelaeaceae), linalool accounted for 95% of total floral fragrance [23]. Linalool is known to be attractive to some parasitoid species [23,24]. For example, behavioral assays demonstrated that linalool is a semiochemical for food foraging of the ichneumonid wasp C. chlorideae [25].
Mate-finding strategies in parasitoids are mainly based on the use of intraspecific sex pheromones [26,27,28]. Parasitoids reproduce in the following ways: the newly emerged females reproduce parthenogenetically, take risks of producing haploid male offspring, or they reproduce sexually, yielding both male and female offspring [29,30,31]. The latter strategy is more commonly executed. For example, virgin females of the parasitoid Cotesia vestalis (Halliday) (Hymenoptera: Braconidae) are not attracted by herbivore-induced plant volatiles (HIPVs) until they mate [32]. Virgin males of the aphid parasitoid Lysiphlebus testaceipes (Cresson) (Hymenoptera: Aphidiinae) are only attracted by conspecific females but not by aphid-induced plant volatiles [33].
Currently, however, sex pheromone communication in parasitoids is less studied. Females of Ascogaster quadridentata Wesmael (Hymenoptera: Braconidae), a braconid parasitoid of codling moth Cydia pomonella L. (Lepidoptera: Tortricidae), release (Z,Z)-9,12-octadecadienal to attract males [34]. As we mentioned before, in N. vitripennis, male-derived pheromones (4R,5R)-5-hydroxy-4-decanolide, (4R,5S)-5-hydroxy-4-decanolide, and 4-methylquinazoline induce mating behaviors of females [10,11,12,31]. (E)-bergamotene serves as a male-produced sex pheromone in Melittobia digitata Dahams (Hymenoptera: Eulophidae) [35]. In the females of Dibrachys cavus (Walker) (Hymenoptera: Pteromalidae), the cuticular hydrocarbon 3-methylalkane is suggested as a key component of the contact sex pheromones [36]. A recent work by Guo et al. found that virgin males of C. chlorideae were attracted to the female-derived sex pheromones tetradecanal and 2-heptadecanone [37].
3. Semiochemicals for Host Foraging
3.1. HIPVS
Host foraging is crucial for the reproductive success of parasitoids. They locate hosts through a progressive process where HIPVs act as instrumental information bases [9,24,38,39,40]. The composition of HIPVs has been elucidated in numerous plant species by gas chromatography–mass spectrometry (GC-MS). Compared to undamaged plants, those infested by herbivore insects released an increased quantity and variety of volatile compounds [41,42,43,44,45]. According to the published data, HIPVs mainly comprise terpenoids, green leaf volatiles (GLVs), aromatic compounds, and amino acid derivatives.
Terpenoids, produced from isoprene pyrophosphate, are regarded as the most abundant class of compounds emitted by herbivore-infested plants. Major terpenoid volatiles emitted from plants include monoterpenoids such as geraniol, (E)-β-ocimene, and citral and sesquiterpenoids such as farnesol and (E)-β-caryophyllene. Additionally, two terpenoids with irregular structures, i.e., DMNT and TMTT, frequently occur in HIPVs [46,47,48,49,50,51]. GLVs, encompassing a range of 6-carbon aldehydes, alcohols, and esters, are generated through the oxidative degradation of leaf lipids and typically exhibit a grassy scent. Several GLVs, such as (Z)-2-hexenal, (Z)-3-hexenol, (E)-2-hexenyl acetate, and (Z)-3-hexenyl acetate, are commonly detected in the HIPVs emitted by various plant species such as maize, soybean, and wheat [41,52,53,54]. Aromatic compounds, derived from phenylalanine metabolism, consist of aromatic rings and different side structures. Additionally, indole derivatives, such as methyl salicylate (MeSA) and indole, arise as byproducts in biosynthesis of the amino acid tryptophan. Amino acid derivatives, synthesized from amino acids, include a diverse assay of amines, oximes, and nitriles. These compounds are often less captured in HIPVs compared to the other three classes of compounds [52,55,56].
Numerous studies indicated that parasitoids use two different modes to detect HIPVs: the “species-specific odor” recognition mode and the “compound ratio” recognition mode [56,57,58,59]. In the “species-specific odor” mode, parasitoids recognize compounds unique to a single species or a closely related group. This is exemplified by herbivorous insects specialized on Brassicaceae, such as the braconid wasp Diaeretiella rapae (M’Intosh) (Hymenoptera: Braconidae), a specialist parasitoid of aphids on Brassicaceae plants (Brassicales). Studies have demonstrated that D. rapae exhibits chemotaxis responses to various isothiocyanate compounds [59,60]. Conversely, the “compound ratio” recognition mode, which predominates among parasitoids, involves the detection of widely occurring volatile compounds, with discrimination based on the specific compound ratios. Closterocerus ruforum (Krausse) (Hymenoptera: Eulophidae), a parasitoid of the common pine sawfly Diprion pini L. (Hymenoptera: Diprionidae), displays chemotaxis toward pine leaves oviposited by D. pini, which emit elevated levels of (E)-β-farnesene compared to uninfested controls. Further behavioral experiments revealed that single (E)-β-farnesene alone is insufficient to attract C. ruforum. Attraction occurs only when it is combined with five other terpenoids, i.e., β-phellandrene, (E)-β-ocimene, (Z)-β-ocimene, (E)-β-caryophyllene, and α-humulene, in ratios matching those emitted by oviposited pine leaves [61]. Similarly, the braconid C. vestalis is attracted to a blend of heptanal, α-pinene, sabinene, and (Z)-3-hexenyl acetate in proportions mirroring those released by Plutella xylostella L. (Lepidoptera: Plutellidae)-infested cabbages [62].
3.2. Host-Associated Chemical Cues
Stimuli used in habitat locations (i.e., long-range) are generally plant volatiles or herbivore pheromones, while stimuli used in host locations (i.e., short-range) are nonvolatile herbivore products [2,3,63]. Parasitoids primarily use HIPVs in long-range foraging [64]. Upon arrival at the herbivore-infested plant, they employed nonvolatile chemical cues from the herbivore hosts [6].
Stimuli derived from the herbivore host are generally the most reliable sources of information. Unfortunately, as herbivore insects are small elements in complex environments, concentrations of the produced chemicals are always tiny. Herbivores are often conspicuous to their parasitoids that exploit kairomones in host locations. For example, parasitoids of the bark beetle I. pini were attracted to the aggregation pheromone 2-methyl-6-methylene-2,7-octadien-4-ol of host insects [65]. Sex pheromones from certain moth species similarly attract parasitoids [66]. For instance, sex pheromones of the stem borer Sesamia nonagrioides (Lefèbvre) (Lepidoptera: Noctuidae) attract the egg parasitoid Telenomus busseolae Gahan (Hymenoptera: Scelionidae) [67]. Egg parasitoids of the genus Trichogramma (Hymenoptera) use sex pheromones of the adult hosts, such as sex pheromones of Manduca sexta L. (Lepidoptera: Sphingidae) [68].
Parasitoids can also use semiochemicals directing the stage of the herbivore. For example, Opius lectus Gahan (Hymenoptera: Braconidae), an egg-larval parasitoid of the apple maggot flies Rhagoletis pomonella (Walsh) (Diptera: Tephritidae), identifies the adult fly’s oviposition-deterring pheromone emitted from the fruit, while O. alloeus Mues, a parasitoid of late larval stages of R. pomonella, was not influenced by the pheromone [69]. Host larvae feces are also attractive to the parasitoids. Wind-tunnel assays showed that feces from the second-instar larvae of P. rapae L. strongly attract the parasitoid Cotesia rubecula (Marshall) (Hymenoptera: Braconidae) [70].
In addition to volatile chemicals, contact chemicals released by herbivore insects can also guide parasitoids to their hosts. Behavioral studies showed that the egg parasitoid T. basalis uses n-nonadecane, a cuticular hydrocarbon from the southern green stink bug N. viridula, to discriminate between female and male hosts [71,72]. Trissolcus brochymenae (Ashmead) (Hymenoptera: Scelionidae), an egg parasitoid of the brassicaceae pest Murgantia histrionica (Hahn), displayed an intense searching behavior in open arenas treated with contact semiochemicals from the host insects [73]. Polypeptides isolated from the host cocoon of Acrolepiopsis assectella (Zeller) (Lepidoptera: Acrolepiidae) was involved in host recognition behavior by the pupal parasitoid Diadromus pulchellus (Wsm) (Hymenoptera: Ichneumonidae) [74]. Probing–oviposition behavior of the parasitoid Eriborus terebrans Gravenhorst (Hymenoptera: Ichneumonidae) were readily stimulated in contact bioassays of the feces, oral secretion, and cuticles of Ostrinia nubilalis (Guenée) caterpillar [75], and sugars and amino acids in these compounds have been identified as semiochemicals in host location by the parasitoids [76,77]. Several other studies have found that volatiles originating from larval feces and sex pheromones from adult hosts can act together as semiochemicals, favoring host location of parasitoids [78,79,80].
Allomones are a type of allelochemical that elicit a behavioral or physiological response in the receiver that is beneficial to the sender [2,3]. These chemical signals are often used by organisms as a defense mechanism. Parasitoids could also be able to detect allomones. For example, defensive compounds such as (E)-2-decenal emitted from the metathoracic gland of Nezara viridula L. (Hemiptera: Pentatomidae) adults acts as a long-range kairomone orienting the egg parasitoid Trissolcus basalis (Wollaston) (Hymenoptera: Scelionidae) females to their hosts [81].
4. Semiochemicals Used by Hyperparasitoids
Hyperparasitoids are insects in the fourth trophic level that parasitize the larvae/pupae of primary parasitoids (the third trophic level) (Table 1). Hyperparasitoid larvae develop at the expense of parasitoids, which can greatly hamper the efficacy of parasitoids in biological control [82]. For example, in cotton fields in northern China, the hyperparasitoids Syrphophagus taeniatus (Förster) and S. aphidivorus (Mayr) (Hymenoptera: Encyrtidae) have been observed to ferociously attack the aphid parasitoids Lysiphlebus testaceipes (Cresson) (Hymenoptera: Aphidiidae) and Lipolexis scutellaris Mackauer (Hymenoptera: Braconidae), which host the cotton aphid species Toxoptera citricida (Kirkaldy) and Aphis gossypii Glover (Homoptera: Aphididae) [83].

