



Hyperhydricity-Induced Physiological Changes and Catechin Accumulation in Blueberry Hybrids (Vaccinium corymbosum × V. angustifolium)




Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials, Culture Conditions
2.2. Shoot Proliferation in Sigma Glass and Growtek Stationary Bioreactor
2.3. Content of Water
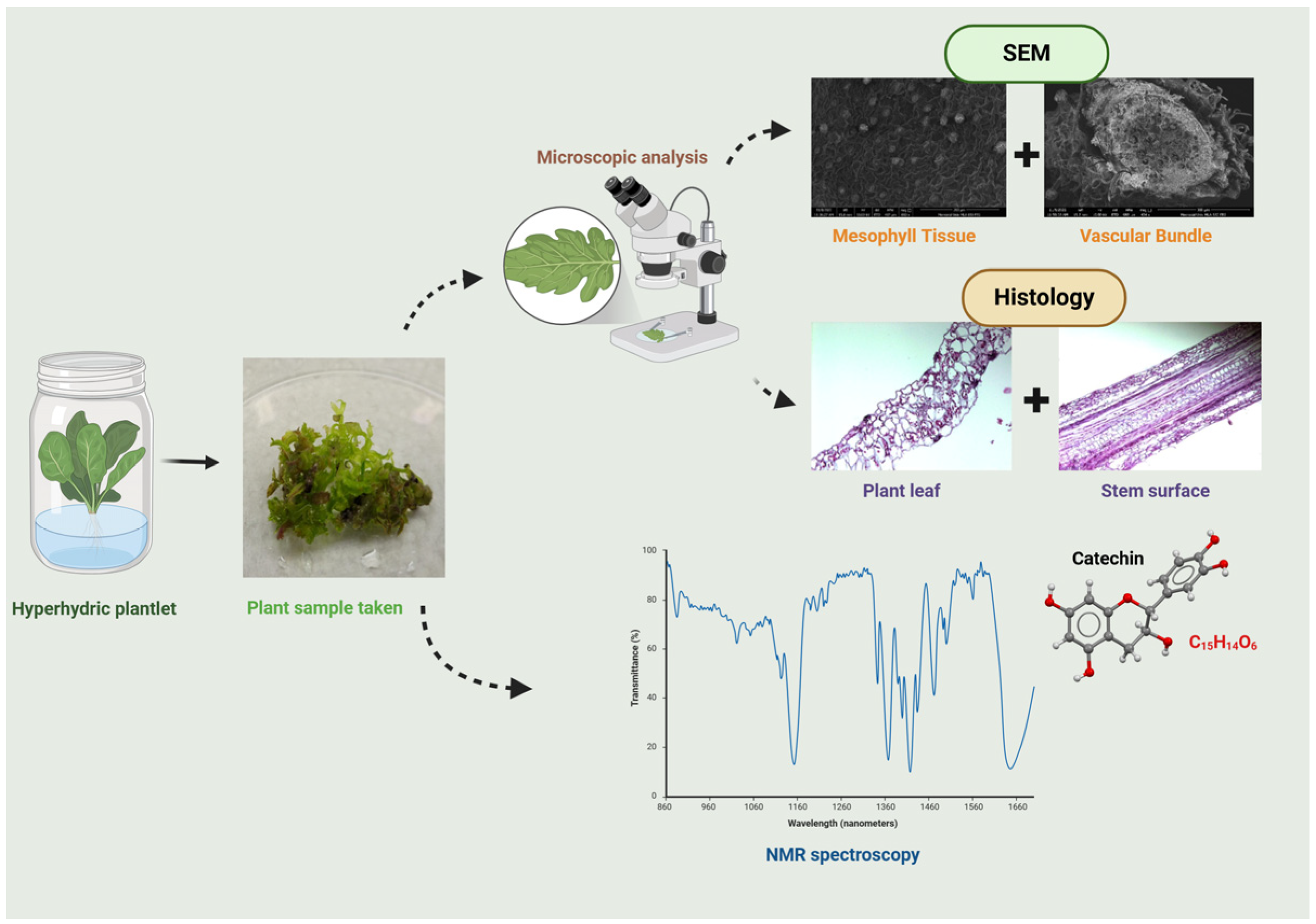
2.4. Observation of SEM
2.5. Histology Analysis
2.6. Bioactive Compound Analysis
2.6.1. Reference Compound and Reagents
2.6.2. Sample Preparation for NMR Analysis
2.6.3. NMR Data Acquisition and Analysis
2.7. Statistical Analysis
3. Results
3.1. Hyperhydricity in Micropropagated Blueberry
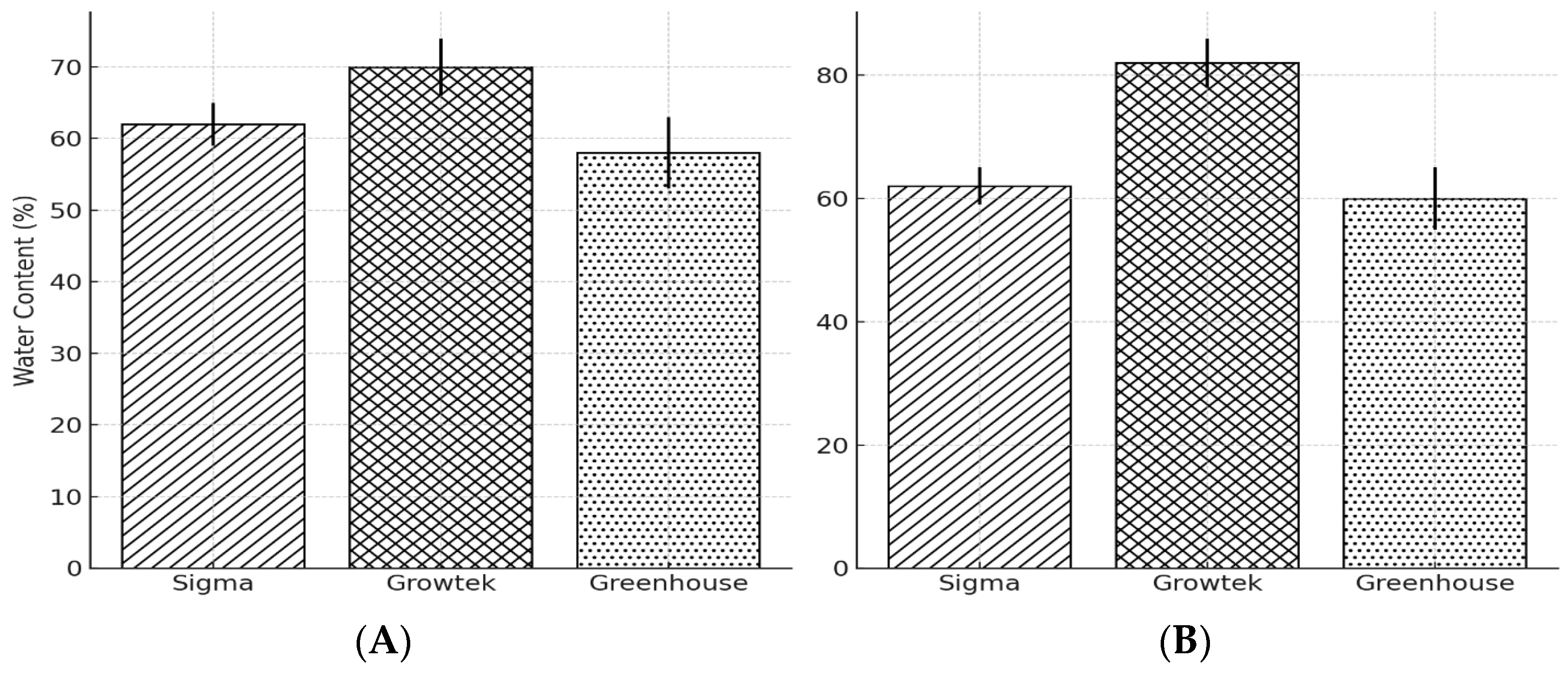
3.2. Water Content of Hyperhydric Plant
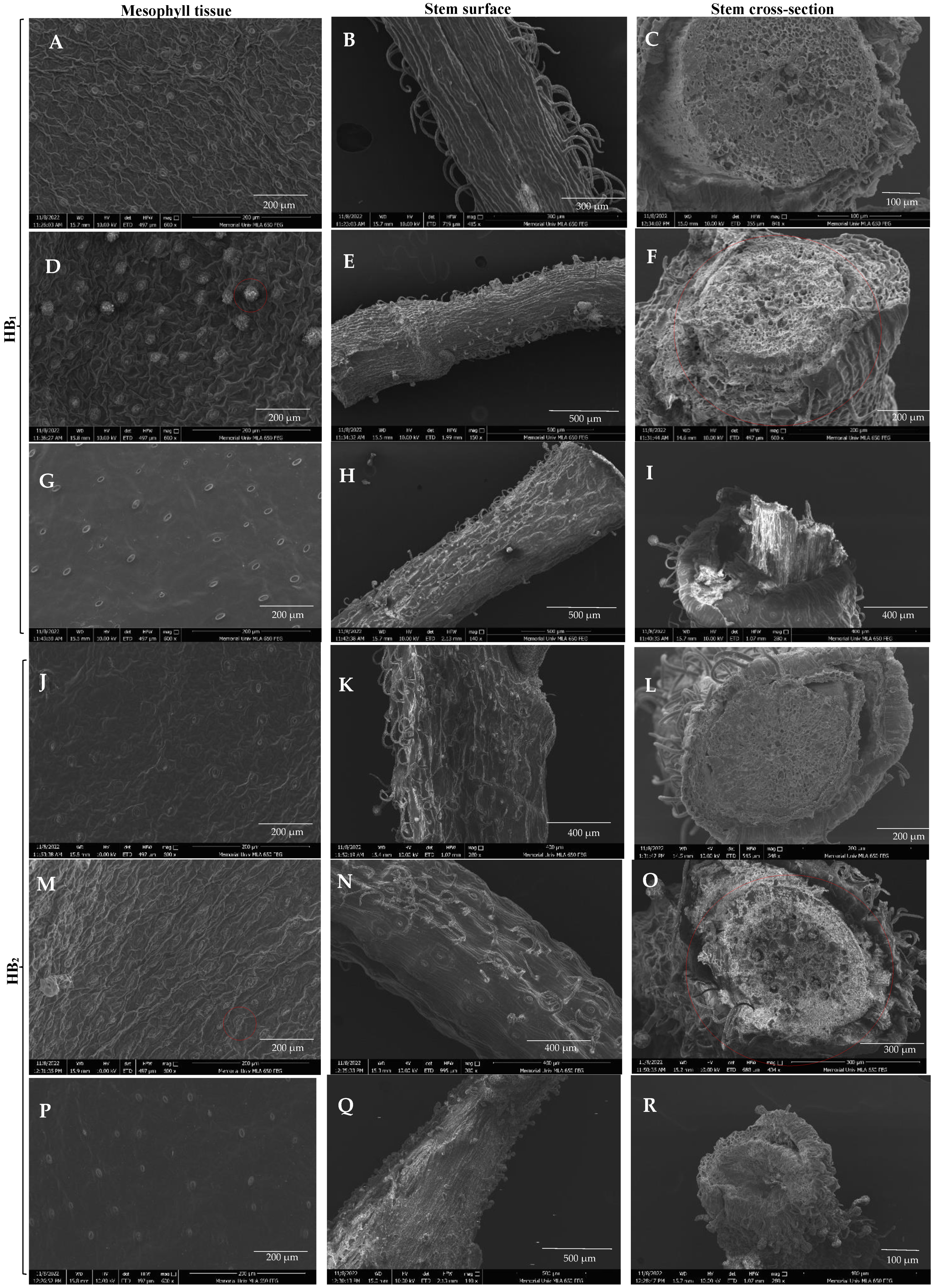
3.3. SEM Analysis of Leaf and Stem
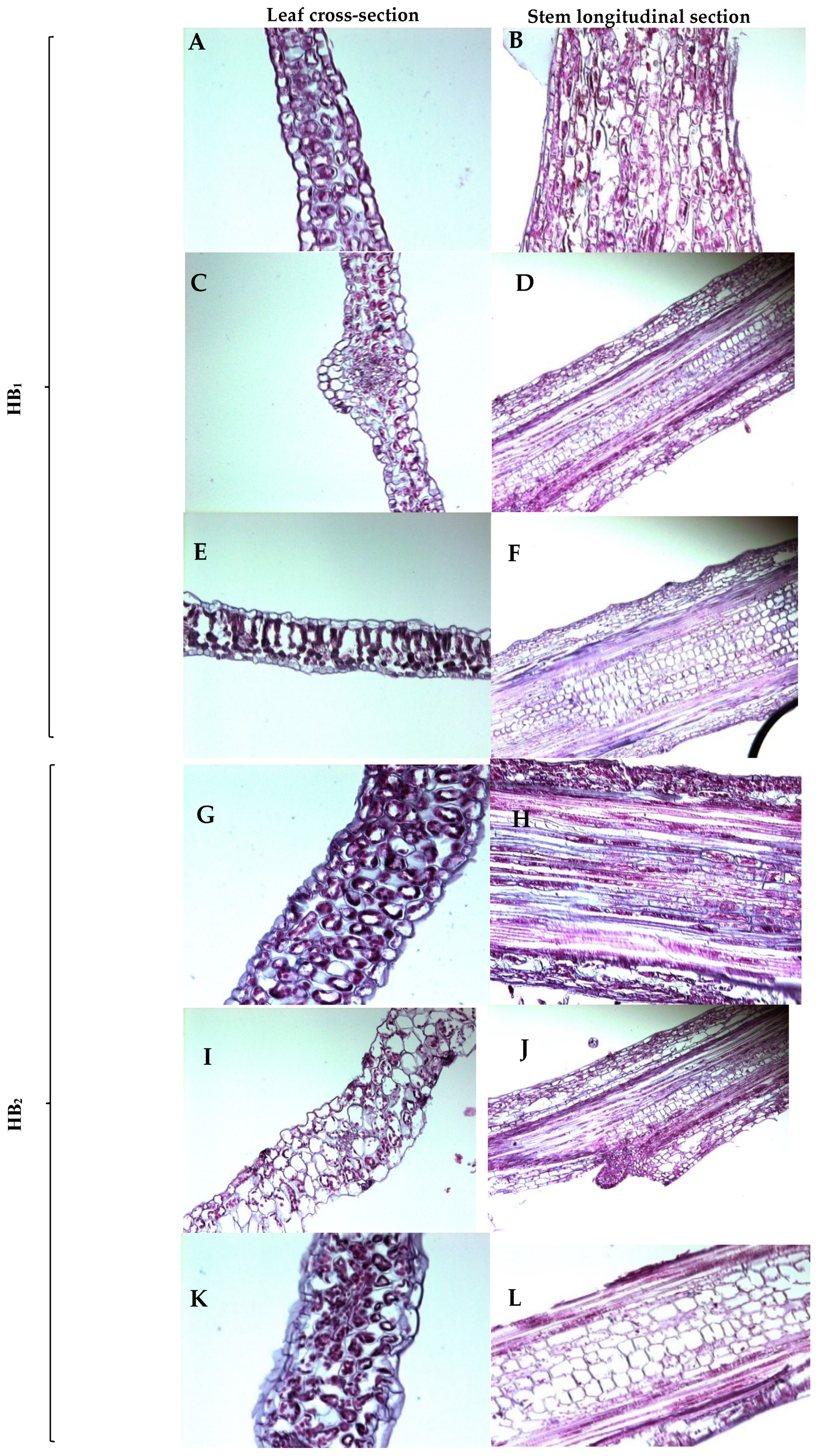
3.4. Histology of Hyperhydric and Non-Hyperhydric Plants
3.5. 1H NMR Analysis of Hyperhydric and Non-Hyperhydric Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosh, A.; Igamberdiev, A.U.; Debnath, S.C. Thidiazuron induced somatic embryogenesis and changes of antioxidant properties in tissue cultures of half-high blueberry plants. Sci. Rep. 2018, 8, 16978. [Google Scholar] [CrossRef] [PubMed]
- Barua, R.; Kundu, S.; Majumder, S.; Marshall, H.D.; Igamberdiev, A.U.; Debnath, S.C. Exploring two bioreactor systems for micropropagation of Vaccinium membranaceum and the antioxidant enzyme profiling in tissue culture-raised plants. Plant Growth Regul. 2025, 105, 1–15. [Google Scholar] [CrossRef]
