Physiological Response of Macadamia (Macadamia integrifolia) Seedlings to Drought Stress


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Drought Treatment
2.2. Measurement Indicators and Methods
2.2.1. Observation of Seedling Morphology
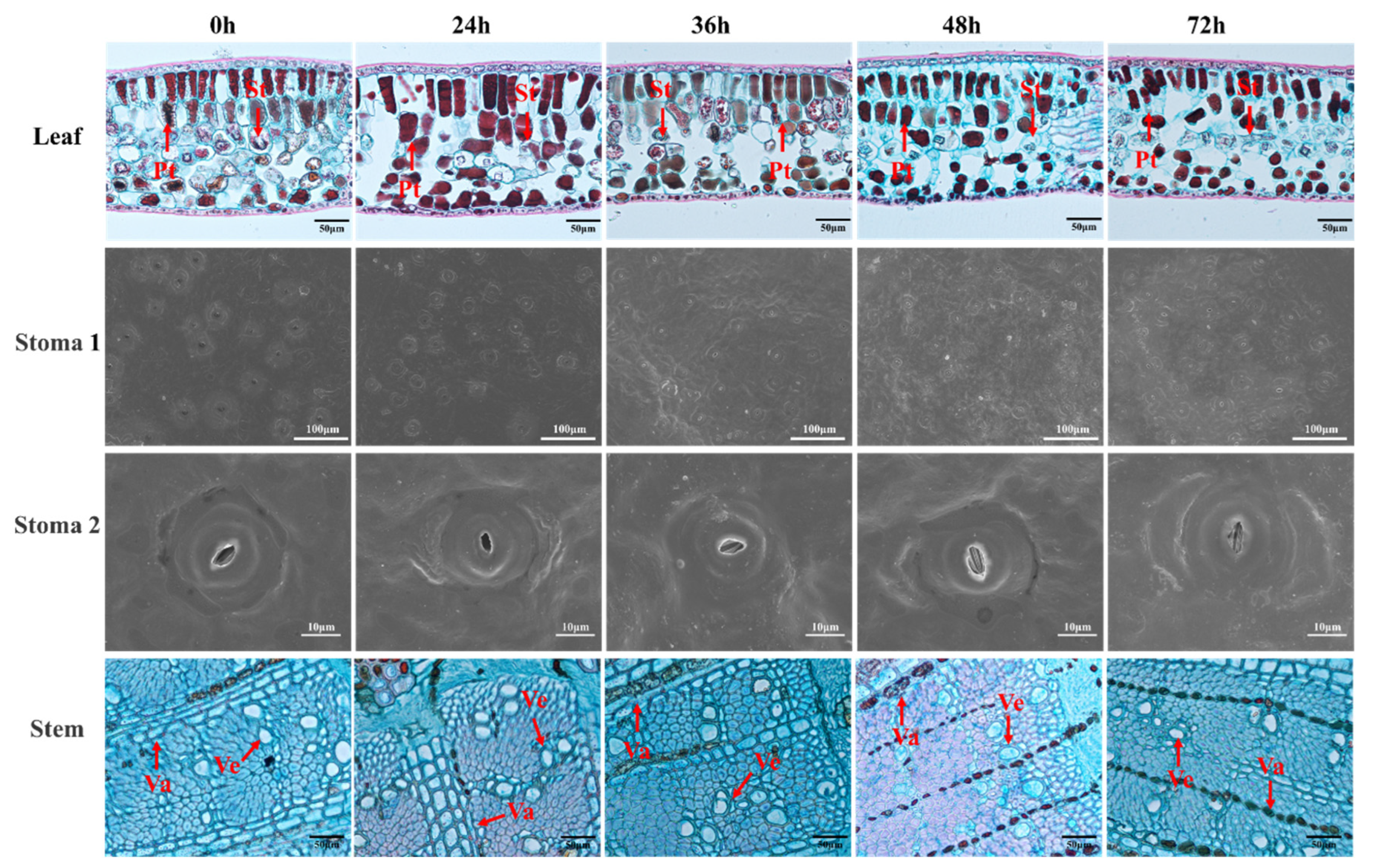
2.2.2. Observation of Leaf and Stem Structure
2.2.3. Measurement of Physiological and Biochemical Indicators
2.3. Statistical Analysis of the Data
3. Results
3.1. Effects of Drought Stress on the Morphology of Macadamia Seedlings
3.2. Effects of Drought Stress on Water Potential, Relative Water Content, Root Activity, Conductivity and MDA of Macadamia Seedings
3.3. Effects of Drought Stress on the Photosynthetic Characteristics of Macadamia Seedling Leaves
3.4. Effect of Drought Stress on Hormones in Macadamia Seedlings
3.5. Effects of Drought Stress on Osmoregulatory Substances in Macadamia Seedlings
3.6. Effects of Drought Stress on Antioxidant Activity in Macadamia Seedlings
4. Discussion
4.1. Effects of Drought Stress on Anatomy and Physiology
4.2. Effects of Drought Stress on Photosynthetic Characteristics
4.3. Effect of Drought Stress on Hormone Levels
4.4. Effects of Drought Stress on Osmoregulatory Substances
4.5. Effects of Drought Stress on Antioxidant Activity
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagao, M.A.; Hirae, H.H.; Stephenson, R.A. Macadamia: Cultivation and physiology. Crit. Rev. Plant Sci. 1992, 10, 441–470. [Google Scholar] [CrossRef]
- Venkatachalam, M.; Sathe, S.K. Chemical composition of selected edible nut seeds. J. Agric. Food Chem. 2006, 54, 4705–4714. [Google Scholar] [CrossRef] [PubMed]
- Shuai, X.; Dai, T.; Chen, M.; Liang, R.; Du, L.; Chen, J.; Liu, C. Comparative study of chemical compositions and antioxidant capacities of oils obtained from 15 macadamia (Macadamia integrifolia) cultivars in China. Foods 2021, 10, 1031. [Google Scholar] [CrossRef]
- Xia, C.; Jiang, S.; Tan, Q.; Wang, W.; Zhao, L.; Zhang, C.; Bao, Y.; Liu, Q.; Wang, W.; Zou, M.; et al. Chromosomal-level genome of macadamia (Macadamia integrifolia). Trop. Plants 2022, 1, 3. [Google Scholar] [CrossRef]
- Tu, X.H.; Wu, B.; Xie, Y.; Xu, L.; Wu, Z.; Lv, X.; Wei, F.; Du, L.; Chen, H. A comprehensive study of raw and roasted macadamia nuts: Lipid profile, physicochemical, nutritional, and sensory properties. Food Sci. Nutr. 2021, 9, 1688–1697. [Google Scholar] [CrossRef]
- Kang, Z.M.; He, F.P.; Geng, J.J.; Zeng, H.; Tu, X.; Wang, W. Evaluation of quality characters of Macadamia suitable for planting in Guizhou. Non-Wood For. Res. 2020, 38, 117–124. [Google Scholar]
- Yang, M.; Mou, Y.; Meng, Y.; Liu, S.; Peng, C.; Zhou, X. Modeling the effects of precipitation and temperature patterns on agricultural drought in China from 1949 to 2015. Sci. Total Environ. 2020, 711, 135139. [Google Scholar] [CrossRef]
