
Synergistic Effects of Elevated CO2 and Enhanced Light Intensity on Growth Dynamics, Stomatal Phenomics, Leaf Anatomy, and Photosynthetic Performance in Tomato Seedlings

 ,
, 
 ,
, 
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material Preparation
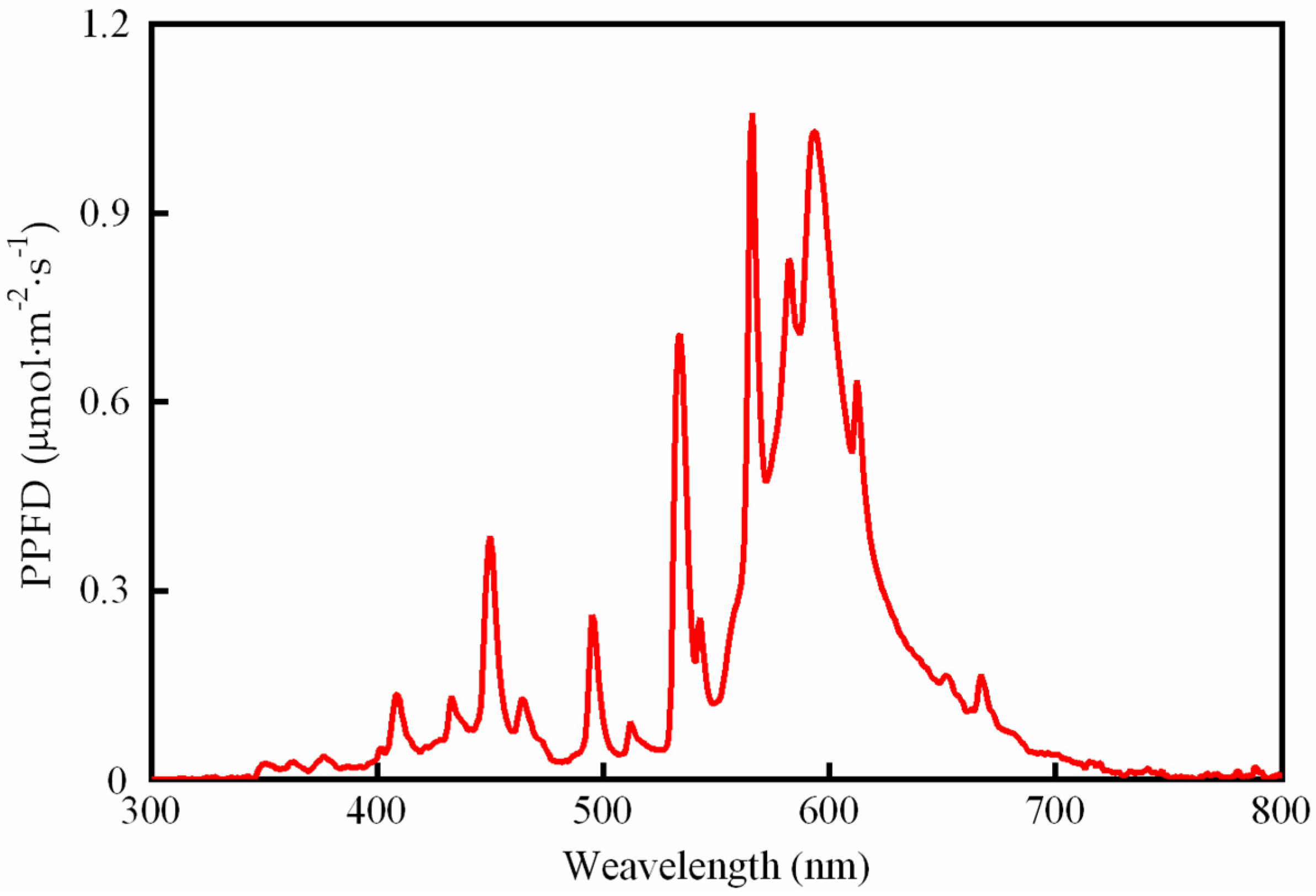
2.2. Experimental Design
2.3. Growth Parameter Quantification
2.4. Determination of Stomatal Traits
2.5. Determination of Leaf Anatomical Structure
2.6. Determination of Gas Exchange and Chlorophyll Fluorescence
2.7. Determination of Dry Mass Accumulation and Seedling Health Index
2.8. Statistical Analysis
3. Results
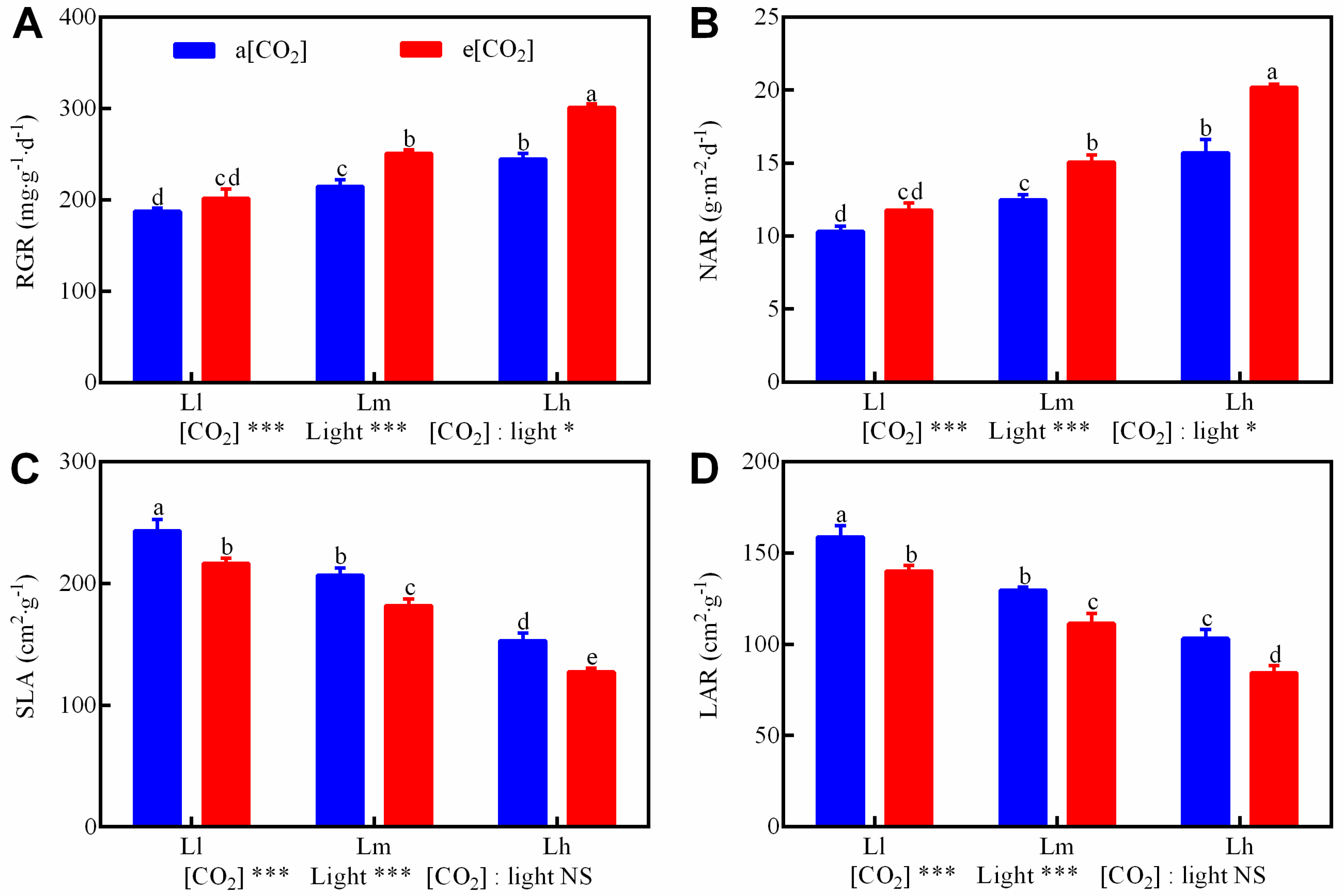
3.1. Growth Traits
3.2. Stomatal Traits
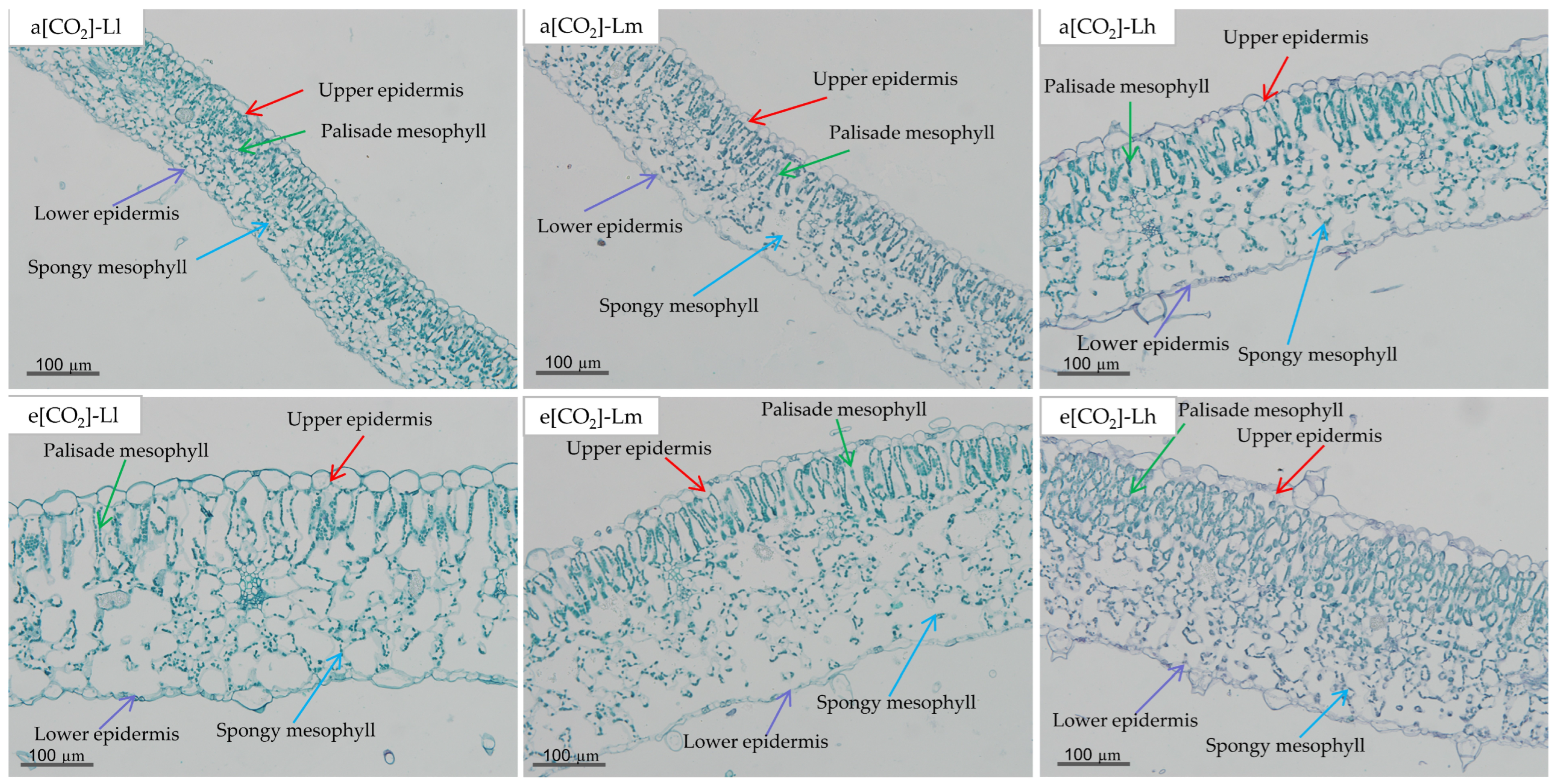
3.3. Leaf Anatomical Structure
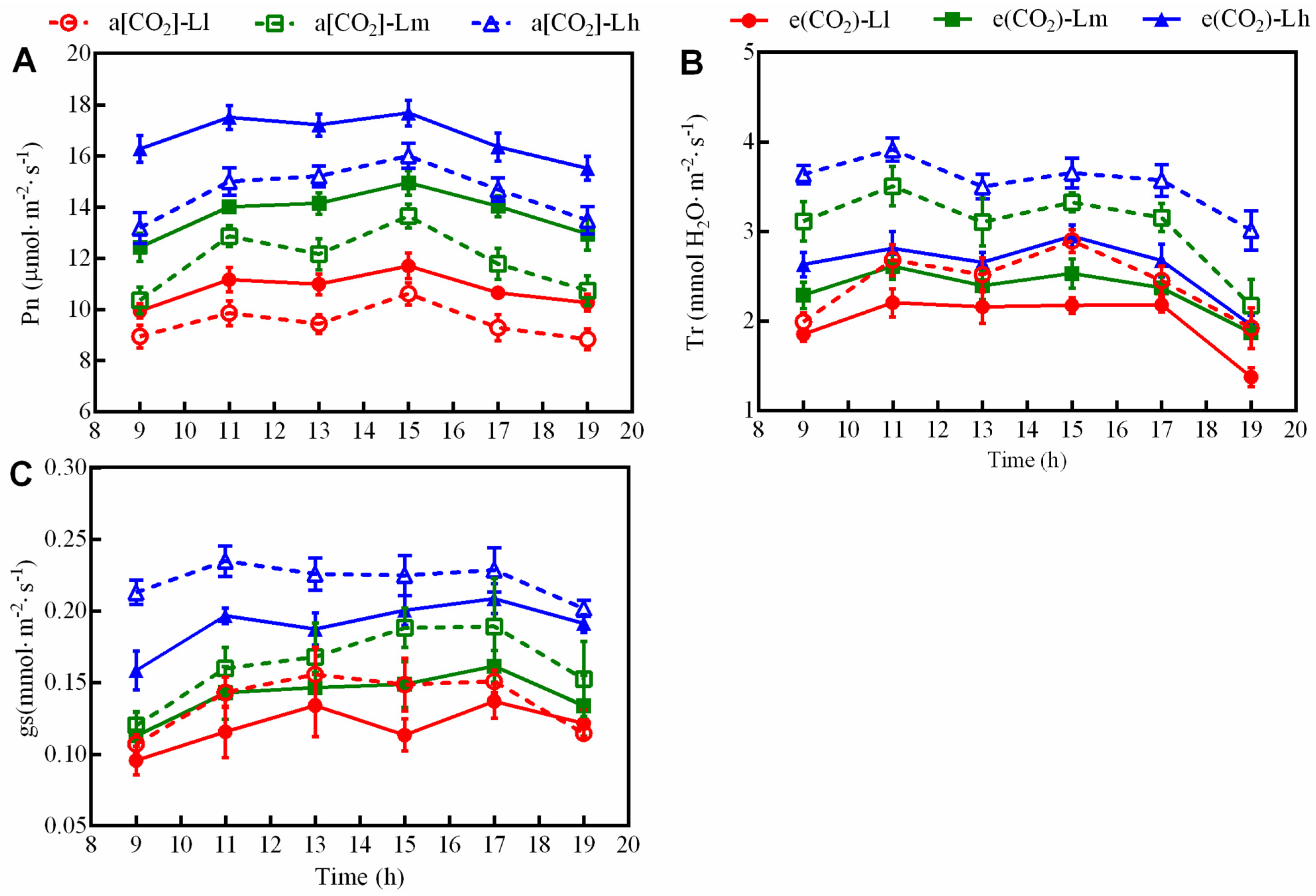
3.4. Diurnal Variation in Photosynthesis, Transpiration Rate, and Stomatal Conductance