- Sikdar, A.; Sharma, U.; Barua, R.; Igamberdiev, A.U.; Debnath, S.C. Epigenomic insight of lingonberry: Health promoting traits during micropropagation. Sci. Rep. 2021, 12, 12487. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Cao, G.; Martin, A.; Sofic, E.; Mcewen, J.; O’Brien, C.; Lischner, N.; Ehlenfeldt, M.; Kalt, W.; Krewer, G.; et al. Antioxidant capacity as influenced by total phenolic and anthocyanin content, maturity, and variety of Vaccinium species. J. Agric. Food Chem. 1998, 46, 2686–2693. [Google Scholar] [CrossRef]
- Smith, G.J.; Thomsen, S.J.; Markham, K.R.; Andary, C.; Cardon, D. The photostabilities of naturally occurring 5-hydroxyflavones, flavonols, their glycosides and their aluminium complexes. J. Photochem. Photobiol. A Chem. 2000, 136, 87–91. [Google Scholar] [CrossRef]
- Riihinen, K.; Jaakola, L.; Kärenlampi, S.; Hohtola, A. Organ specific distribution of phenolic compounds in bilberry (Vaccinium myrtillus) and “northblue” blueberry (Vaccinium rymbosum x V. angustifolium). Food Chem. 2008, 110, 156–160. [Google Scholar] [CrossRef]
- Takeshita, M.; Ishida, Y.I.; Akamatsu, E.; Ohmori, Y.; Sudoh, M.; Oto, H.; Tsubouchi, H.; Kataoka, H. Proanthocyanidin from blueberry leaves suppresses expression of subgenomic hepatitis C virus RNA. J. Biol. Chem. 2009, 284, 21165–21176. [Google Scholar] [CrossRef]