Table 1.
Semiochemical sources and effects on hyperparasitoid species.
Over the past two decades, a few studies of the chemical cues exploited by hyperparasitoids in host locations have been documented [84,91,92,93]. Like their parasitoid hosts, hyperparasitoids use chemical information to locate their hosts (although other physical cues may be involved). HIPVs are found to influence behaviors of hyperparasitoids [85,94]. Chemical compositions of the HIPV blend change according to the parasitism status of the attacking herbivores, allowing hyperparasitoids to locate their host [84,86]. Lysibia nana (Gravenhorst) (Hymenoptera: Ichneumonidae), a specialized hyperparasitoid of the cocoons of genus Cotesia, prefers plant volatiles emitted by cabbage plants attacked by parasitized caterpillars than plant volatiles induced by unparasitized caterpillars [94]. Injection with polydnavirus and/or venom of the parasitoid Cotesia glomerata L. (Hymenoptera: Braconidae) affects the composition of oral secretions of herbivores and thereby changes the profile of volatiles emitted by the host plant, making these volatiles reliable indicators of host presence to L. nana [95]. Pteromalus semotus (Walker) (Hymenoptera: Pteromalidae), another hyperparasitoid of Cotesia parasitoids (Hymenoptera), is attracted by plant volatiles induced by parasitized herbivores [86].
Semiochemicals involved in the foraging behavior in several aphid-associated hyperparasitoid species have been extensively studied. Some species were shown to respond to HIPVs, while others have responses to aphid-associated chemical cues [87,96,97]. A study on four species of aphid–hyperparasitoids demonstrated that contact cues of parasitized aphids are vital in parasitoid locations [88]. Aphid honeydews may also emit semiochemicals that could be exploited by aphid–hyperparasitoids. Aphid–hyperparasitoids from Encyrtidae, Alloxystidae, and Megaspilidae show antennal drumming responses to substrates smeared with aphid honeydews [87,96].
During foraging at a close range, hyperparasitoids may also use information from their parasitoid host or the host insect of the parasitoids (the second trophic level). For example, the hyperparasitoid Baryscapus galactopus (Ratzeburg) (Hymenoptera: Eulophidae) uses body chemicals of the herbivore insect to discriminate parasitized P. rapae from unparasitized ones [96,97]. Analysis of volatiles from caterpillar bodies found that 2,3-butanedione was present in higher concentrations in the headspace of parasitized caterpillars [88]. Hyperparasitoids may also respond to cuticular hydrocarbons of the herbivore host. Alloxysta victrix (Westwood) (Hymenoptera: Charipinae) females have a longer dwelling time on spots with extracts of parasitized aphids compared with those of unparasitized aphids [88].
The information available for sex communication in hyperparasitoids is limited. In the aphid hyperparasitoid Dendrocerus carpenteri (Curtis) (Hymenoptera: Ceraphronidae), males were attracted by a sex pheromone released by conspecific females [89]. According to Y-tube olfactometer experiments, 6-Methyl-5-hepten-2-one, produced by the aphid hyperparasitoid A. victrix, acts as an intraspecific sex pheromone in sex communication [90].
5. Chemosensory Mechanisms of Parasitoids
5.1. Olfactory Mechanisms
While behavioral responses have been extensively studied, the olfactory mechanisms governing semiochemical perception in parasitoids remain poorly understood. However, genome and transcriptome analyses have characterized odorant-binding protein (OBP) repertoires in at least 11 parasitoid species, advancing our understanding of their roles in olfactory stimulus detection [98,99,100,101,102,103,104,105,106]. For example, seven MmedOBPs in Microplitis mediator Haliday (Hymenoptera: Braconidae) have been functionally characterized, revealing binding affinities to diverse odorants [107]. Notably, MmedOBP4 exhibits strong affinities to aromatic compounds, while MmedOBP4 and MmedOBP6 bind terpenoids, such as α-humulene, nerolidol, and limonene [107], suggesting their involvement in HIPV perception.
Odorant receptor (OR) genes have been increasingly identified in parasitoids by genome and/or transcriptome analyses [105,106,108,109,110]. For example, the braconid species M. mediator possess 169 OR genes [110], and the jewel wasp N. vitripennis has 301 OR genes [108]. Two hundred and eleven ORs were identified in the antennal transcriptome of C. chlorideae [111]. Although a link between the number of ORs and the complexity of chemical space is not immediate, the increased number of OR genes in parasitoid species may relate to the detection of a wide range of semiochemicals [8,25,111].
In the ichneumonid wasp C. chlorideae, seven ORs show female head-biased expression, likely linked to host-seeking behaviors [111]. Among these, CchlOR62 is exclusively and highly expressed in female antennae, and when expressed in Xenopus oocytes, it responds specifically to (Z)-jasmone, an attractant for female C. chlorideae [111]. RNAi combined with behavioral assays of Anastatus japonicus Ashmead (Hymenoptera: Eupelmidae), a parasitoid of Tessaratoma papillosa Drury (Hemiptera: Tessaratomidae), demonstrated that AjapOR35 detects the oviposition attractants (E)-α-farnesene and β-caryophyllene [112]. Recently, odorant receptor co-receptor (ORco) genes of four Drosophila (Diptera)-parasitoid species were cloned. ORco-deficient male parasitoids, but not females, lost their courtship abilities. Moreover, ORco-deficient female parasitoids showed impaired host-searching performance and decreased oviposition rates [113]. To date, only one study has characterized ORs involved in sex pheromone detection in parasitoids [37]. In C. chlorideae, expression of six male antennae-biased ORs in Drosophila T1 neurons revealed that CchlOR18 and CchlOR47 were tuned to female-derived sex pheromone components, tetradecanal and 2-heptadecanone, respectively. RNAi of these ORs individually or together abolished attraction of male parasitoids to these compounds [37].
SNMPs, homologs of the human CD36 fatty acid transport proteins, are critical for chemoreception, particularly pheromone detection in Drosophila and some lepidopteran species [114]. In M. mediator, two SNMP transcripts, MmedSNMP1 and MmedSNMP2, were identified, with MmedSNMP1 showing significantly higher transcript levels in antennal flagella in both sexes, as evidenced by reverse-transcription PCR and in situ hybridization [115]. IRs, close relatives of the ancient ionotropic glutamate receptors, detect acids and amines [116]. RNAseq of M. mediator antennae identified 17 IRs, most of which are enriched in adult antennae [117]. Despite these advances, most olfactory genes in parasitoids remain uncharacterized, hindering the understanding of the olfactory mechanisms in parasitoids.
5.2. Gustatory Mechanisms
Parasitoids mainly exploit volatile semiochemicals in the initial steps of the host location whereas contact semiochemicals become progressively more important when parasitoids are in close proximity to their host insects. Most of the studies on chemosensory mechanism of parasitoids have been focused on the detection of volatile chemicals (olfaction) [7]. In contrast, relatively few studies have focused on the role of contact chemoreception (gustation) [118]. In the last two decades, a few studies have been executed in this field, especially on egg parasitoids. Egg parasitoids have developed specialized strategies for successful foraging, based on the exploitation of chemical cues which are related to the host eggs [66,119]. In Trichogrammatidae and Platygastridae, the main host-related information exploited by female wasps are kairomones associated with the herbivore host, such as scales left on the leaves by females or chemicals left behind by herbivore hosts while walking on the plants; these associated compounds may be linked to gustatory perception in parasitoids [119,120,121,122]. Methanolic extracts obtained from male and female host epidermis elicited action potentials in taste neurons housed in antennal sensilla chaetica of the egg parasitoid T. brochymenae, indicating that these sensilla are involved in the perception of non-volatile host kairomones [73,123].
Gustatory receptors (GRs) are mainly expressed in taste organs and are associated with contact chemoreception [124]. Unfortunately, our knowledge about the GRs in parasitoids is still lacking. Research in this direction is currently limited to the identification of GR genes by transcriptomic and/or genomics analyses [108,125]. The jewel wasp N. vitripennis has 47 apparently intact GR genes in their genome. Two candidate sugar receptors, a DmGR43a (detect fructose in Drosophila melanogaster (Meigen) (Diptera: Drosophilidae)) orthologue, and several additional GR lineages have been identified, including significant gene subfamily expansions related to the 10 GRs found in the honey bee Apis mellifera L. (Hymenoptera: Apidae) [108].
6. Application of Semiochemicals in Pest Management
Application of semiochemicals regulating parasitoid behaviors in pest management has been mainly focused on HIPVs. Current evidence highlights three primary strategies for leveraging HIPVs in parasitoid-based strategies: (1) deploying devices containing attractive odorants to enhance the attractiveness of host plants to parasitoids; (2) using neighboring plants to mediate tritrophic interactions; and (3) cultivating new plant varieties engineered to emit elevated levels of key volatiles that manipulate insect behaviors [2,3,7,14,126,127].
Hanging devices dispensing attractive odorants has gained widespread attention over the past three decades due to its efficacy and operational simplicity. A typical example is MeSA [126,127]. Beyond its role as a plant defense hormone, MeSA is induced by certain sucking insects [128]. Commercially available MeSA dispensers can successfully attract beneficial insects, such as lacewings and other parasitoids, to strawberry fields [128]. Similarly, (Z)-jasmone, an HIPV emitted by plants damaged by caterpillars or aphids [41,43,48,111,128,129], enhances parasitoid attraction when applied to crops like tomatoes [130] or rice [131]. Our field studies showed that its application to tobacco plants greatly increases the parasitism rate of H. armigera by the ichneumonid C. chlorideae [57,111]. Additionally, DMNT has been proven to be an attractant for a variety of parasitoids targeting caterpillars on cotton, maize, pepper, and cowpea [41,43,132,133,134,135,136]. In field cage studies, DMNT shows a strong attractive effect for M. mediator [136]. As a GLV, (Z)-3-hexenyl acetate, produced by numerous plant species following mechanical or herbivore damages [39,137], attracts Trichogramma chilonis Ishii (Hymenoptera: Trichogrammatidae) and Cotesia plutellae Kurdjumov (Hymenoptera: Braconidae) [79] and increases parasitism of Lygus lineolaris Goeze (Hemiptera: Miridae) by Anaphes iole Girault (Hymenoptera: Mymaridae) in cotton fields [138].