- Wang, C.L.; Zhong, S.B.; Yao, G.N.; Huang, Q.Y. BME spatiotemporal estimation of annual precipitation and detection of drought hazard clusters using space–time scan statistics in the Yun-Gui-Guang region, Mainland China. J. Appl. Meteorol. Climatol. 2017, 56, 2301–2316. [Google Scholar] [CrossRef]
- Kiem, A.S.; Johnson, F.; Westra, S.; Dijk, A.; Rouillard, A.; Sivakumar, B.; Mehrotra, R. Natural hazards in Australia: Droughts. Clim. Chang. 2016, 139, 37–54. [Google Scholar] [CrossRef]
- Wilhite, D.A.; Sivakumar MV, K.; Pulwarty, R. Managing drought risk in a changing climate: The role of national drought policy. Weather. Clim. Extrem. 2014, 3, 4–13. [Google Scholar] [CrossRef]
- Moore, C.E.; Meacham-Hensold, K.; Lemonnier, P.; Slattery, R.A.; Benjamin, C.; Bernacchi, C.J.; Lawson, T.; Cavanagh, A.P. The effect of increasing temperature on crop photosynthesis: From enzymes to ecosystems. J. Exp. Bot. 2021, 72, 2822–2844. [Google Scholar] [CrossRef]
- Zahra, N.; Hafeez, M.B.; Ghaffar, A.; Zeidi, M.A.; Siddique, K.H.M.; Farooq, M. Plant photosynthesis under heat stress: Effects and management. Environ. Exp. Bot. 2023, 206, 105178. [Google Scholar] [CrossRef]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, F.; Kuromori, T.; Urano, K.; Shinozaki, K.Y.; Shinozaki, K. Drought stress responses and resistance in plants: From cellular responses to long-distance intercellular communication. Front. Plant Sci. 2020, 11, 556972. [Google Scholar] [CrossRef]
- Urano, K.; Maruyama, K.; Ogata, Y.; Morishita, Y.; Takeda, M.; Sakurai, N.; Suzuki, H.; Saito, K.; Shibata, D.; Kobayashi, M. Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics. Plant J. 2009, 57, 1065–1078. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R. Plant responses to multifactorial stress combination. New Phytol. 2022, 234, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional genomics in plant abiotic stress responses and tolerance: From gene discovery to complex regulatory networks and their application in breeding. Proc. Jpn. Acad. Ser. B 2022, 98, 470–492. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef]
- Shi, H.; Chen, L.; Ye, T.; Liu, X.; Ding, K.; Chan, Z. Modulation of auxin content in Arabidopsis confers improved drought stress resistance. Plant Physiol. Biochem. 2014, 82, 209–217. [Google Scholar] [CrossRef]
- Rivero, R.M.; Shulaev, V.; Blumwald, E. Cytokinin-dependent photorespiration and the protection of photosynthesis during water deficit. Plant Physiol. 2009, 150, 1530–1540. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef]
- Rademacher, W. Growth retardants: Effects on gibberellin biosynthesis and other metabolic pathways. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 501–531. [Google Scholar] [CrossRef]
- Stephenson, R.A.; Gallagher, E.C.; Doogan, V.J.; Mayer, D.G. Nitrogen and environmental factors influencing macadamia quality. Aust. J. Exp. Agric. 2000, 40, 1145–1150. [Google Scholar] [CrossRef]
- Emmanuel, J.Z.; Yoseph, N.A.; Kadmiel, M. Climatic suitability predictions for the cultivation of macadamia in Malawi using climate change scenarios. bioRxiv 2021. [Google Scholar] [CrossRef]