- Skupien, K. Evaluation of chemical composition of fresh and frozen blueberry fruit (Vaccinium corymbosum L.). Acta Sci. Pol. Hortorum Cultus 2006, 5, 19–25. [Google Scholar]
- Sweeney, M.I.; Kalt, W.; MacKinnon, S.L.; Ashby, J.; Gottschall-Pass, K.T. Feeding rats diets enriched in lowbuch blueberries for six weeks decreases ischemia-induced brain damage. Nutr. Neurosci. 2002, 5, 427–431. [Google Scholar] [CrossRef]
- Neto, C.C. Cranberry and Blueberry: Evidence for protective effects against cancer and vascular diseases. Mol. Nutr. Food Res. 2007, 51, 652–664. [Google Scholar] [CrossRef]
- Martineau, L.C.; Couture, A.; Spoor, D.; Benhaddou-Andaloussi, A.; Harris, C.; Meddah, B.; Leduc, C.; Burt, A.; Vuong, T.; Mai Le, P.; et al. Anti-Diabetic properties of the canadian lowbush blueberry Vaccinium angustifolium Ait. Phytomedicine 2006, 13, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Papandreou, M.A.; Dimakopoulou, A.; Linardaki, Z.I.; Cordopatis, P.; Klimis-Zacas, D.; Margarity, M.; Lamari, F.N. Effect of a polyphenol-rich wild blueberry extract on cognitive performance of mice, brain antioxidant markers and acetylcholinesterase activity. Behav. Brain Res. 2009, 198, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Li, J.; Ji, H.; An, L.; Xia, X. Hyperhydricity induced ultrastructural and physiological changes in blueberry (Vaccinium spp.). Plant Cell Tissue Organ Cult. 2018, 133, 65–76. [Google Scholar] [CrossRef]