3.5. Chlorophyll Fluorescence Parameters
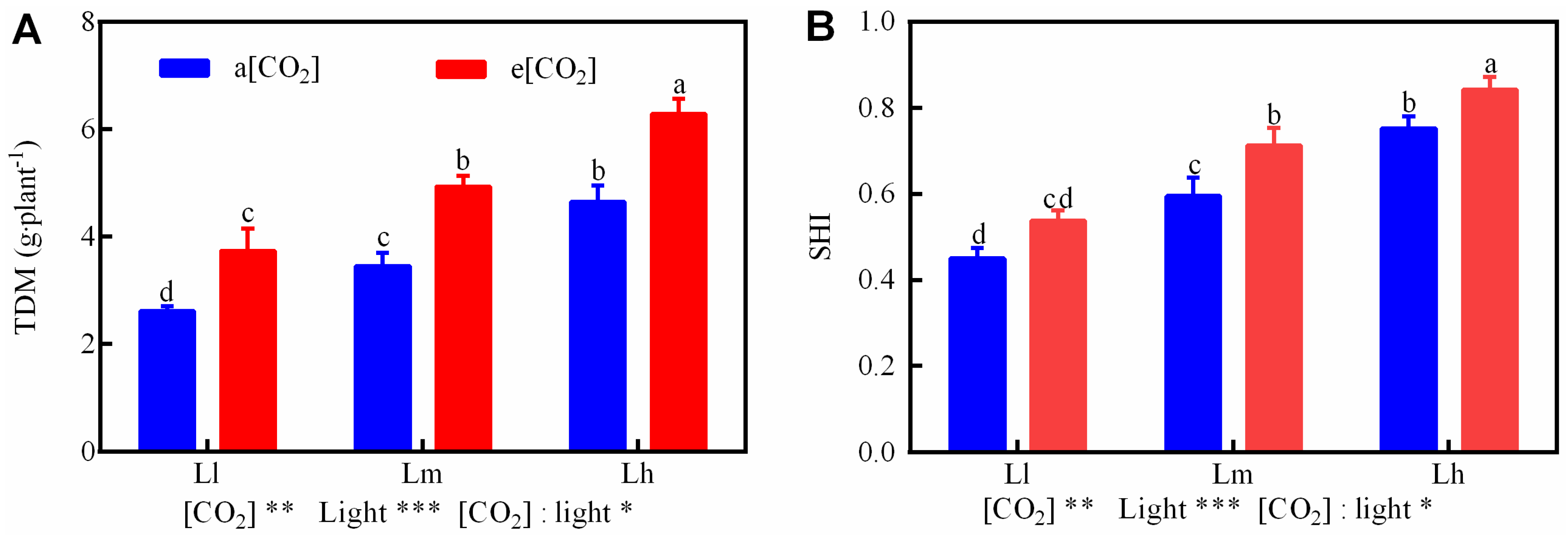
3.6. Total Dry Mass and Seedling Health Index
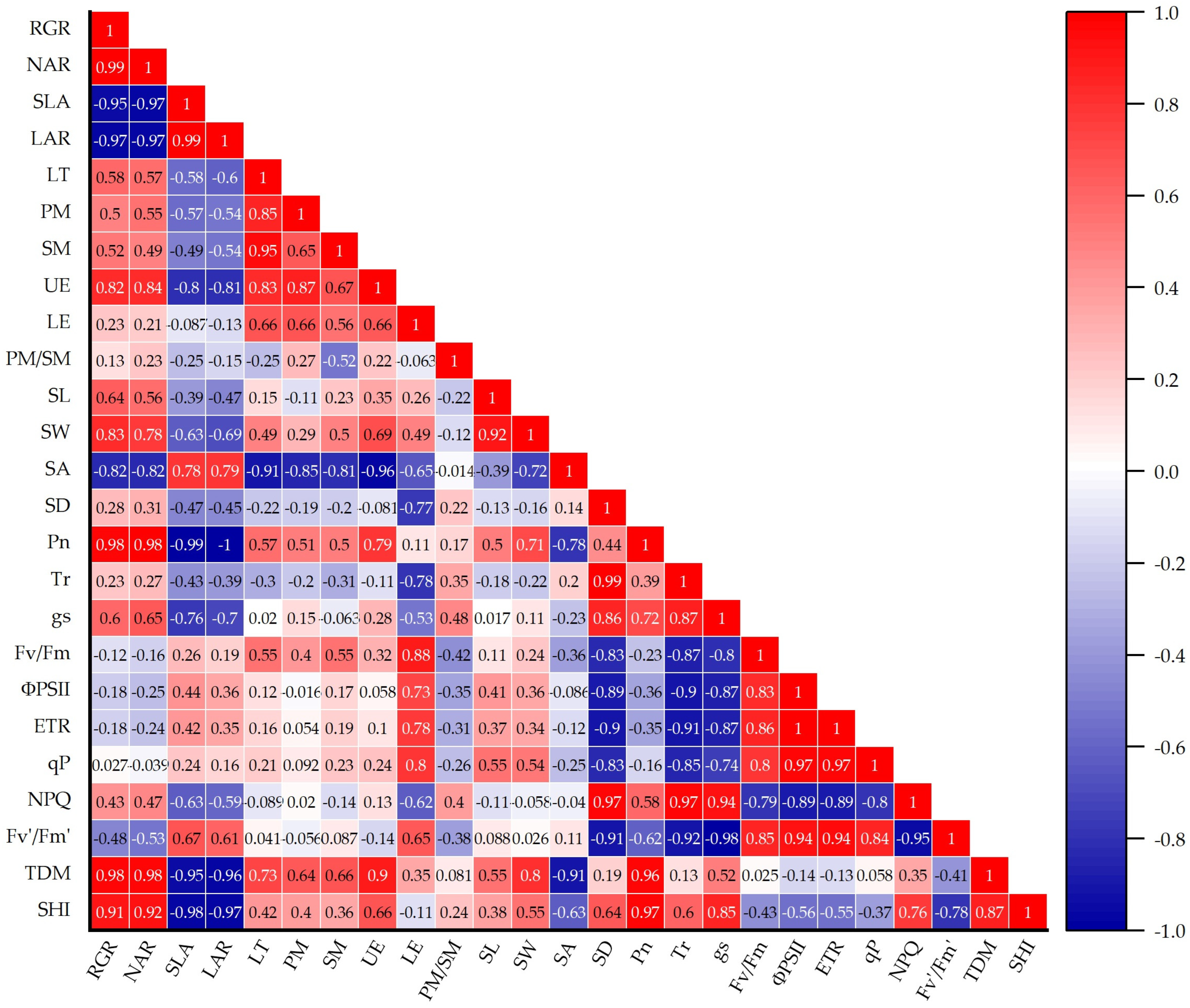
3.7. Correlations Among Plant Physiological and Morphological Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| a[CO2] | Ambient CO2 concentration |
| ANOVA | Analysis of variance |
| ATP | Adenosine triphosphate |
| CAM | Crassulacean acid metabolism |
| DAT | Day after treatment |
| e[CO2] | Elevated CO2 concentration |
| ETR | Electron transport rate |
| FAO | Food and Agriculture Organization of the United Nations |
| Fm | Maximum fluorescence |
| Fm′ | Maximum fluorescence under actinic light |
| F0 | Initial fluorescence |
| F0′ | Minimum fluorescence under actinic light |
| Fv/Fm | Maximum PSII photochemical efficiency |
| FS | Steady-state fluorescence |