- Bringhenti, T.; Joubert, E.; Abdulai, I.; Hoffmann, M.P.; Moriondo, M.; Taylor, P.J.; Roetter, R.P. Effects of environmental drivers and irrigation on yields of macadamia orchards along an altitudinal gradient in South Africa. Sci. Hortic. 2023, 321, 112326. [Google Scholar] [CrossRef]
- Theunis, G.S.; Nicolette, J.T.; Stephanie, J.E.M. The seasonal regulation of gas exchange and water relations of field grown macadamia. Sci. Hortic. 2020, 267, 109346. [Google Scholar] [CrossRef]
- Mlungisi, S.; Michele, T.; Alistair, C. The Macadamia bloom—What are the hydrological implications? Sci. Hortic. 2022, 292, 110628. [Google Scholar] [CrossRef]
- Kang, Z.M.; Zhang, W.; Guo, G.Z.; Pan, X.J.; Huang, D.; Wang, R.; Shen, X.J. Morphological and physiological responses of 14 macadamia rootstocks to drought stress and a comprehensive evaluation of drought resistance. Environ. Exp. Bot. 2024, 219, 105630. [Google Scholar] [CrossRef]
- Yao, X.C.; Meng, L.F.; Zhao, W.L.; Mao, G.L. Changes in the morphology traits, anatomical structure of the leaves and transcriptome in Lycium barbarum L. under salt stress. Front. Plant Sci. 2023, 14, 1090366. [Google Scholar] [CrossRef]
- Matthaeus, W.J.; Schmidt, J.; White, J.D.; Zechmann, B. Novel perspectives on stomatal impressions: Rapid and non-invasive surface characterization of plant leaves by scanning electron microscopy. PLoS ONE 2020, 15, e0238589. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Hou, K.; Zhang, H.; Wang, X.; Wu, W. Integrating transcriptomics and metabolomics to studies key metabolism, pathways and candidate genes associated with drought-tolerance in Carthamus tinctorius L. under drought stress. Ind. Crops Prod. 2020, 151, 112465. [Google Scholar] [CrossRef]
- Barranco, D.; Ruiz, N.; Gomes, M. Frost tolerance of eight olive cultivars. HortScience 2005, 40, 558–560. [Google Scholar] [CrossRef]
- Xue, W.; Liu, W.; Ma, R.; Zhang, S.H.; Yu, X.; Li, T.; Luan, X.Y.; Cui, X.W.; Liu, J.; Zhang, C.W. The toxic mechanism of tetracycline on root tips in hulless barley (Hordeum vulgare L. var. nudum). J. Hazard. Mater. 2023, 460, 132453. [Google Scholar] [CrossRef]
- Zhang, Q.; Xing, D.; Wu, Y.; Zhao, K.; Wang, J.; Mao, R. Effects of Low-Phosphorus Stress on Use of Leaf Intracellular Water and Nutrients, Photosynthesis, and Growth of Brassica napus L. Horticulturae 2024, 10, 821. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, Y.; Wang, X.; Yan, C.; Ma, C.; Liu, J.; Dong, S. Effects of Different Drought Degrees on Physiological Characteristics and Endogenous Hormones of Soybean. Plants 2022, 11, 2282. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, C.; Kuang, J.; Ge, Q.; Zhang, Y.; Wang, Z. Overexpression of SmLEA enhances salt and drought tolerance in Escherichia coli and Salvia miltiorrhiza. Protoplasma 2014, 251, 1191–1199. [Google Scholar] [CrossRef]