The use of companion plants exemplifies the “push–pull” strategy [15], which simultaneously attracts parasitoids in the main crop and repels herbivore pests to the trap crop (Figure 1). This approach has been proven effective in various contexts [91,139,140,141]. In sub-Saharan African, wheat plants sprayed with low levels of (Z)-jasmone repel the grain aphid Sitobion avenae (Fabricius) (Homoptera: Aphididae) while attracting its parasitoids A. ervi [14,142,143]. Field plots of wheat were sprayed with (Z)-jasmone at a rate equivalent to 50 g per hectare (in four consecutive seasons in mid-May and early June), and aphid counts were conducted at weekly intervals. The researchers found that aphid infestations were significantly reduced in (Z)-jasmone-treated plots when compared to the control field [141]. Furthermore, this could also be explained by the “prime” effect. It has been reported that (Z)-jasmone could prime inducible production of defense compounds in intact plants, including wheat, maize, soybean, tomato, and pepper [144,145,146,147,148,149,150]. For example, the soybean Glycine max L. (Fabales: Fabaceae), after being treated with (Z)-jasmone, emits a blend of volatiles that attracts the stink bug egg parasitoid Telenomus podisi Ashmead (Hymenoptera: Scelionidae) [146].
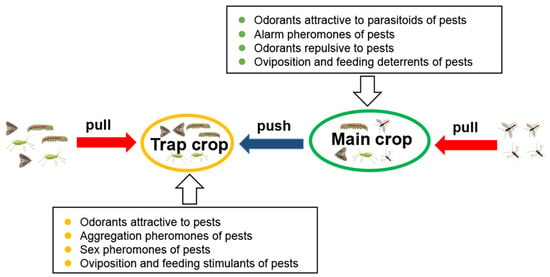
Figure 1.
Schematic of “push–pull” strategy applied in a tritrophic context. The push–pull strategy is an integrated pest management approach which leverages insect behaviors such as orientation, avoidance, feeding, mating, and oviposition by combining plant resources and artificial substances in main crops and trap crops. The “push” element primarily includes alarm pheromones, repellents, oviposition, and feeding deterrents. The “pull” element usually includes attractive host plant volatiles, sex pheromones, oviposition and feeding stimulants, and aggregation pheromones.
Cultivating plant varieties with enhanced volatile emission to attract parasitoids has been practically fulfilled with genetically modified plants. Typical evidence came from Arabidopsis thaliana (L.) (Brassicales: Brassicaceae) engineered to increase emissions of putative attractants, demonstrating enhanced parasitoid attraction in greenhouse assays [151]. Likewise, transgenic tobacco emitting elevated levels of the aphid alarm pheromone (E)-β-farnesene repels the green peach aphid Myzus persicae (Sulzer) (Hemiptera: Aphididae) while attracting its parasitoid D. rapae in behavioral bioassays [152,153]. Moreover, researchers have reported that supplemental feeding is useful to enhance the efficiency of parasitoids. For example, the parasitoid complex of the bark beetle Dendroctonus frontalis (Coleoptera: Scolytidae) in the southern United States consists of several species that may be important in regulating parasitism rate. Supplemental feedings of these parasitoids with an artificial diet have been shown to increase their longevity and fecundity [154]. This exemplifies the “feeding stimulants” component in the “pull” element.
Several studies tested the application of host-associated volatiles to enhance the efficacy of parasitoids. Among these, host sex pheromones and aggregation pheromones have been applied either as tools for monitoring parasitoid populations or for enhancing parasitism rate in the field. For example, a blend of the synthetic sex pheromone of Aphis fabae Scopoli (Hemiptera: Aphididae), (4aS,7S,7aR)-(+)-nepetalactone and (1R,4aS,7S,7aR)-(-)-nepetalactol could effectively attract three parasitoid species, i.e., Praon abjectum Haliday, P. dorsal Haliday, and P. volucre Haliday (Hymenoptera: Braconidae), to water traps [155]. In the Mediterranean basin, the sex pheromone of citrus mealybug Planococcus citri (Risso) and vine mealybug P. ficus (Hemiptera: Pseudococcidae), (S)-(+)-lavandulyl senecioate, applied in the vineyards, citrus or fig orchards could significantly increase the mealybug parasitism rate by Anagyrus sp. (Hymenoptera) [156]. In the soybean fields of South Korea, traps baited with the aggregation pheromone from Riptortus pedestris Fabricius (Hemiptera: Alydidae) could effectively attract its egg parasitoids, Ooencyrtus nezarae Ishii (Hymenoptera: Encyrtidae) and Gryon japonicum Ashmead (Hymenoptera: Scelionidae) [157]. Also, recent research identified that the synthetic compound (2,4,4-trimethyl-2-cyclohexenyl) methyl butyrate could attract two parasitoids, Anagyrus sawadai Ishii (Hymenoptera: Encyrtidae) and Leptomastix dactylopii Howard (Hymenoptera: Encyrtidae), which could effectively control the mealybug Planococcus kraunhiae Kuwana (Hemiptera: Pseudococcidae) [158,159]. Relevant semiochemicals for parasitoids cited in this review were summarized in Table 2.
Table 2.
Parasitoid species and elicited behaviors by semiochemicals (or blends).
A push–pull strategy could also be applied for the management of hyperparasitoids. Specific semiochemicals may be applied to push hyperparasitoids away (or pull them into traps), thus keeping primary parasitoids away from the attacks of hyperparasitoids [92,93]. HIPVs are of potential interest for push–pull strategies for hyperparasitoid management acted out at longer distances. Cabbage plants emit HIPVs in reply to infestation by parasitized herbivores, which can attract hyperparasitoids [5,6,84]. Garden experiments showed that the hyperparasitoid L. nana preferred plants that were induced by C. glomerata-parasitized Pieris brassicae L. (Lepidoptera: Pieridae) caterpillars [84]. Semiochemicals that act over a relatively short range mainly refer to pheromones. Pheromones could be directly applied to the dispensers, for example, using containers with parasitized insects smeared with the pheromones. The function of 6-methyl-5-hepten-2-one as a species-specific sex pheromone in A. victrix has been demonstrated [90,160]. However, this compound induces dispersal effect in the aphid parasitoid Aphidius uzbekistanicus Luzhetzki (Hymenoptera: Braconidae) [161]. Research on semiochemical applications at the fourth trophic level is still in its early stages compared to the extensively studied primary parasitoids. Increased efforts are required to explore semiochemical application at the fourth trophic level.
7. Future Perspectives
Enormous efforts have been made in the identification of semiochemicals used by insect parasitoids and hyperparasitoids in complex chemical environments and their application in insect pest management [2,6,7,14,162,163,164,165]. However, there are still several questions needing to be tackled in the future. First, a common issue is the discrepancy between field results and laboratory test results. The main reason is that the use of semiochemicals to manipulate parasitoids’ behavior is accomplished in environments that are complex. Hence, biotic factors, e.g., the recipient organisms (including plants and other insects, etc.) and their associated physiological and/or behavioral changes should be investigated. Besides this, the consequences of using specific chemical compounds in the field at the population level should be researched, such as population dynamics in different times and spaces [162,163]. Abiotic factors such as temperature and humidity should also be carefully considered. For example, the parasitism of Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae) decreases by nearly 40% at temperatures exceeding 35 °C [166]. Second, application of our knowledge into commercial products for pest management needs to be accelerated. This involves the progress of synthetic biology in the production of semiochemicals. By engineering plant or yeast metabolic pathways, synthetic biology enables these organisms for producing specific semiochemicals [167]. Compared to industrial synthetic chemicals, this method could realize the purpose of consistent production and release of semiochemicals in the field. Also, it can overcome limitations of other natural induction approaches, such as herbivore damage synthetic compounds. Third, the chemosensory mechanisms underlying semiochemical perception in parasitoids/hyperparasitoids remain largely unknown. Chemosensory proteins, especially ORs and GRs tuned to behavior-relevant semiochemicals in parasitoids/hyperparasitoids, await functional characterization. Furthermore, sex communication underlying sex pheromone identification and corresponding chemosensory mechanisms in parasitoid/hyperparasitoids needs to be unveiled. Systematic functional characterization of chemosensory receptors is essential to fully elucidate how parasitoids exploit chemical cues of varying reliability and detectability in host locations. Also, elucidation of chemosensory receptors could pave the way for “reverse chemical ecology”. For example, in the parasitoid C. chlorideae, functional studies of female antennae-prominent CchlOR62 demonstrated that this receptor responds robustly to the HIPV component (Z)-jasmone. Behavioral assays confirmed that (Z)-jasmone strongly attracts female parasitoids and significantly enhances their parasitism efficiency in early-instar H. armigera larvae [111]. Fortunately, the technique of studying insect chemosensory receptors is being updated gradually [168]. For example, calcium imaging (or patch clamp) of HEK293/Sf9 cells and the Xenopus (Anura) oocyte expression system (coupled with two-electrode voltage clamp recording) served as simple yet effective techniques for in vitro gene expression and functional characterization and have been extensively applied in studies of insect chemosensory receptors [169,170]. An in vivo expression system for deorphanization of insects ORs., i.e., the “empty neuron system” of Drosophila, has also been successfully established [28,168]. Furthermore, the recently emerged CRISPR/Cas9 gene-editing technology, distinguished by its operational simplicity and stable efficacy, has been successfully utilized in numerous investigations validating insect chemosensory functions [171,172]. We believe that with the accumulation of our knowledge and the advancement of techniques, the aforementioned issues will be resolved effectively and progressively. This will concretely facilitate the development of a practical way to use parasitoids as a resource to curb the explode of insect pests within agricultural ecosystems.