| Fv′/Fm′ | Effective photochemical quantum efficiency |
| gs | Stomatal conductance |
| LAR | Leaf area ratio |
| LE | Lower epidermis |
| Lh | High light intensity |
| Ll | Low light intensity |
| Lm | Moderate light intensity |
| LT | Leaf thickness |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NAR | Net assimilation rate |
| NPQ | Non-photochemical quenching |
| PM | Palisade mesophyll |
| PM/SM | The ratio of palisade mesophyll to spongy mesophyll |
| Pn | Net photosynthetic rate |
| PPFD | Photosynthetic photon flux density |
| PSII | Photosystem II |
| ΦPSII | Actual PSII quantum yield |
| qP | Photochemical quenching coefficient |
| RGR | Relative growth rate |
| RuBisCO | Ribulose-1,5-bisphosphate carboxylase/oxygenase |
| RuBP | Ribulose 1,5-bisphosphate |
| ROS | Reactive oxygen species |
| SA | Stomatal aperture |
| SD | Stomatal density |
| SE | Standard error |
| SHI | Seedling health index |
| SL | Stomatal length |
| SLA | Specific leaf area |
| SM | Spongy mesophyll |
| SW | Stomatal width |
| TDM | Total dry mass per plant |
| Tr | Transpiration rate |
| UE | Upper epidermis |
References
- Xu, X.; Yang, F.; Song, J.; Zhang, R.; Cai, W. Does the daily light integral influence the sowing density of tomato plug seedlings in a controlled environment? Horticulturae 2024, 10, 730. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.; Yan, H.; Ullah, I.; Zuo, Z.; Li, L.; Yu, J. Effects of irrigation quantity and biochar on soil physical properties, growth characteristics, yield and quality of greenhouse tomato. Agric. Water Manag. 2020, 241, 106263. [Google Scholar] [CrossRef]
- FAO. FAOSTAT Database 2023. Available online: http://www.fao.org/statistics/en (accessed on 1 March 2025).