- Skriver, K.; Mundy, J. Gene expression in response to abscisic acid and osmotic stress. Plant Cell 1990, 2, 503. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Karimi, M.; Venditti, A.; Zahra, N.; Siddique, K.H.M.; Farooq, M. Plant Adaptation to Drought Stress: The Role of Anatomical and Morphological Characteristics in Maintaining the Water Status. J. Soil Sci. Plant Nutr. 2024, 25, 409–427. [Google Scholar] [CrossRef]
- Zhu, J.C.; Cai, D.F.; Wang, J.P.; Cao, J.H.; Wen, Y.C.; He, J.P.; Zhao, L.; Wang, D.G.; Zhang, S.F. Physiological and anatomical changes in two rapeseed (Brassica napus L.) genotypes under drought stress conditions. Oil Crop Sci. 2021, 6, 97–104. [Google Scholar] [CrossRef]
- Haffani, S.; Mezni, M.; Nasri, M.B.; Chaibi, W. Comparative leaf water relations and anatomical responses of three vetch species (Vicia narbonensis L., V. sativa L. and V. villosa Roth.) to cope with water stress. Crop Pasture Sci. 2017, 68, 691–702. [Google Scholar] [CrossRef]
- Zimmermann, J.; Link, R.M.; Hauck, M.; Leuschner, C.; Schuldt, B. 60-year record of stem xylem anatomy and related hydraulic modification under increased summer drought in ring-and diffuse-porous temperate broad-leaved tree species. Trees 2021, 35, 919–937. [Google Scholar] [CrossRef]
- Weber AP, M.; Bar-Even, A. Update: Improving the efficiency of photosynthetic carbon reactions. Plant Physiol. 2019, 179, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.H.; Xu, X.F.; Sun, Y.M.; Zhang, J.L.; Li, C.Z. Influence of drought hardening on the resistance physiology of potato seedlings under drought stress. J. Integr. Agric. 2018, 17, 336–347. [Google Scholar] [CrossRef]
- Trabelsi, L.; Gargouri, K.; Hassena, A.B.; Trabelsi, L.; Gargouri, K.; Hassena, A.B.; Mbadra, C.; Ghrab, M.; Ncube, B.; Staden, V.J. Impact of drought and salinity on olive water status and physiological performance in an arid climate. Agric. Water Manag. 2019, 213, 749–759. [Google Scholar] [CrossRef]
- Liu, D.; Guo, H.L.; Yan, L.P.; Cao, L.; Zhai, S.S.; Xu, Y. Physiological, Photosynthetic and Stomatal Ultrastructural Responses of Quercus acutissima Seedlings to Drought Stress and Rewatering. Forests 2023, 15, 71. [Google Scholar] [CrossRef]
- Arunyanark, A.; Jogloy, S.; Akkasaeng, C.; Vorasoot, N.; Kesmala, R.C.; Nageswara, R.; Wright, G.C.; Patanothai, A. Chlorophyll stability is an indicator of drought tolerance in peanut. J. Agron. Crop Sci. 2008, 194, 113–125. [Google Scholar] [CrossRef]
- Huang, B.; Chen, Y.E.; Zhao, Y.Q.; Ding, C.B.; Liao, J.Q.; Hu, C.; Zhou, L.J.; Zhang, Z.W.; Yuan, S.; Yuan, M. Exogenous melatonin alleviates oxidative damages and protects photosystem II in maize seedlings under drought stress. Front. Plant Sci. 2019, 10, 677. [Google Scholar] [CrossRef]
- Peterson, R.B.; Sivak, M.N.; Walker, D.A. Relationship between steady-state fluorescence yield and photosynthetic efficiency in spinach leaf tissue. Plant Physiol. 1988, 88, 158–163. [Google Scholar] [CrossRef]