Author Contributions
Y.S.; funding acquisition, writing—original draft preparation. C.T.; funding acquisition, writing—original draft preparation. P.X.; investigation, writing—review and editing. J.D.; conceptualization, writing—review and editing. S.W.; conceptualization, supervision, validation. All authors have read and agreed to the published version of the manuscript.
Funding
Funding for this research was provided by the Open Project Program of Key Laboratory of Tabacco Pest Monitoring Controlling & Integrated Management (KLTPMIMT2022-19), the Beijing Agriculture Innovation Consortium (BAIC04-2025), the Independent Innovation Project of Henan Academy of Agricultural Sciences (2025ZC52), and the Henan province science and technology research and development plan joint fund (advantage discipline cultivation) project (242301420136).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhong, G.H.; Cui, G.F.; Yi, X.; Sun, R.R.; Zhang, J.J. Insecticide cytotoxicology in China: Current status and challenges. Pestic. Biochem. Physiol. 2016, 132, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Sobhy, I.S.; Erb, M.; Lou, Y.; Turlings, T.C.J. The prospect of applying chemical elicitors and plant strengtheners to enhance the biological control of crop pests. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20120283. [Google Scholar] [CrossRef] [PubMed]
- Wajnberg, E.; Desouhant, E. Editorial overview: Behavioral ecology: Behavioral ecology of insects: Current research and potential applications. Curr. Opin. Insect Sci. 2018, 27, 8–11. [Google Scholar] [CrossRef] [PubMed]
- You, L.S.; Lei, R.H.; Jiang, J.X.; Bo, L.Y.; Xiao, Z.S. Bionomic of Campoletis chlorideae (Hymenoptera: Ichneumonidae) as a parasitoid of the cotton bollworm Helicoverpa armigera (Lepidoptera: Noctuidae). Entomol. Sin. 2002, 9, 29–37. [Google Scholar]
- Poelman, E.H.; Cusumano, A.; de Boer, J.G. The ecology of hyperparasitoids. Annu. Rev. Entomol. 2022, 67, 143−161. [Google Scholar] [CrossRef]
- Aartsma, Y.; Bianchi, F.J.J.A.; van der Werf, W.; Poelman, E.H.; Dicke, M. Herbivore-induced plant volatiles and tritrophic interactions across spatial scales. New Phytol. 2017, 216, 1054–1063. [Google Scholar] [CrossRef]
- Turlings, T.C.J.; Erb, M. Tritrophic interactions mediated by herbivore induced plant volatiles: Mechanisms ecological relevance and application potential. Annu. Rev. Entomol. 2018, 63, 433−452. [Google Scholar] [CrossRef]
- Guo, H.; Wang, C.Z. The ethological significance and olfactory detection of herbivore-induced plant volatiles in interactions of plants herbivorous insects and parasitoids. Arthropod-Plant Interact. 2019, 13, 161–179. [Google Scholar] [CrossRef]
- Ali, M.Y.; Naseem, T.; Holopainen, J.K.; Liu, T.; Zhang, J.; Zhang, F. Tritrophic interactions among arthropod natural enemies’ herbivores and plants considering volatile blends at different scale levels. Cells 2023, 12, 251. [Google Scholar] [CrossRef]
- Steiner, S.; Ruther, J. Mechanism and behavioral context of male sex pheromone release in Nasonia vitripennis. J. Chem. Ecol. 2009, 35, 416−421. [Google Scholar] [CrossRef]
- Ruther, J.; Steiner, S.; Garbe, L.-A. 4-Methylquinazoline is a minor component of the male sex pheromone in Nasonia vitripennis. J. Chem. Ecol. 2008, 34, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Niehuis, O.; Buellesbach, J.; Gibson, J.D.; Pothmann, D.; Hanner, C.; Mutti, N.S.; Judson, A.K.; Gadau, J.; Ruther, J.; Schmitt, T. Behavioral and genetic analyses of Nasonia shed light on the evolution of sex pheromones. Nature 2013, 494, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Tamiru, A.; Bruce, T.J.; Woodcock, C.M.; Caulfield, J.C.; Midega, C.A.; Ogol, C.K.; Mayon, P.; Birkett, M.A.; Pickett, J.A.; Khan, Z.R. Maize landraces recruit egg and larval parasitoids in response to egg deposition by an herbivore. Ecol. Lett. 2011, 14, 1075–1083. [Google Scholar] [CrossRef]
- Pickett, J.A.; Khan, Z.R. Plant volatile-mediated signaling and its application in agriculture: Successes and challenges. New Phytol. 2016, 212, 856–870. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Midega, C.A.; Hooper, A.; Pickett, J. Push-Pull: Chemical ecology-based integrated pest management technology. J. Chem. Ecol. 2016, 42, 689−697. [Google Scholar] [CrossRef]
- Aartsma, Y.; Cusumano, A.; Fernández de Bobadilla, M.; Rusman, Q.; Vosteen, I.; Poelman, E.H. Understanding insect foraging in complex habitats by comparing trophic levels: Insights from specialist host-parasitoid-hyperparasitoid systems. Curr. Opin. Insect Sci. 2019, 32, 54−60. [Google Scholar] [CrossRef]
- Guo, H.; Wang, C.Z. Harnessing semiochemicals for parasitoid-based biological control: From laboratory identification to field applications. Crop Health 2025, 3, 21. [Google Scholar] [CrossRef]
- Hansson, B.S.; Stensmyr, M.C. Evolution of insect olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373−391. [Google Scholar] [CrossRef]
- Wäckers, F.L. Assessing the suitability of flowering herbs as parasitoid food sources: Flower attractiveness and nectar accessibility. Biol. Control 2004, 29, 307–314. [Google Scholar] [CrossRef]
- Jacob, H.S.; Evans, E.W. Influence of food deprivation on foraging decisions of the parasitoid Bathyplectes curculionis (Hymenoptera: Ichneumonidae). Ann. Entomol. Soc. Am. 2001, 94, 605–611. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Tollsten, L.; Bergström, L.G. Floral scents—A checklist of volatile compounds isolated by head-space techniques. Phytochem. 1993, 33, 253–280. [Google Scholar] [CrossRef]
- Borg Karlson, A.K.; Unelius, C.R.; Valterova, I.; Nilsson, L.A. Floral fragrance chemistry in the early flowering shrub Daphne mezereum. Phytochemistry 1995, 41, 1477–1483. [Google Scholar] [CrossRef]
- Du, Y.; Poppy, G.M.; Powell, W.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Identification of semiochemicals released during aphid feeding that attract parasitoid Aphidius ervi. J. Chem. Ecol. 1998, 24, 1355–1368. [Google Scholar] [CrossRef]
- Yan, Z.G.; Wang, C.Z. Identification of Mythimna separata-induced maize volatile synomones that attract the parasitoid Campoletis chlorideae. J. Appl. Entomol. 2006, 130, 213–219. [Google Scholar] [CrossRef]
- Salerno, G.; Iacovone, A.; Carlin, S.; Frati, F.; Conti, E.; Anfora, G. Identification of sex pheromone components in Trissolcus brochymenae females. J. Insect Physiol. 2012, 58, 1635–1642. [Google Scholar] [CrossRef]
- Ruther, J. Novel insights into pheromone-mediated communication in parasitic hymenopterans. In Chemical Ecology of Insect Parasitoids; Wajnberg, E., Colazza, S., Eds.; John Wiley & Sons: New York, NY, USA, 2013; pp. 112–144. [Google Scholar]
- Guo, H.; Huang, L.Q.; Wang, C.Z. Infochemicals used by parasitoids to find mates and hosts and their application in pest control. Chin. J. Appl. Entomol. 2023, 60, 375−388. [Google Scholar]
- Fauvergue, X.; Lo, G.A.; Lo, P.M. Virgins in the wild: Mating status affects the behavior of a parasitoid foraging in the field. Oecologia 2008, 156, 913–920. [Google Scholar] [CrossRef]
- Kant, R.; Minor, M.; Trewick, S. Mating or ovipositing? A crucial decision in the life history of the cabbage aphid parasitoid Diaeretiella rapae (M’Intosh). Ecol. Entomol. 2012, 37, 169–174. [Google Scholar] [CrossRef]
- Steiner, S.; Ruther, J. How important is sex for females of a haplodiploid species under local mate competition? Behav. Ecol. 2009, 20, 570–574. [Google Scholar] [CrossRef]
- Kugimiya, S.; Shimoda, T.; Wajnberg, E.; Uefune, M.; Takabayashi, J. Host-searching responses to herbivory-associated chemical information and patch use depend on mating status of female solitary parasitoid wasps. Ecol. Entomol. 2010, 35, 279–286. [Google Scholar] [CrossRef]
- Auguste, A.; Fauvergue, X. Intimate rendezvous in a tritrophic context? Nothing but the girls for male Lysiphlebus testaceipes. Ethology 2015, 121, 236–244. [Google Scholar] [CrossRef]
- Delury, N.C.; Gries, G.; Gries, R.; Judd, G.J.R.; Brown, J.J. Sex pheromone of Ascogaster quadridentata, a parasitoid of Cydia pomonella. J. Chem. Ecol. 1999, 25, 2229–2245. [Google Scholar] [CrossRef]