- Olmos, E.; Hellin, E. Ultrastructural differences of hyperhydric and normal leaves from regenerated carnation plants. Sci. Hortic. 1998, 75, 91–101. [Google Scholar] [CrossRef]
- Liu, P.; Lindstedt, A.; Markkinen, N.; Sinkkonen, J.; Suomela, J.P.; Yang, B. Characterization of metabolite profiles of leaves of bilberry (Vaccinium myrtillus L.) and lingonberry (Vaccinium vitis-idaea L.). J. Agric. Food Chem. 2014, 62, 12015–12026. [Google Scholar] [CrossRef]
- Brown, E.; Gill, C.; Stewart, D.; McDougall, G. Lingonberries (Vaccinium vitis-idaea L.) and blueberries (Vaccinium crymbosum) contain discrete epicatechin anthocyanin derivatives linked by ethyl bridges. J. Berry Res. 2016, 6, 13–23. [Google Scholar] [CrossRef]
- Nierop, K.G.J.; Preston, C.M.; Kaal, J. Thermally Assisted Hydrolysis and Methylation of Purified Tannins from Plants. Anal. Chem. 2005, 77, 5604–5614. [Google Scholar] [CrossRef]
- Gao, H.; Xia, X.; An, L.; Xin, X.; Liang, Y. Reversion of hyperhydricity in Pink (Dianthus chinensis L.) plantlets by AgNO3 and its associated mechanism during in vitro culture. Plant Sci. 2017, 254, 1–11. [Google Scholar] [CrossRef]
- Picoli, E.A.T.; Paiva, E.A.S.; Xavier, A.; Aguiar, R.M.; Carolino, S.M.B.; Fari, M.G.; Otoni, W.C. Ultrastructural and biochemical aspects of normal and hyperhydric eucalypt. Int. J. Hortic. Sci. 2008, 14, 61–69. [Google Scholar] [CrossRef]