- Akhlaq, M.; Zhang, C.; Yan, H.; Ou, M.; Zhang, W.; Liang, S.; Ikram, R.M.A. Response of tomato growth to continuous elevated CO2 concentration under controlled environment. Int. J. Agric. Biol. Eng. 2022, 15, 51–59. [Google Scholar] [CrossRef]
- Kumi, F.; Mao, H.; Li, Q.; Luhua, H. Assessment of tomato seedling substrate-root quality using X-ray computed tomography and scanning electron microscopy. Appl. Eng. Agric. 2016, 32, 417–427. [Google Scholar] [CrossRef]
- Song, J.; Zhang, R.; Yang, F.; Wang, J.; Cai, W.; Zhang, Y. Nocturnal LED supplemental lighting improves quality of tomato seedlings by increasing biomass accumulation in a controlled environment. Agronomy 2024, 14, 1888. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, W.; Liu, C.; Xu, C.; Wei, G.; Cui, B.; Hou, J.; Wan, H.; Chen, Y.; Zhang, J.; et al. Effect of CO2 elevation on tomato gas exchange, root morphology and water use efficiency under two N-fertigation levels. Plants 2024, 13, 2373. [Google Scholar] [CrossRef]
- An, W.; Wang, G.; Dou, J.; Zhang, Y.; Zhang, Q.; He, Y.; Tang, Z.; Yu, J. Protective mechanisms of exogenous melatonin on chlorophyll metabolism and photosynthesis in tomato seedlings under heat stress. Front. Plant Sci. 2025, 16, 1519950. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, P.; Fadiji, T.; Li, Z.; Ni, J. Biomechanical response of the above-ground organs in tomato seedling at different age levels under wind-flow disturbance. Sci. Hortic. 2023, 312, 111835. [Google Scholar] [CrossRef]
- Ma, G.; Chen, X.; Liu, Y.; Hu, J.; Han, L.; Mao, H. Effects of compound biochar substrate coupled with water and nitrogen on the growth of cucumber plug seedlings. Agronomy 2022, 12, 2855. [Google Scholar] [CrossRef]
- Pan, T.; Ding, J.; Qin, G.; Wang, Y.; Xi, L.; Yang, J.; Li, J.; Zhang, J.; Zou, Z. Interaction of supplementary light and CO2 enrichment improves growth, photosynthesis, yield, and quality of tomato in autumn through spring greenhouse production. Hort. Sci. 2019, 54, 246–252. [Google Scholar] [CrossRef]
- Amarasinghe, A.A.Y.; Polwaththa, K.P.G.D.M.; Suratissa, D.M. Effects of elevated CO2 and light intensity on growth, yield, and nutritional quality of tomato (Solanum lycopersicum) in controlled environment agriculture systems. Am. J. Life Sci. Innov. 2025, 4, 1–7. [Google Scholar] [CrossRef]
- Esmaili, M.; Aliniaeifard, S.; Mashal, M.; Ghorbanzadeh, P.; Seif, M.; Gavilan, M.U.; Carrillo, F.F.; Lastochkina, O.; Li, T. CO2 enrichment and increasing light intensity till a threshold level, enhance growth and water use efficiency of lettuce plants in controlled environment. Not. Bot. Horti Agrobot. 2020, 48, 2244–2262. [Google Scholar] [CrossRef]
- Koo, J.K.; Hwang, H.S.; Hwang, J.H.; Park, E.W.; Yu, J.; Yun, J.H.; Hwang, S.Y.; Choi, H.E.; Hwang, S.J. Supplemental lighting and CO2 enrichment on the growth, fruit quality, and yield of cucumber. Hortic. Environ. Biotechnol. 2025, 66, 77–85. [Google Scholar] [CrossRef]
- Li, X.; Kang, S.; Li, F.; Zhang, X.; Huo, Z.; Ding, R.; Tong, L.; Du, T.; Li, S. Light supplement and carbon dioxide enrichment affect yield and quality of off-season pepper. Agron. J. 2017, 109, 2107–2118. [Google Scholar] [CrossRef]
- Pan, T.; Wang, Y.; Wang, L.; Ding, J.; Cao, Y.; Qin, G.; Yan, L.; Xi, L.; Zhang, J.; Zou, Z. Increased CO2 and light intensity regulate growth and leaf gas exchange in tomato. Physiol. Plant. 2020, 168, 694–708. [Google Scholar] [CrossRef]
- Soukharev, B. Why Humans Would Not Be Able to Stop Global Warming in the Coming Decades Even if There Were No Climate Feedbacks. In Global Warming and Mass Migration, 1st ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2025; Volume 4, pp. 113–173. [Google Scholar] [CrossRef]
- Kumari, S.; Agrawal, M. Growth, yield and quality attributes of a tropical potato variety (Solanum tuberosum L. cv Kufri chandramukhi) under ambient and elevated carbon dioxide and ozone and their interactions. Ecotoxicol. Environ. Saf. 2014, 101, 146–156. [Google Scholar] [CrossRef]
- Long, S.P.; Ainsworth, E.A.; Rogers, A.; Ort, D.R. Rising atmospheric carbon dioxide: Plants FACE the future. Annu. Rev. Plant Biol. 2004, 55, 591–628. [Google Scholar] [CrossRef]
- Keeley, J.E.; Rundel, P.W. Evolution of CAM and C4 carbon-concentrating mechanisms. Int. J. Plant Sci. 2003, 164, S55–S77. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. Does enhanced photosynthesis enhance growth? Lessons learned from CO2 enrichment studies. Plant Physiol. 2011, 155, 117–124. [Google Scholar] [CrossRef]
- Zhang, C.; Akhlaq, M.; Yan, H.; Ni, Y.; Liang, S.; Zhou, J.; Xue, R.; Li, M.; Adnan, R.M.; Li, J. Chlorophyll fluorescence parameter as a predictor of tomato growth and yield under CO2 enrichment in protective cultivation. Agric. Water Manag. 2023, 284, 108333. [Google Scholar] [CrossRef]