- Cônsoli, F.L.; Williams, H.J.; Vinson, S.B.; Matthews, R.W.; Cooperband, M.F. Trans-bergamotenes-male pheromone of the ectoparasitoid Melittobia digitata. J. Chem. Ecol. 2002, 28, 1675–1689. [Google Scholar] [CrossRef]
- Ruther, J.; Döring, M.; Steiner, S. Cuticular hydrocarbons as contact sex pheromone in the parasitoid Dibrachys cavus. Entomol. Exp. Appl. 2011, 140, 59–68. [Google Scholar] [CrossRef]
- Guo, H.; Mo, B.T.; Li, G.C.; Li, Z.L.; Huang, L.Q.; Sun, Y.L.; Dong, J.F.; Smith, D.P.; Wang, C.Z. Sex pheromone communication in an insect parasitoid Campoletis chlorideae Uchida. Proc. Natl. Acad. Sci. USA 2022, 119, e2215442119. [Google Scholar] [CrossRef]
- Burke, G.R.; Sharanowski, B.J. Parasitoid wasps. Curr. Biol. 2024, 10, R483−R488. [Google Scholar] [CrossRef]
- De Moraes, C.M.; Lewis, W.J.; Pare, P.W.; Alborn, H.T.; Tumlinson, J.H. Herbivore-infested plants selectively attract parasitoids. Nature 1998, 393, 570−573. [Google Scholar] [CrossRef]
- Dicke, M. Behavioural and community ecology of plants that cry for help. Plant Cell Environ. 2009, 32, 654−665. [Google Scholar] [CrossRef]
- Loughrin, J.H.; Manukian, A.; Heath, R.R.; Turlings, T.C.J.; Tumlinson, J.H. Diuenal cycle of emission of induced volatile terpenoids herbivore-injured cotton plants. Proc. Natl. Acad. Sci. USA 1994, 91, 11836−11840. [Google Scholar] [CrossRef]
- Turlings, T.C.J.; Loughrin, J.H.; McCall, P.J.; Röse, U.S.; Lewis, W.J.; Tumlinson, J.H. How caterpillar damaged plants protect themselves by attracting parasitic wasps. Proc. Natl. Acad. Sci. USA 1995, 92, 4169–4174. [Google Scholar] [CrossRef] [PubMed]
- Röse, U.S.R.; Tumlinson, J.H. Volatiles released from cotton plants in response to Helicoverpa zea feeding damage on cotton flower buds. Planta 2004, 218, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.G.; Wang, C.Z. Similar attractiveness of maize volatiles induced by Helicoverpa armigera and Pseudaletia separata to the generalist parasitoid Campoletis chlorideae. Entomol. Exp. Appl. 2006, 118, 87–96. [Google Scholar] [CrossRef]
- Sun, J.G.; Huang, L.Q.; Wang, C.Z. Electrophysiological and behavioral responses of Helicoverpa assulta (Lepidoptera: Noctuidae) to tobacco volatiles. Arthropod-Plant Interact. 2012, 6, 375–384. [Google Scholar] [CrossRef]
- Turlings, T.C.J.; Tumlinson, J.H.; Heath, R.R.; Proveaux, A.T.; Doolittle, R.E. Isolation and identification of allelochemicals that attract the larval parasitoid Cotesia marginiventris (Cresson) to the microhabitat of one of its hosts. J. Chem. Ecol. 1991, 17, 2235−2251. [Google Scholar] [CrossRef]
- Wu, H.; Li, R.T.; Dong, J.F.; Jiang, N.J.; Huang, L.Q.; Wang, C.Z. An odorant receptor and glomerulus responding to farnesene in Helicoverpa assulta (Lepidoptera: Noctuidae). Insect Biochem. Mol. Biol. 2019, 115, 103106. [Google Scholar] [CrossRef]
- Loughrin, J.H.; Manukian, A.; Heath, R.R.; Tumlinson, J.H. Volatiles emitted by different cotton varieties damaged by feeding beet armyworm larvae. J. Chem. Ecol. 1995, 21, 1217−1227. [Google Scholar] [CrossRef]
- Arimura, G.; Matsui, K.; Takabayashi, J. Chemical and molecular ecology of herbivore-induced plant volatiles: Proximate factors and their ultimate functions. Plant Cell Physiol. 2009, 50, 911–923. [Google Scholar] [CrossRef]
- Marmolejo, L.O.; Thompson, M.N.; Helms, A.M. Defense suppression through interplant communication depends on the attacking herbivore species. J. Chem. Ecol. 2021, 47, 1049–1061. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E.; Gershenzon, J. Biochemistry of plant volatiles. Plant Physiol. 2004, 135, 1893–1902. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef]
- ul Hassan, M.N.; Zainal, Z.; Ismail, I. Green leaf volatiles: Biosynthesis biological functions and their applications in biotechnology. Plant Biotechnol. J. 2015, 13, 727–739. [Google Scholar] [CrossRef]
- Maffei, M.E.; Gertsch, J.; Appendino, G. Plant volatiles: Production function and pharmacology. Nat. Prod. Rep. 2011, 28, 1359–1380. [Google Scholar] [CrossRef] [PubMed]
- Clavijo McCormick, A.; Gershenzon, J.; Unsicker, S.B. Little peaks with big effects: Establishing the role of minor plant volatiles in plant-insect interactions. Plant Cell Environ. 2014, 37, 1836–1844. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Jander, G. Molecular ecology of plant volatiles in interactions with insect herbivores. J. Exp. Bot. 2022, 73, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.L.; Dong, J.F.; Huang, L.Q.; Wang, C.Z. The cotton bollworm endoparasitoid Campoletis chlorideae is attracted by cis-jasmone or cis-3-hexenyl acetate but not by their mixtures. Arthropod-Plant Interact. 2020, 14, 169–179. [Google Scholar] [CrossRef]
- Gouinguené, S.; Pickett, J.A.; Wadhams, L.J.; Birkett, M.A.; Turlings, T.C.J. Antennal electrophysiological responses of three parasitic wasps to caterpillar-induced volatiles from maize (Zea mays mays) cotton (Gossypium herbaceum) and cowpea (Vigna unguiculata). J. Chem. Ecol. 2005, 31, 1023–1038. [Google Scholar] [CrossRef]
- Pope, T.W.; Kissen, R.; Grant, M.; Pickett, J.A.; Rossiter, J.T.; Powell, G. Comparative innate responses of the aphid parasitoid Diaeretiella rapae to alkenyl glucosinolate derived isothiocyanates nitriles and epithionitriles. J. Chem. Ecol. 2008, 34, 1302–1310. [Google Scholar] [CrossRef]
- Blande, J.D.; Pickett, J.A.; Poppy, G.M. A comparison of semiochemically mediated interactions involving specialist and generalist Brassica-feeding aphids and the braconid parasitoid Diaeretiella rapae. J. Chem. Ecol. 2007, 33, 767–779. [Google Scholar] [CrossRef]
- Beyaert, I.; Wäschke, N.; Scholz, A.; Varama, M.; Reinecke, A.; Hilker, M. Relevance of resource-indicating key volatiles and habitat odour for insect orientation. Anim. Behav. 2010, 79, 1077–1086. [Google Scholar] [CrossRef]
- Shiojiri, K.; Ozawa, R.; Kugimiya, S.; Uefune, M.; van Wijk, M.; Sabelis, M.W. Herbivore-specific density-dependent induction of plant volatiles: Honest or ‘cry wolf’ signals? PLoS ONE 2010, 5, e12161. [Google Scholar] [CrossRef] [PubMed]
- Plouvier, W.N.; Wajnberg, E. Improving the efficiency of augmentative biological control with arthropod natural enemies: A modeling approach. Biol. Control 2018, 125, 121−130. [Google Scholar] [CrossRef]
- van Oudenhove, L.; Mailleret, L.; Fauvergue, X. Infochemical use and dietary specialization in parasitoids: A meta-analysis. Ecol. Evol. 2017, 7, 4804−4811. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.R.; Borden, J.H. β-Phellandrene: Kairomone for pine engraver Ips pini (Say) (Coleoptera: Scolytidae). J. Chem. Ecol. 1990, 16, 2519−2531. [Google Scholar] [CrossRef]
- Fatouros, N.E.; Dicke, M.; Mumm, R.; Meiners, T.; Hilker, M. Foraging behavior of egg parasitoids exploiting chemical information. Behav. Ecol. 2008, 19, 677–689. [Google Scholar] [CrossRef]
- Colazza, S.; Rosi, M.C.; Clemente, A. Response of egg parasitoid Telenomus busseolae to sex pheromone of Sesamia nonagrioides. J. Chem. Ecol. 1997, 23, 2437−2444. [Google Scholar] [CrossRef]
- Noldus, L.P.J.J. Semiochemicals foraging behaviour and quality of entomophagous insects for biological control. J. Appl. Entomol. 1989, 108, 425−451. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Webster, R.P. Oviposition-deterring pheromone of Rhagoletis pomonella: A kairomone for its parasitoid Opius lectus. J. Chem. Ecol. 1978, 4, 481−494. [Google Scholar] [CrossRef]
- Agelopoulos, N.G.; Dicke, M.; Posthumus, M.A. Role of volatile infochemicals emitted by feces of larvae in host-searching behavior of parasitoid Cotesia rubecula (Hymenoptera: Braconidae): A behavioral and chemical study. J. Chem. Ecol. 1995, 21, 1789−1811. [Google Scholar] [CrossRef]
- Colazza, S.; Aquila, G.; De Pasquale, C.; Peri, E.; Millar, J.G. The egg parasitoid Trissolcus basalis uses n-nonadecane a cuticular hydrocarbon from its stink bug host Nezara viridula to discriminate between female and male hosts. J. Chem. Ecol. 2007, 33, 1405–1420. [Google Scholar] [CrossRef]
- Lo Giudice, D.; Riedel, M.; Rostás, M.; Peri, E.; Colazza, S. Host sex discrimination by an egg parasitoid on brassica leaves. J. Chem. Ecol. 2011, 37, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Iacovone, A.; French, A.S.; Tellier, F.; Cusumano, A.; Clément, G.; Gaertner, C.; Conti, E.; Salerno, G.; Marion-Poll, F. The role of contact chemoreception in the host location process of an egg parasitoid. J. Insect Physiol. 2016, 91−92, 63−75. [Google Scholar] [CrossRef] [PubMed]