- Guo, C.F.; Sun, Y.; Tang, Y.H.; Zhang, M.G. Effect of water stress on chlorophyll fluorescence in leaves of tea plant (Camellia sinensis). Chin. J. Eco-Agric. 2009, 17, 560–564. [Google Scholar] [CrossRef]
- Gilmore, A.M.; Yamamoto, H.Y. Zeaxanthin formation and energy-dependent fluorescence quenching in pea chloroplasts under artificially mediated linear and cyclic electron transport. Plant Physiol. 1991, 96, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Demmig-Adams, B.; Adams, W.W. An integrative approach to photoinhibition and photoprotection of photosynthesis. Environ. Exp. Bot. 2018, 154, 1–3. [Google Scholar] [CrossRef]
- Epron, D.; Dreyer, E. Effects of severe dehydration on leaf photosynthesis in Quercus petraea (Matt.) Liebl.: Photosystem II efficiency, photochemical and nonphotochemical fluorescence quenching and electrolyte leakage. Tree Physiol. 1992, 10, 273–284. [Google Scholar] [CrossRef]
- Havaux, M.; Strasser, R.J.; Greppin, H. A theoretical and experimental analysis of the q P and q N coefficients of chlorophyll fluorescence quenching and their relation to photochemical and nonphotochemical events. Photosynth. Res. 1991, 27, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Magdaong NC, M.; Blankenship, R.E. Photoprotective, excited-state quenching mechanisms in diverse photosynthetic organisms. J. Biol. Chem. 2018, 293, 5018–5025. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Mizoi, J.; Shinozaki, K.; Shinozaki, K.Y. Complex plant responses to drought and heat stress under climate change. Plant J. 2024, 117, 1873–1892. [Google Scholar] [CrossRef]
- Barcia, R.A.; Pena, L.B.; Zawoznik, M.S.; Benavides, M.P.; Gallego, S.M. Osmotic adjustment and maintenance of the redox balance in root tissue may be key points to overcome a mild water deficit during the early growth of wheat. Plant Growth Regul. 2014, 74, 107–117. [Google Scholar] [CrossRef]
- Lintunen, A.; Paljakka, T.; Jyske, T.; Peltoniemi, M.; Sterck, F.; Arx, G.V.; Cochard, H.; Copini, P.; Maria, C.C.; Sylvain, D. Osmolality and non-structural carbohydrate composition in the secondary phloem of trees across a latitudinal gradient in Europe. Front. Plant Sci. 2016, 7, 726. [Google Scholar] [CrossRef]
- Hein, J.A.; Sherrard, M.E.; Manfredi, K.P.; Abebe, T. The fifth leaf and spike organs of barley (Hordeum vulgare L.) display different physiological and metabolic responses to drought stress. BMC Plant Biol. 2016, 16, 248. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Ashraf, U.; Hussain, S.; Shahzad, B.; Khan, I.; Wang, L. Effect of progressive drought stress on growth, leaf gas exchange, and antioxidant production in two maize cultivars. Environ. Sci. Pollut. Res. 2016, 23, 17132–17141. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, L.; Garnczarska, M. Contribution of exogenous proline to abiotic stresses tolerance in plants: A review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Ahanger, M.A.; Su, Y.Y.; Lei, Y.F.; Nabil, S.A.; Mustafa, N.S.A.; Ahmad, P.; Zhang, L.X. Improved drought tolerance by AMF inoculation in maize (Zea mays) involves physiological and biochemical implications. Plants 2019, 8, 579. [Google Scholar] [CrossRef] [PubMed]