- Liu, B.; LI, M.; Li, Q.; Cui, Q.; Zhang, W.; Ai, X.; Bi, H. Combined effects of elevated CO2 concentration and drought stress on photosynthetic performance and leaf structure of cucumber (Cucumis sativus L.) seedlings. Photosynthetica 2018, 56, 942–952. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef]
- Yang, J.; Liang, T.; Liu, L.; Pan, T.; Zou, Z. Stomatal opening and growth in tomato seedlings treated with different proportions of red and blue light. Can. J. Plant Sci. 2019, 99, 688–700. [Google Scholar] [CrossRef]
- Zhou, J.; Li, P.; Wang, J.; Fu, W. Growth, photosynthesis, and nutrient uptake at different light intensities and temperatures in lettuce. Hort. Sci. 2019, 54, 1925–1933. [Google Scholar] [CrossRef]
- Song, J.; Fan, Y.; Li, X.; Li, Y.; Mao, H.; Zou, Z.; Zou, Z. Effects of daily light integral on tomato (Solanum lycopersicon L.) grafting and quality in a controlled environment. Int. J. Agric. Biol. Eng. 2022, 15, 44–50. [Google Scholar] [CrossRef]
- Tan, Y.; Duan, Y.; Chi, Q.; Wang, R.; Yin, Y.; Cui, D.; Li, S.; Wang, A.; Ma, R.; Li, B.; et al. The role of reactive oxygen species in plant response to radiation. Int. J. Mol. Sci. 2023, 24, 3346. [Google Scholar] [CrossRef]
- Li, Y.; Xin, G.; Liu, C.; Shi, Q.; Yang, F.; Wei, M. Effects of red and blue light on leaf anatomy, CO2 assimilation and the photosynthetic electron transport capacity of sweet pepper (Capsicum annuum L.) seedlings. Bmc Plant Biol. 2020, 20, 1–16. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Ntagkas, N.; Siebenkäs, A.; Mäenpää, M.; Matsubara, S.; Pons, T. A meta-analysis of plant responses to light intensity for 70 traits ranging from molecules to whole plant performance. New Phytol. 2019, 223, 1073–1105. [Google Scholar] [CrossRef]
- Ghorbanzadeh, P.; Aliniaeifard, S.; Esmaeili, M.; Mashal, M.; Azadegan, B.; Seif, M. Dependency of growth, water use efficiency, chlorophyll fluorescence, and stomatal characteristics of lettuce plants to light intensity. J. Plant Growth Regul. 2021, 40, 2191–2207. [Google Scholar] [CrossRef]
- Fan, X.; Xu, Z.; Liu, X.; Tang, C.; Wang, L.; Han, X. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hortic. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- He, W.; Huang, Z.; Li, J.; Su, W.; Gan, L.; Xu, Z. Effect of different light intensities on the photosynthate distribution in cherry tomato seedlings. J. Hortic. Sci. Biotechnol. 2019, 94, 611–619. [Google Scholar] [CrossRef]
- Khan, I.; Sohail; Zaman, S.; Li, G.; Fu, M. Adaptive responses of plants to light stress: Mechanisms of photoprotection and acclimation. A review. Front. Plant Sci. 2025, 16, 1550125. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Xing, D.; Wu, Y.; Wang, W.; Li, M.; Solangi, K. Diurnal variation in transport and use of intracellular leaf water and related photosynthesis in three karst plants. Agronomy 2022, 12, 2758. [Google Scholar] [CrossRef]
- Yan, H.; Acquah, S.; Zhang, C.; Wang, G.; Huang, S.; Zhang, H.; Zhao, B.; Wu, H. Energy partitioning of greenhouse cucumber based on the application of Penman-Monteith and Bulk Transfer models. Agric. Water Manag. 2019, 217, 201–211. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Casson, S.A.; Franklin, K.A.; Gray, J.E.; Grierson, C.S.; Whitelam, G.C.; Hetherington, A.M. Phytochrome B and PIF4 regulate stomatal development in response to light quantity. Curr. Biol. 2009, 19, 229–234. [Google Scholar] [CrossRef]
- Ando, E.; Kinoshita, T. Red light-induced phosphorylation of plasma membrane H+-ATPase in stomatal guard cells. Plant Physiol. 2018, 178, 838–849. [Google Scholar] [CrossRef]
- Akhlaq, M.; Miao, H.; Zhang, C.; Yan, H.; Run, X.; Chauhdary, J.N.; Rehman, M.M.U.; Li, J.; Ren, J. Resilience assessment of tomato crop chlorophyll fluorescence against water stress under elevated CO2 and protective cultivation. Irrig. Drain. 2025, 0, 1–19. [Google Scholar] [CrossRef]
- Chen, D.; Yuan, K.; Zhang, J.; Wang, Z.; Sun, Z.; Zhang, H.; Hu, J. Response analysis of fluorescence parameters of tomato seedlings oriented to vertical light environment adaptation. Plant Sci. 2022, 314, 111118. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, Z.; Li, J.; Chang, Y.; Du, Q.; Pan, T. Regulation of vapor pressure deficit by greenhouse micro-fog systems improved growth and productivity of tomato via enhancing photosynthesis during summer season. PLoS ONE 2015, 10, e0133919. [Google Scholar] [CrossRef]
- Mishra, A.K.; Gupta, S.; Agrawal, S.B.; Tiwari, S. Role of stomatal and leaf anatomical features in defining plant performance under elevated carbon dioxide and ozone, in the changing climate scenario. Environ. Sci. Pollut. Res. 2025, 32, 2536–2550. [Google Scholar] [CrossRef] [PubMed]
- Janova, J.; Kubasek, J.; Grams, T.E.E.; Zeisler-Diehl, V.; Schreiber, L.; Santrucek, J. Effect of light-induced changes in leaf anatomy on intercellular and cellular components of mesophyll resistance for CO2 in Fagus sylvatica. Plant Biol. 2024, 26, 842–854. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Chang, Z.; Lu, Y.; Tian, Y.; Zhou, H.; Wang, Y.; Liu, L.; Wang, P.; Zheng, Y.; Wu, J. Drought dampens the positive acclimation responses of leaf photosynthesis to elevated [CO2] by altering stomatal traits, leaf anatomy, and Rubisco gene expression in Pyrus. Environ. Exp. Bot. 2023, 211, 105375. [Google Scholar] [CrossRef]