- de Almeida, M.; de Almeida, C.V.; Graner, E.M.; Brondani, G.E.; de Abreu-Tarazi, M.F. Pre-Procambial Cells Are Niches for Pluripotent and Totipotent Stem-like Cells for Organogenesis and Somatic Embryogenesis in the Peach Palm: A Histological Study. Plant Cell Rep. 2012, 31, 1495–1515. [Google Scholar] [CrossRef]
- Ivanova, M.; Van Staden, J. Effect of ammonium ions and cytokinins on hyperhydricity and multiplication rate of in vitro regenerated shoots of Aloe polyphylla. Plant Cell Tissue Organ Cult. 2008, 92, 227–231. [Google Scholar] [CrossRef]
- Williams, R.R.; Taji, A.M. Effect of temperature, gel concentration and cytokinins on vitrffication of Olearia microdisca (J.M. Black) in vitro shoot cultures. Plant Cell Tissue Organ Cult. 1991, 26, 1–6. [Google Scholar] [CrossRef]
- Modi, P.; Sinha, A.; Kothari, S.L. Floriculture and ornamental biotechnology reduction of hyperhydricity in micropropagated french marigold (Tagetes patula L.) plants by modified medium parameters. Floric. Ornam. Biotechnol. 2009, 3, 40–45. [Google Scholar]
- Yadav, M.K.; Gaur, A.K.; Garg, G.K. Development of suitable protocol to overcome hyperhydricity in carnation during micropropagation. Plant Cell Tissue Organ Cult. 2003, 72, 153–156. [Google Scholar] [CrossRef]
- Debnath, S.C. Temporary Immersion and Stationary Bioreactors for Mass Propagation of True-to-Type Highbush, Half-High, and Hybrid Blueberries (Vaccinium spp.). J. Hortic. Sci. Biotechnol. 2017, 92, 72–80. [Google Scholar] [CrossRef]