- Bénédet, F.; Bigot, Y.; Renault, S.; Pouzat, J.; Thibout, E. Polypeptides of Acrolepiopsis assectella cocoon (Lepidoptera: Yponomeutoidea): An external host-acceptance kairomone for the parasitoid Diadromus pulchellus (Hymenoptera: Ichneumonidae). J. Insect Physiol. 1999, 45, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.Z.; Swedenborg, P.D.; Jones, R.L. Host-seeking behavior of Eriborus terebrans (Hymenoptera: Ichneumonidae) toward the European corn borer and the role of chemical stimuli. Ann. Entomol. Soc. Am. 1992, 85, 72–79. [Google Scholar] [CrossRef]
- Heath, R.R.; Ferkovich, S.M.; Greany, P.D.; Eller, F.J.; Dueben, B.D.; Tilden, R.L. Progress in the isolation and characterization of a host hemolymph ovipositional kairomone for the endoparasitoid Microplitis croceipes. Arch. Insect Biochem. Physiol. 1990, 13, 255–265. [Google Scholar] [CrossRef]
- Rutledge, C.E. A survey of identified kairomones and synomones used by insect parasitoids to locate and accept their hosts. Chemoecology 1996, 7, 121–131. [Google Scholar] [CrossRef]
- Metzger, M.; Fischbein, D.; Auguste, A.; Fauvergue, X.; Bernstein, C.; Desouhant, E. Synergy in information use for mate finding: Demonstration in a parasitoid wasp. Anim. Behav. 2010, 79, 1307–1315. [Google Scholar] [CrossRef]
- Reddy, G.V.; Holopainen, J.K.; Guerrero, A. Olfactory responses of Plutella xylostella natural enemies to host pheromone larval frass and green leaf cabbage volatiles. J. Chem. Ecol. 2002, 28, 131−143. [Google Scholar] [CrossRef]
- Steiner, S.; Steidle, J.L.M.; Ruther, J. Host-associated kairomones used for habitat orientation in the parasitoid Lariophagus distinguendus (Hymenoptera: Pteromalidae). J. Stored Prod. Res. 2007, 43, 587–593. [Google Scholar] [CrossRef]
- Mattiacci, L.; Vinson, S.B.; Williams, H.J. A long-range attractant kairomone for egg parasitoid Trissolcus basalis, isolated from defensive secretion of its host, Nezara viridula. J. Chem. Ecol. 1993, 19, 1167–1181. [Google Scholar] [CrossRef]
- Buitenhuis, R.; Vet, L.E.M.; Boivin, G.; Brodeur, J. Foraging behaviour at the fourth trophic level: A comparative study of host location in aphid hyperparasitoids. Entomol. Exp. Appl. 2005, 114, 107–117. [Google Scholar] [CrossRef]
- Zhu, Y.L.; Yang, F.; Yao, Z.W.; Wu, Y.K.; Liu, B.; Lu, Y.H. A molecular detection approach for a cotton aphid-parasitoid complex in northern China. Sci. Rep. 2019, 9, 15836. [Google Scholar] [CrossRef] [PubMed]
- Poelman, E.H.; Bruinsma, M.; Zhu, F.; Weldegergis, B.T.; Boursault, A.E.; Jongema, Y.; van Loon, J.J.; Vet, L.E.; Harvey, J.A.; Dicke, M. Hyperparasitoids use herbivore-induced plant volatiles to locate their parasitoid. PLoS Biol. 2012, 10, e1001435. [Google Scholar] [CrossRef] [PubMed]
- Poelman, E.H.; Harvey, J.A.; van Loon, J.J.A.; Vet, L.E.M.; Dicke, M. Variation in herbivore-induced plant volatiles corresponds with spatial heterogeneity in the level of parasitoid competition and parasitoid exposure to hyperparasitism. Funct. Ecol. 2013, 27, 1107–1116. [Google Scholar] [CrossRef]
- Zhu, F.; Cusumano, A.; Bloem, J.; Weldegergis, B.T.; Villela, A.; Fatouros, N.E.; van Loon, J.J.; Dicke, M.; Harvey, J.A.; Vogel, H.; et al. Symbiotic polydnavirus and venom reveal parasitoid to its hyperparasitoids. Proc. Natl. Acad. Sci. USA 2018, 115, 5205–5210. [Google Scholar] [CrossRef]
- Buitenhuis, R.; McNeil, J.; Boivin, G.; Brodeur, J. The role of honeydew in host searching of aphid hyperparasitoids. J. Chem. Ecol. 2004, 30, 273−285. [Google Scholar] [CrossRef]
- Grasswitz, T.R. Contact kairomones mediating the foraging behavior of the aphid hyperparasitoid Alloxysta victrix (Westwood) (Hymenoptera: Charipidae). J. Insect Behav. 1998, 11, 539–548. [Google Scholar] [CrossRef]
- Schworer, U.; Völkl, W.; Hoffmann, K.H. Foraging for mates in the hyperparasitic wasp Dendrocerus carpenteri: Impact of unfavourable weather conditions and parasitoid age. Oecologia 1999, 119, 73–80. [Google Scholar] [CrossRef]
- Micha, S.G.; Stammel, J.; Höller, C. 6-Methyl-5-heptene-2-one, a putative sex and spacing pheromone of the aphid hyperparasitoid Alloxysta victrix (Hymenoptera: Alloxystidae). Eur. J. Entomol. 1993, 90, 439–442. [Google Scholar]
- Kaplan, I. Attracting carnivorous arthropods with plant volatiles: The future of biocontrol or playing with fire? Biol. Control 2012, 60, 77–89. [Google Scholar] [CrossRef]
- Tougeron, K.; Tena, A. Hyperparasitoids as new targets in biological control in a global change context. Biol. Control 2018, 130, 164–167. [Google Scholar] [CrossRef]
- Cusumano, A.; Harvey, J.A.; Bourne, M.E.; Poelman, E.H.; de Boer, J.G. Exploiting chemical ecology to manage hyperparasitoids in biological control of arthropod pests. Pest Manag. Sci. 2020, 76, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Poelman, E.H.; Kos, M. Complexity of plant volatile-mediated interactions beyond the third trophic level. In Deciphering Chemical Language of Plant Communication; Blande, J.D., Glinwood, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 211–225. [Google Scholar]
- Budenberg, W. Honeydew as a contact kairomone for aphid parasitoids. Entomol. Exp. Appl. 1990, 55, 139−148. [Google Scholar] [CrossRef]
- Völkl, W.; Sullivan, D.J. Foraging behaviour, host plant and host location in the aphid hyperparasitoid Euneura augarus. Entomol. Exp. Appl. 2000, 97, 47–56. [Google Scholar] [CrossRef]
- Zhu, F.; Weldegergis, B.T.; Lhie, B.; Harvey, J.A.; Dicke, M.; Poelman, E.H. Body odors of parasitized caterpillars give away the presence of parasitoid larvae to their primary hyperparasitoid enemies. J. Chem. Ecol. 2014, 40, 986–995. [Google Scholar] [CrossRef]
- Vieira, F.G.; Forêt, S.; He, X.; Rozas, J.; Field, L.M.; Zhou, J.J. Unique features of odorant-binding proteins of the parasitoid wasp Nasonia vitripennis revealed by genome annotation and comparative analyses. PLoS ONE 2012, 7, e43034. [Google Scholar] [CrossRef]
- Nishimura, O.; Brillada, C.; Yazawa, S.; Maffei, M.E.; Arimura, G. Transcriptome pyrosequencing of the parasitoid wasp Cotesia vestalis: Genes involved in the antennal odorant-sensory system. PLoS ONE 2012, 7, e50664. [Google Scholar] [CrossRef]
- Wang, J.; Li, D.Z.; Min, S.F.; Mi, F.; Zhou, S.S.; Wang, M.Q. Analysis of chemosensory gene families in the beetle Monochamus alternatus and its parasitoid Dastarcus helophoroides. Comp. Biochem. Physiol. Part D Genom. Proteom. 2014, 11, 1–8. [Google Scholar] [CrossRef]
- Wang, S.N.; Peng, Y.; Lu, Z.Y.; Dhiloo, K.H.; Gu, S.H.; Li, R.J. Identification and expression analysis of putative chemosensory receptor genes in Microplitis mediator by antennal transcriptome screening. Int. J. Biol. Sci. 2015, 11, 737–751. [Google Scholar] [CrossRef]
- Wang, S.N.; Shan, S.; Liu, J.T.; Li, R.J.; Lu, Z.Y.; Dhiloo, K.H.; Khashaveh, A.; Zhang, Y.J. Characterization of antennal chemosensilla and associated odorant binding as well as chemosensory proteins in the parasitoid wasp Microplitis mediator (Hymenoptera: Braconidae). Sci. Rep. 2018, 8, 7649. [Google Scholar] [CrossRef]
- Farias, L.R.; Schimmelpfeng, P.H.; Togawa, R.C.; Costa, M.M.; Grynberg, P.; Martins, N.F.; Borges, M.; Blassioli-Moraes, M.C.; Laumann, R.A.; Bao, S.N.; et al. Transcriptome-based identification of highly similar odorant-binding proteins among neotropical stink bugs and their egg parasitoid. PLoS ONE 2015, 10, e0132286. [Google Scholar] [CrossRef]
- Li, K.; Yang, X.; Xu, G.; Cao, Y.; Lu, B.; Peng, Z. Identification of putative odorant binding protein genes in Asecodes hispinarum, a parasitoid of coconut leaf beetle (Brontispa longissima) by antennal RNA-Seq analysis. Biochem. Biophys. Res. Commun. 2015, 467, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Sheng, S.; Liao, C.W.; Zheng, Y.; Zhou, Y.; Xu, Y.; Song, W.M.; He, P.; Zhang, J.; Wu, F.A. Candidate chemosensory genes identified in the endoparasitoid Meteorus pulchricornis (Hymenoptera: Braconidae) by antennal transcriptome analysis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 22, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.B.; Wu, H.; Yi, J.Q.; Song, Z.W.; Li, D.S.; Zhang, G.R. Transcriptome characterization and gene expression analysis related to chemoreception in Trichogramma chilonis, an egg parasitoid. Gene 2018, 678, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, L.Z.; Gu, S.H.; Cui, J.J.; Gao, X.W.; Zhang, Y.J.; Guo, Y.Y. Binding characterization of recombinant odorant-binding proteins from the parasitic Wasp Microplitis mediator (Hymenoptera: Braconidae). J. Chem. Ecol. 2011, 37, 189–194. [Google Scholar] [CrossRef]