- Li, X.; Schmid, B.; Wang, F.; Wang, T. Net assimilation rate determines the growth rates of 14 species of subtropical forest trees. PLoS ONE 2016, 11, e0150644. [Google Scholar] [CrossRef]
- Oguchi, R.; Ozaki, H.; Hanada, K.; Hikosaka, K. Which plant trait explains the variations in relative growth rate and its response to elevated carbon dioxide concentration among Arabidopsis thaliana ecotypes derived from a variety of habitats? Oecologia 2016, 180, 865–876. [Google Scholar] [CrossRef]
- Shipley, B. Trade-offs between net assimilation rate and specific leaf area in determining relative growth rate: Relationship with daily irradiance. Func. Ecol. 2002, 16, 682–689. [Google Scholar] [CrossRef]
- Centritto, M.; Lee, H.S.J.; Jarvis, P.G. Increased growth in elevated [CO2]: An early, short-term response? Glob. Change Biol. 1999, 5, 623–633. [Google Scholar] [CrossRef]
- Fu, W.; Li, P.; Wu, Y. Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce. Sci. Hortic. 2012, 135, 45–51. [Google Scholar] [CrossRef]
- Hattori, T.; Sonobe, K.; Inanaga, S.; An, P.; Tsuji, W.; Araki, H.; Eneji, A.E.; Morita, S. Short term stomatal responses to light intensity changes and osmotic stress in sorghum seedlings raised with and without silicon. Environ. Exp. Bot. 2007, 60, 177–182. [Google Scholar] [CrossRef]
- Driesen, E.; Ende, W.V.D.; Proft, M.D.; Saeys, W. Influence of environmental factors light, CO2, temperature, and relative humidity on stomatal opening and development: A review. Agronomy 2020, 10, 1975. [Google Scholar] [CrossRef]
- Lawson, T.; von Caemmerer, S.; Baroli, I. Photosynthesis and Stomatal Behaviour. In Progress in Botany, 1st ed.; Lüttge, U., Beyschlag, W., Büdel, B., Francis, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 72, pp. 265–304. [Google Scholar] [CrossRef]
- Lee, S.H.; Tewari, R.K.; Hahn, E.J.; Paek, K.Y. Photon flux density and light quality induce changes in growth, stomatal development, photosynthesis and transpiration of Withania somnifera (L.) Dunal. plantlets. Plant Cell Tissue Organ Cult. 2007, 90, 141–151. [Google Scholar] [CrossRef]
- Bao, Y.; Liu, X.; Feng, C.; Niu, M.; Liu, C.; Wang, H.; Yin, W.; Xia, X. Light and light signals regulate growth and development in woody plants. Forests 2024, 15, 523. [Google Scholar] [CrossRef]
- Wang, N.; Gao, G.; Wang, Y.; Wang, D.; Wang, Z.; Gu, J. Coordinated responses of leaf and absorptive root traits under elevated CO2 concentration in temperate woody and herbaceous species. Environ. Exp. Bot. 2020, 179, 104199. [Google Scholar] [CrossRef]
- De Graaff, M.A.; Van Groenigen, K.J.A.N.; Six, J.; Hungate, B.; Van Kessel, C. Interactions between plant growth and soil nutrient cycling under elevated CO2: A meta-analysis. Global Change Biol. 2006, 12, 2077–2091. [Google Scholar] [CrossRef]
- Radochová, B.; Tichá, I. Leaf anatomy during leaf development of photoautotrophically in vitro-grown tobacco plants as affected by growth irradiance. Biol. Plant. 2009, 53, 21–27. [Google Scholar] [CrossRef]
- Arantes, M.K.; da Silva Filho, M.P.; Pennacchi, J.P.; das Chagas Mendonça, A.M.; Barbosa, J.P.R.A.D. Phenotypic plasticity of leaf anatomical traits helps to explain gas-exchange response to water shortage in grasses of different photosynthetic types. Theor. Exp. Plant Physiol. 2020, 32, 341–356. [Google Scholar] [CrossRef]
- Matthews, J.S.; Vialet-Chabrand, S.R.M.; Lawson, T. Diurnal variation in gas exchange: The balance between carbon fixation and water loss. Plant Physiol. 2017, 174, 614–623. [Google Scholar] [CrossRef]
- Akhlaq, M.; Zhang, Z.; Yan, H.; Liang, S.; Ni, Y.; Zhou, J.; Xue, R.; Li, J.; Hussain, Z.; Iqbal, S. Exploring adequate CO2 elevation for optimum tomato growth and yield under protected cultivation. J. Plant Physiol. 2023, 289, 154093. [Google Scholar] [CrossRef]
- Kitao, M.; Tobita, H.; Kitaoka, S.; Harayama, H.; Yazaki, K.; Komatsu, M.; Agathokleous, E.; Koike, T. Light energy partitioning under various environmental stresses combined with elevated CO2 in three deciduous broadleaf tree species in Japan. Climate 2019, 7, 79. [Google Scholar] [CrossRef]
- Ma, F.; Na, X.; Xu, T. Drought responses of three closely related Caragana species: Implication for their vicarious distribution. Ecol. Evol. 2016, 6, 2763–2773. [Google Scholar] [CrossRef]
- Borsuk, A.M.; Roddy, A.B.; Theroux-Rancourt, G.; Brodersen, C.R. Structural organization of the spongy mesophyll. New Phytol. 2022, 234, 946–960. [Google Scholar] [CrossRef] [PubMed]