- Acharya, B.R.; Assmann, S.M. Hormone interactions in stomatal function. Plant Mol. Biol. 2009, 69, 451–462. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Díez-Villaseñor, C.; García-Martínez, J.; Soria, E. Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. J. Mol. Evol. 2005, 60, 174–182. [Google Scholar] [CrossRef]
- Louro, R.P.; Dos Santos, A.V.; Machado, R.D. Ultrastructure of Eucalyptus grandis × Eucalyptus europhylla I. shoots cultivated in vitro in multiplication and elongation rooting media. Int. J. Plant Sci. 1999, 160, 217–227. [Google Scholar] [CrossRef]
- Jausoro, V.; Llorente, B.E.; Apóstolo, N.M. Structural differences between hyperhydric and normal in vitro shoots of Handroanthus impetiginosus (Mart. Ex DC) Mattos (Bignoniaceae). Plant Cell Tissue Organ Cult. 2010, 101, 183–191. [Google Scholar] [CrossRef]
- Kevers, C.; Franck, T.; Strasser, R.J.; Dommes, J.; Gaspar, T. Hyperhydricity of micropropagated shoots: A typically stress induced change of physiological state. Plant Cell Tissue Organ Cult. 2004, 77, 181–191. [Google Scholar] [CrossRef]
- Gantait, S.; Mahanta, M. Hyperhydricity-induced changes among in vitro regenerants of Gerbera. S. Afr. J. Bot. 2022, 149, 496–501. [Google Scholar] [CrossRef]
- Chobot, V.; Huber, C.; Trettenhahn, G.; Hadacek, F. (±)-Catechin: Chemical weapon, antioxidant, or stress regulator? J. Chem. Ecol. 2009, 35, 980–996. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hybrid | Culture Conditions | Shoot No. per Explants | Hyperhydricity (%) |