- Robertson, H.M.; Gadau, J.; Wanner, K.W. The insect chemoreceptor superfamily of the parasitoid jewel wasp Nasonia vitripennis. Insect Mol. Biol. 2010, 19, 121−136. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, Y.; Li, D.; Fan, Y. Transcriptomics and identification of the chemoreceptor superfamily of the pupal parasitoid of the oriental fruit fly Spalangia endius Walker (Hymenoptera: Pteromalidae). PLoS ONE 2014, 9, e87800. [Google Scholar] [CrossRef]
- Wang, S.N.; Shan, S.; Zheng, Y.; Peng, Y.; Lu, Z.Y.; Yang, Y.Q.; Li, R.J.; Zhang, Y.J.; Guo, Y.Y. Gene structure and expression characteristic of a novel odorant receptor gene cluster in the parasitoid wasp Microplitis mediator (Hymenoptera: Braconidae). Insect Mol. Biol. 2017, 26, 420–431. [Google Scholar] [CrossRef]
- Sun, Y.L.; Dong, J.F.; Ning, C.; Ding, P.P.; Huang, L.Q.; Sun, J.G.; Wang, C.Z. An odorant receptor mediates the attractiveness of cis-jasmone to Campoletis chlorideae, the endoparasitoid of Helicoverpa armigera. Insect Mol. Biol. 2019, 12, 23−34. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Q.; Guo, J.; Li, J.; Wang, J.; Wen, M.; Zhao, H.; Ren, B. Molecular basis of peripheral olfactory sensing during oviposition in the behavior of the parasitic wasp Anastatus japonicus. Insect Biochem. Mol. Biol. 2017, 89, 58–70. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, J.; Wang, Y.; Lu, Y.; Dong, Z.; Shi, W.; Pang, L.; Ren, S.; Chen, X.; Huang, J. The odorant receptor co-receptor gene contributes to mating and host-searching behaviors in parasitoid wasps. Pest Manag. Sci. 2023, 79, 454−463. [Google Scholar] [CrossRef]
- Rogers, M.E.; Sun, M.; Lerner, M.R.; Vogt, R.G. SNMP-1, a novel membrane protein of olfactory neurons of the silk moth Antheraea polyphemus with homology to the CD36 family of membrane proteins. J. Biol. Chem. 1997, 272, 14792–14799. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.; Wang, S.; Song, X.; Khashaveh, A.; Lu, Z.; Dhiloo, K.H.; Li, R.; Gao, X.; Zhang, Y. Molecular characterization and expression of sensory neuron membrane proteins in the parasitoid Microplitis mediator (Hymenoptera: Braconidae). Insect Sci. 2020, 27, 425−439. [Google Scholar] [CrossRef] [PubMed]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-N.; Peng, Y.; Lu, Z.-Y.; Dhiloo, K.H.; Zheng, Y.; Shan, S.; Li, R.-J.; Zhang, Y.-J.; Guo, Y.-Y. Cloning and expression profile of ionotropic receptors in the parasitoid wasp Microplitis mediator (Hymenoptera: Braconidae). J. Insect Physiol. 2016, 90, 27−35. [Google Scholar] [CrossRef]
- van Baaren, J.; Boivin, G.; Bourdais, D.; Roux, O. Antennal sensilla of hymenopteran parasitic wasps: Variations linked to host exploitation behavior. In Modern Research and Educational Topics in Microscopy; Vilas, A.M., Alvarez, J.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 345–352. [Google Scholar]
- Colazza, S.; Peri, E.; Salerno, G.; Conti, E. Host searching by egg parasitoids: Exploitation of host chemical cues. In Egg Parasitoids in Agroecosystems with Emphasis on Trichogramma; Cônsoli, F.L., Parra, J.R.P., Zucchi, R.A., Eds.; Springer: London, UK, 2010; pp. 97–147. [Google Scholar]
- Borges, M.; Colazza, S.; Ramirez-Lucas, P.; Chauhan, K.R.; Moraes, M.C.B.; Aldrich, J.R. Kairomonal effect of walking traces from Euschistus heros (Heteroptera: Pentatomidae) on two strains of Telenomus podisi (Hymenoptera: Scelionidae). Physiol. Entomol. 2003, 28, 49–355. [Google Scholar] [CrossRef]
- Conti, E.; Salerno, G.; Bin, F.; Williams, H.J.; Vinson, S.B. Chemical cues from Murgantia histrionica eliciting host location and recognition in the egg parasitoid Trissolcus brochymenae. J. Chem. Ecol. 2003, 29, 115–130. [Google Scholar] [CrossRef]
- Salerno, G.; Conti, E.; Peri, E.; Colazza, S.; Bin, F. Kairomone involvement in the host specificity of the egg parasitoid Trissolcus basalis (Hymenoptera: Scelionidae). Eur. J. Entomol. 2006, 103, 311–318. [Google Scholar] [CrossRef]
- Iacovone, A.; Salerno, G.; French, A.S.; Conti, E.; Marion-Poll, F. Antennal gustatory perception and behavioral responses in Trissolcus brochymenae females. J. Insect Physiol. 2015, 78, 15–25. [Google Scholar] [CrossRef]
- Clyne, P.J.; Warr, C.G.; Freeman, M.R.; Lessing, D.; Kim, J.H.; Carlson, J.R. A novel family of divergent seven-transmembrane proteins: Candidate odorant receptors in Drosophila. Neuron 1999, 22, 327−338. [Google Scholar] [CrossRef]
- Ruschioni, S.; van Loon, J.J.A.; Smid, H.M.; van Lenteren, J.C. Insects can count: Sensory basis of host discrimination in parasitoid wasps revealed. PLoS ONE 2015, 10, e0138045. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C. Effect of methyl salicylate-based lures on beneficial and pest arthropods in strawberry. Environ. Entomol. 2010, 39, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Saona, C.; Kaplan, I.; Braasch, J.; Chinnasamy, D.; Williams, L. Field responses of predaceous arthropods to methyl salicylate: A meta-analysis and case study in cranberries. Biol. Control 2011, 59, 294–303. [Google Scholar] [CrossRef]
- Zhang, P.; Xu, C.; Zhang, J.; Lu, Y.; Wei, J.; Liu, Y.; David, A.; Boland, W.; Turlings, T.C.J. Phloem-feeding whiteflies can fool their host plants but not their parasitoids. Funct. Ecol. 2013, 27, 1304–1312. [Google Scholar] [CrossRef]
- Lou, Y.G.; Baldwin, I.T. Manduca sexta recognition and resistance among allopolyploid Nicotiana host plants. Proc. Natl. Acad. Sci. USA 2003, 100, 14581−14586. [Google Scholar] [CrossRef]
- Thaler, J.S. Jasmonate-inducible plant defences cause increased parasitism of herbivores. Nature 1999, 399, 686–688. [Google Scholar] [CrossRef]
- Lou, Y.G.; Du, M.H.; Turlings, T.C.; Cheng, J.A.; Shan, W.F. Exogenous application of jasmonic acid induces volatile emissions in rice and enhances parasitism of Nilaparvata lugens eggs by the parasitoid Anagrus nilaparvatae. J. Chem. Ecol. 2005, 31, 1985−2002. [Google Scholar] [CrossRef]
- McCall, P.J.; Turlings, T.C.J.; Loughrin, J.; Proveaux, A.T.; Tumlinson, J.H. Herbivore-induced volatile emissions from cotton (Gossypium hirsutum L.) seedlings. J. Chem. Ecol. 1994, 20, 3039–3050. [Google Scholar] [CrossRef]
- Röse, U.S.R.; Manukian, A.; Heath, R.R.; Tumlinson, J.H. Volatile semiochemicals released from undamaged cotton leaves (A systemic response of living plants to caterpillar damage). Plant Physiol. 1996, 111, 487–495. [Google Scholar] [CrossRef]
- Kappers, I.F.; Aharoni, A.; van Herpen, T.W.; Luckerhoff, L.L.; Dicke, M.; Bouwmeester, H.J. Genetic engineering of terpenoid metabolism attracts bodyguards to Arabidopsis. Science 2005, 309, 2070–2072. [Google Scholar] [CrossRef]
- D’Alessandro, M.; Brunner, V.; von Mérey, G.; Turlings, T.C.J. Strong attraction of the parasitoid Cotesia marginiventris towards minor volatile compounds of maize. J. Chem. Ecol. 2009, 35, 999. [Google Scholar] [CrossRef][Green Version]
- Yu, H.L.; Zhang, Y.J.; Wyckhuys, K.A.G.; Wu, K.M.; Gao, X.W.; Wu, K.M. Electrophysiological and behavioral Responses of Microplitis mediator (Hymenoptera: Braconidae) to caterpillar-induced volatiles from cotton. Environ. Entomol. 2010, 39, 600–609. [Google Scholar] [CrossRef]
- Clavijo McCormick, A.; Unsicker, S.B.; Gershenzon, J. The specificity of herbivore-induced plant volatiles in attracting herbivore enemies. Trends Plant Sci. 2012, 17, 303−310. [Google Scholar] [CrossRef] [PubMed]
- Williams, L., 3rd; Rodriguez-Saona, C.; Castle, S.C.; Zhu, S. EAG active herbivore-induced plant volatiles modify behavioral responses and host attack by an egg parasitoid. J. Chem. Ecol. 2008, 34, 1190–1201. [Google Scholar] [CrossRef] [PubMed]
- James, D.G. Synthetic herbivore-induced plant volatiles as field attractants for beneficial insects. Environ. Entomol. 2003, 32, 977–982. [Google Scholar] [CrossRef]