- Marçal, D.M.S.; Avila, R.T.; Quiroga-Rojas, L.F.; de Souza, R.P.B.; Junior, C.C.G.; Ponte, L.R.; Barbosa, M.L.; Oliveira, L.A.; Martins, S.C.V.; Ramalho, J.D.C.; et al. Elevated [CO2] benefits coffee growth and photosynthetic performance regardless of light availability. Plant Physiol. Biochem. 2021, 158, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Dun, Z.; Wang, Y.; Yang, D.; Xiong, D.; Cui, K.; Peng, S.; Huang, J. Effect of stomatal morphology on leaf photosynthetic induction under fluctuating light in rice. Front. Plant Sci. 2022, 12, 754790. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zou, J.; Lin, S.; Jin, C.; Shi, M.; Yang, B.; Yang, Y.; Jin, D.; Li, R.; Li, Y.; et al. Effects of different light intensity on the growth of tomato seedlings in a plant factory. PLoS ONE 2023, 18, e0294876. [Google Scholar] [CrossRef]
- O’Carrigan, A.; Hinde, E.; Lu, N.; Xu, X.; Duan, H.; Huang, G.; Mak, M.; Bellotti, B.; Chen, Z. Effects of light irradiance on stomatal regulation and growth of tomato. Environ. Exp. Bot. 2014, 98, 65–73. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | SL (µm) | SW (µm) | SA (µm) | SD (No.mm−2) |
|---|---|---|---|---|
| a[CO2]-Ll | 22.52 ± 0.56 c | 10.50 ± 0.67 cd | 2.05 ± 0.11 bc | 302.61 ± 18.59 d |
| e[CO2]-Ll | 18.87 ± 0.62 d | 10.50 ± 0.36 cd | 1.96 ± 0.12 c | 229.30 ± 15.51 e |
| a[CO2]-Lm | 23.15 ± 0.33 c | 11.50 ± 0.30 c | 2.29 ± 0.10 b | 440.11 ± 19.73 b |
| e[CO2]-Lm | 26.30 ± 0.48 b | 14.64 ± 0.28 b | 2.11 ± 0.10 bc | 335.96 ± 10.99 c |
| a[CO2]-Lh | 19.10 ± 0.26 d | 10.03 ± 0.20 d | 2.41 ± 0.08 a | 543.30 ± 15.44 a |
| e[CO2]-Lh | 27.53 ± 0.51 a | 16.64 ± 0.67 a | 2.15 ± 0.17 bc | 351.01 ± 11.62 c |
| Analysis of variance | ||||
| [CO2] | ** | ** | ** | ** |
| Light | * | ** | * | *** |
| [CO2]:light | NS | NS | * | ** |
| Treatments | LT (μm) | PM (μm) | SM (μm) | UE (μm) | LE (μm) | PM/SM |
|---|---|---|---|---|---|---|
| a[CO2]-Ll | 133.39 ± 3.19 d | 39.83 ± 0.83 e | 76.89 ± 0.85 f | 12.50 ± 0.21 e | 4.17 ± 0.07 e | 0.52 ± 0.01 a |
| e[CO2]-Ll | 295.46 ± 2.55 a | 102.08 ± 1.95 a | 166.30± 1.81 b | 20.83 ± 0.43 b | 6.25 ± 0.12 a | 0.53 ± 0.02 c |
| a[CO2]-Lm | 158.82 ± 2.39 c | 49.17 ± 2.00 d | 88.93 ± 1.65 e | 16.29 ± 0.42 d | 4.43 ± 0.10 d | 0.55 ± 0.02 a |
| e[CO2]-Lm | 299.71 ± 8.98 a | 66.67 ± 1.25 c | 208.33 ± 1.97 a | 19.48 ± 0.43 bc | 5.23 ± 0.14 c | 0.32± 0.02 b |
| a[CO2]-Lh | 243.17 ± 5.04 b | 79.33 ± 0.75 b | 141.58 ± 3.46 d | 18.75 ± 0.30 c | 3.59 ± 0.23 f | 0.56 ± 0.01 b |
| e[CO2]-Lh | 283.84 ± 9.30 a | 96.26 ± 4.08 a | 156.11 ± 2.77 c | 25.71 ± 0.36 a | 5.76 ± 0.21 b | 0.62 ± 0.02 a |
| Analysis of variance | ||||||
| [CO2] | ** | ** | ** | * | * | ** |
| Light | * | * | ** | * | ** | ** |
| [CO2]:light | ** | * | * | ** | ** | * |
| Treatments | Fv/Fm | ΦPSII | ETR | qP | NPQ | Fv′/Fm′ |
|---|---|---|---|---|---|---|
| a[CO2]-Ll | 0.54 ± 0.01 d | 0.51 ± 0.00 d | 125.85 ± 1.15 c | 0.50 ± 0.01 c | 0.68 ± 0.01 b | 0.52 ± 0.02 c |
| e[CO2]-Ll | 0.63 ± 0.01 c | 0.58 ± 0.01 c | 158.54 ± 0.94 b | 0.61 ± 0.01 b | 0.59 ± 0.00 c | 0.66 ± 0.00 b |
| a[CO2]-Lm | 0.70 ± 0.02 b | 0.63 ± 0.01 b | 144.56 ± 2.54 ab | 0.56 ± 0.02 b | 0.65 ± 0.01 c | 0.61 ± 0.01 ab |
| e[CO2]-Lm | 0.78 ± 0.04 a | 0.71 ± 0.00 a | 167.63 ± 3.98 a | 0.66 ± 0.01 a | 0.53 ± 0.01 d | 0.70 ± 0.00 a |
| a[CO2]-Lh | 0.69 ± 0.02 b | 0.64 ± 0.01 b | 153.54 ± 6.37 b | 0.58 ± 0.02 ab | 0.71 ± 0.01 a | 0.63 ± 0.01 b |
| e[CO2]-Lh | 0.74 ± 0.01 a | 0.69 ± 0.01 a | 169.93 ± 5.23 a | 0.65 ± 0.03 a | 0.64 ± 0.02 c | 0.69 ± 0.02 a |
| Analysis of variance | ||||||
| [CO2] | * | ** | * | * | ** | ** |
| Light | ** | *** | * | * | ** | * |
| [CO2]:light | * | ** | * | * | ** | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, T.; Zhang, W.; Du, W.; Fu, B.; Zhou, X.; Cao, K.; Bao, E.; Wang, Y.; Lv, G. Synergistic Effects of Elevated CO2 and Enhanced Light Intensity on Growth Dynamics, Stomatal Phenomics, Leaf Anatomy, and Photosynthetic Performance in Tomato Seedlings. Horticulturae 2025, 11, 760. https://doi.org/10.3390/horticulturae11070760
Pan T, Zhang W, Du W, Fu B, Zhou X, Cao K, Bao E, Wang Y, Lv G. Synergistic Effects of Elevated CO2 and Enhanced Light Intensity on Growth Dynamics, Stomatal Phenomics, Leaf Anatomy, and Photosynthetic Performance in Tomato Seedlings. Horticulturae. 2025; 11(7):760. https://doi.org/10.3390/horticulturae11070760
Chicago/Turabian StylePan, Tonghua, Wenya Zhang, Wentao Du, Bingyan Fu, Xiaoting Zhou, Kai Cao, Encai Bao, Yunlong Wang, and Gaoqiang Lv. 2025. "Synergistic Effects of Elevated CO2 and Enhanced Light Intensity on Growth Dynamics, Stomatal Phenomics, Leaf Anatomy, and Photosynthetic Performance in Tomato Seedlings" Horticulturae 11, no. 7: 760. https://doi.org/10.3390/horticulturae11070760
APA StylePan, T., Zhang, W., Du, W., Fu, B., Zhou, X., Cao, K., Bao, E., Wang, Y., & Lv, G. (2025). Synergistic Effects of Elevated CO2 and Enhanced Light Intensity on Growth Dynamics, Stomatal Phenomics, Leaf Anatomy, and Photosynthetic Performance in Tomato Seedlings. Horticulturae, 11(7), 760. https://doi.org/10.3390/horticulturae11070760