|---|---|---|---|
| HB1 | Sigma | 6.78 ± 0.14 c | 15.45 ± 1.59 d |
| Growtek | 7.00 ± 0.10 b | 70.29 ± 1.67 b | |
| HB2 | Sigma | 7.25 ± 0.19 b | 18.67 ± 1.72 c |
| Growtek | 8.20 ± 0.18 a | 75.21 ± 1.89 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barua, R.; Kundu, S.; Igamberdiev, A.U.; Debnath, S.C. Hyperhydricity-Induced Physiological Changes and Catechin Accumulation in Blueberry Hybrids (Vaccinium corymbosum × V. angustifolium). Horticulturae 2025, 11, 418. https://doi.org/10.3390/horticulturae11040418
Barua R, Kundu S, Igamberdiev AU, Debnath SC. Hyperhydricity-Induced Physiological Changes and Catechin Accumulation in Blueberry Hybrids (Vaccinium corymbosum × V. angustifolium). Horticulturae. 2025; 11(4):418. https://doi.org/10.3390/horticulturae11040418
Chicago/Turabian StyleBarua, Rajesh, Sayani Kundu, Abir U. Igamberdiev, and Samir C. Debnath. 2025. "Hyperhydricity-Induced Physiological Changes and Catechin Accumulation in Blueberry Hybrids (Vaccinium corymbosum × V. angustifolium)" Horticulturae 11, no. 4: 418. https://doi.org/10.3390/horticulturae11040418
APA StyleBarua, R., Kundu, S., Igamberdiev, A. U., & Debnath, S. C. (2025). Hyperhydricity-Induced Physiological Changes and Catechin Accumulation in Blueberry Hybrids (Vaccinium corymbosum × V. angustifolium). Horticulturae, 11(4), 418. https://doi.org/10.3390/horticulturae11040418