- James, D.G.; Grasswitz, T.R. Synthetic herbivore-induced plant volatiles increase field captures of parasitic wasps. BioControl 2005, 50, 871–880. [Google Scholar] [CrossRef]
- Uefune, M.; Choh, Y.; Abe, J.; Shiojiri, K.; Sano, K.; Takabayashi, J. Application of synthetic herbivore-induced plant volatiles causes increased parasitism of herbivores in the field. J. Appl. Entomol. 2012, 136, 561–567. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Pickett, J.A.; Smart, L.E. cis-Jasmone switches on plant defence against insects. Pestic. Outlook 2003, 3, 96−98. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Martin, J.L.; Pickett, J.A.; Pye, B.J.; Smart, L.E.; Wadhams, L.J. cis-Jasmone treatment induces resistance in wheat plants against the grain aphid Sitobion avenae (Fabricius) (Homoptera: Aphididae). Pest Manag. Sci. 2003, 59, 1031−1036. [Google Scholar] [CrossRef]
- Pickett, J.A.; Birkett, M.A.; Bruce, T.J.; Chamberlain, K.; Gordon-Weeks, R.; Matthes, M.C.; Napier, J.A.; Smart, L.E.; Woodcock, C.M. Developments in aspects of ecological phytochemistry: The role of cis-jasmone in inducible defence systems in plants. Phytochemistry 2007, 68, 2937–2945. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Matthes, M.C.; Chamberlain, K.; Woodcock, C.M.; Mohib, A.; Webster, B.; Smart, L.E.; Birkett, M.A.; Pickett, J.A.; Napier, J.A. cis-Jasmone induces Arabidopsis genes that affect the chemical ecology of multitrophic interactions with aphids and their parasitoids. Proc. Natl. Acad. Sci. USA 2008, 105, 4553–4558. [Google Scholar] [CrossRef] [PubMed]
- Moraes, M.C.B.; Laumann, R.A.; Pareja, M.; Sereno, F.T.P.S.; Michereff, M.F.F.; Birkett, M.A.; Pickett, J.A.; Borges, M. Attraction of the stink bug egg parasitoid Telenomus podisi to defence signals from soybean activated by treatment with cis-jasmone. Entomol. Exp. Appl. 2009, 131, 178–188. [Google Scholar] [CrossRef]
- Dewhirst, S.Y.; A Birkett, M.; Loza-Reyes, E.; Martin, J.L.; Pye, B.J.; E Smart, L.; Hardie, J.; A Pickett, J. Activation of defence in sweet pepper, Capsicum annum, by cis-jasmone, and its impact on aphid and aphid parasitoid behavior. Pest Manag. Sci. 2012, 68, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Oluwafemi, S.; Dewhirst, S.Y.; Veyrat, N.; Powers, S.; Bruce, T.J.A.; Caulfield, J.C.; Pickett, J.A.; Birkett, M.A. Priming of production in maize of volatile organic defence compounds by the natural plant activator cis-Jasmone. PLoS ONE 2013, 8, e62299. [Google Scholar] [CrossRef]
- Disi, J.O.; Zebelo, S.; Ngumbi, E.; Fadamiro, H.Y. cis-Jasmone primes defense pathways in tomato via emission of volatile organic compounds and regulation of genes with consequences for Spodoptera exigua oviposition. Arthropod-Plant Interact. 2017, 11, 591–602. [Google Scholar] [CrossRef]
- Sobhy, I.S.; Woodcock, C.M.; Powers, S.J.; Caulfiled, J.C.; Pickett, J.A.; Birkett, M.A. cis-Jasmone elicits aphid-induced stress signaling in potatoes. J. Chem. Ecol. 2017, 43, 39–52. [Google Scholar] [CrossRef]
- Schnee, C.; Kollner, T.G.; Held, M.; Turlings, T.C.J.; Gershenzon, J.; Degenhardt, J. The products of a single maize sesquiterpene synthase form a volatile defense signal that attracts natural enemies of maize herbivores. Proc. Natl. Acad. Sci. USA 2006, 103, 1129–1134. [Google Scholar] [CrossRef]
- Foster, S.P.; Denholm, I.; Thompson, R.; Poppy, G.M.; Powell, W. Reduced response of insecticide-resistant aphids and attraction of parasitoids to aphid alarm pheromone; a potential fitness trade-off. Bull. Entomol. Res. 2005, 95, 37–46. [Google Scholar] [CrossRef]
- Verheggen, F.J.; Arnaud, L.; Bartram, S.; Gohy, M.; Haubruge, E. Aphid and plant volatiles induce oviposition in an aphidophagous hoverfly. J. Chem. Ecol. 2008, 34, 301–307. [Google Scholar] [CrossRef]
- Stephen, F.M.; Browne, L.E. Application of Eliminade™ parasitoid food to boles and crowns of pines (Pinaceae) infested with Dendroctonus frontalis (Coleoptera: Scolytidae). Can. Ent. 2000, 132, 983−985. [Google Scholar] [CrossRef]
- Hardie, J.; Nottingham, S.F.; Powell, W.; Wadhams, L.J. Synthetic aphid sex pheromone lures female parasitoids. Entomol. Exp. Appl. 1991, 61, 97–99. [Google Scholar] [CrossRef]
- Franco, J.C.; da Silva, E.B.; Fortuna, T.; Cortegano, E.; Branco, M.; Suma, P.; La Torre, I.; Russo, A.; Elyahu, M.; Protasov, A.; et al. Vine mealybug sex pheromone increases citrus mealybug parasitism by Anagyrus sp. near pseudococci (Girault). Biol. Control 2011, 58, 230–238. [Google Scholar] [CrossRef]
- Lim, U.T.; Mainali, B.P. Effect of aggregation pheromone trap of Riptortus pedestris (Hemiptera: Alydidae) on the distribution and composition of its egg parasitoids. J. Econ. Entomol. 2013, 106, 1973–1978. [Google Scholar] [CrossRef] [PubMed]
- Teshiba, M.; Sugie, H.; Tsutsumi, T.; Tabata, J. A new approach for mealybug management: Recruiting an indigenous, but ‘non-natural’ enemy for biological control using an attractant. Entomol. Exp. Appl. 2012, 142, 211–215. [Google Scholar] [CrossRef]
- Teshiba, M.; Tabata, J. Suppression of population growth of the Japanese mealybug, Planococcus kraunhiae (Hemiptera: Pseudococcidae), by using an attractant for indigenous parasitoids in persimmon orchards. Appl. Entomol. Zool. 2017, 52, 153–158. [Google Scholar] [CrossRef]
- Völkl, W.; Hübner, G.; Dettner, K. Interactions between Alloxysta brevis (Hymenoptera, Cynipoidea, Alloxystidae) and honeydew-collecting ants: How an aphid-hyperparasitoid overcomes ant aggression by chemical defense. J. Chem. Ecol. 1994, 20, 2901–2915. [Google Scholar] [CrossRef]
- Höller, C.; Micha, S.G.; Schulz, S.; Francke, W.; Pickett, J.A. Enemy-induced dispersal in a parasitic wasp. Experientia 1994, 50, 182–185. [Google Scholar] [CrossRef]
- Lewis, W.J.; Martin, W.R. Semiochemicals for use with parasitoids: Status and future. J. Chem. Ecol. 1990, 16, 3067–3089. [Google Scholar] [CrossRef]
- Meinwald, J.; Eisner, T. Chemical ecology in retrospect and prospect. Proc. Natl. Acad. Sci. USA 2008, 105, 4539–4540. [Google Scholar] [CrossRef]
- Jiang, Y.; Xiu, C.; Pan, H.; Liu, X. Recruitment of Hippodamia variegata by active volatiles from Glycyrrhiza uralensis and Alhagi sparsifolia plants infested with Aphis atrata. Pest Manag. Sci. 2024, 80, 355−365. [Google Scholar] [CrossRef]
- Osman, I.; Wang, Z.; Li, H.; Li, E.; Feng, H.; Yin, J.; Liang, G.; Liu, Z.; Ning, D.; Li, K.; et al. Attraction of Telenomus remus to egg volatiles of Spodoptera litura and oviposition-induced plant volatiles from tobacco (Nicotiana tabacum). Ecol. Evol. 2025, 15, e72012. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Zevallos, D.M.; Blande, J.D. Challenges of climate change and air pollution for volatile-mediated plant–parasitoid signalling. Curr. Opin. Insect Sci. 2024, 66, 101290. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; MO, B.T.; Zhang, J.Z. Semiochemicals for pest control in the age of synthetic biology. Emerg. Sci. Technol. 2025, 4, 1–12. [Google Scholar] [CrossRef]
- Fleischer, J.; Pregitzer, P.; Breer, H.; Krieger, J. Access to the odor world: Olfactory receptors and their role for signal transduction in insects. Cell. Mol. Life. Sci. 2018, 75, 485–508. [Google Scholar] [CrossRef]
- Zhang, J.; Raza, S.A.K.; Wei, Z.; Keesey, I.W.; Parker, A.L.; Feistel, F.; Chen, J.; Cassau, S.; Fandino, R.A.; Grosse-Wilde, E.; et al. Competing beetles attract egg laying in a hawkmoth. Curr. Biol. 2022, 32, 861−869.e8. [Google Scholar] [CrossRef]
- Andersson, M.N.; Biswas, T.; Yuvaraj, J.K. Highly Expressed odorant receptor orthologs detect the aggregation pheromone lineatin in Trypodendron Ambrosia Beetles. Mol. Ecol. 2025, 34, e70064. [Google Scholar] [CrossRef]
- Chang, H.; Unni, A.P.; Tom, M.T.; Cao, Q.; Liu, Y.; Wang, G.; Llorca, L.C.; Brase, S.; Bucks, S.; Weniger, K.; et al. Odorant detection in a locust exhibits unusually low redundancy. Curr. Biol. 2023, 33, 5427–5438.e5. [Google Scholar] [CrossRef]
- Sun, H.; Bu, L.A.; Su, S.C.; Guo, D.; Gao, C.F.; Wu, S.F. Knockout of the odorant receptor co-receptor, orco, impairs feeding, mating and egg-laying behavior in the fall armyworm Spodoptera frugiperda. Insect Biochem. Mol. Biol. 2023, 152, 103889. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.