- Zegaoui, Z.; Planchais, S.; Cabassa, C.; Djebbar, R.; Belbachir, O.A.; Carol, P. Variation in relative water content, proline accumulation and stress gene expression in two cowpea landraces under drought. J. Plant Physiol. 2017, 218, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Khoyerdi, F.F.; Shamshiri, M.H.; Estaji, A. Changes in some physiological and osmotic parameters of several pistachio genotypes under drought stress. Sci. Hortic. 2016, 198, 44–51. [Google Scholar] [CrossRef]
- Kishor PB, K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Ahmed, C.B.; Rouina, B.B.; Sensoy, S.; Boukhris, M.; Abdallah, F.B. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Slabbert, M.M.; Krüger, G.H.J. Antioxidant enzyme activity, proline accumulation, leaf area and cell membrane stability in water stressed Amaranthus leaves. S. Afr. J. Bot. 2014, 95, 123–128. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Emam, Y.; Pessarakli, M. Changes in endogenous hormonal status in corn (Zea mays) hybrids under drought stress. J. Plant Nutr. 2013, 36, 1695–1707. [Google Scholar] [CrossRef]
- Zhao, Y. Auxin biosynthesis and its role in plant development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef]
- Kurepa, J.; Smalle, J.A. Auxin/Cytokinin Antagonistic Control of the Shoot/Root Growth Ratio and Its Relevance for Adaptation to Drought and Nutrient Deficiency Stresses. Int. J. Mol. Sci. 2022, 23, 1933. [Google Scholar] [CrossRef]
- Nir, I.; Moshelion, M.; Weiss, D. The Arabidopsis gibberellin methyl transferase 1 suppresses gibberellin activity, reduces whole-plant transpiration and promotes drought tolerance in transgenic tomato. Plant Cell Environ. 2014, 37, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Furutera, A.; Seki, K.; Toyoda, Y.; Tanaka, K.; Sugimoto, Y. Malondialdehyde generated from peroxidized linolenic acid causes protein modification in heat-stressed plants. Plant Physiol. Biochem. 2008, 46, 786–793. [Google Scholar] [CrossRef]
- Miller, G.A.D.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Chang-Quan, W.; Rui-Chang, L. Enhancement of superoxide dismutase activity in the leaves of white clover (Trifolium repens L.) in response to polyethylene glycol-induced water stress. Acta Physiol. Plant. 2008, 30, 841–847. [Google Scholar] [CrossRef]
- Sekmen, A.H.; Ozgur, R.; Uzilday, B.; Turkan, I. Reactive oxygen species scavenging capacities of cotton (Gossypium hirsutum) cultivars under combined drought and heat induced oxidative stress. Environ. Exp. Bot. 2014, 99, 141–149. [Google Scholar] [CrossRef]
- Gokce, A.; Sekmen Cetinel, A.H.; Turkan, I. Involvement of GLR-mediated nitric oxide effects on ROS metabolism in Arabidopsis plants under salt stress. J. Plant Res. 2024, 137, 485–503. [Google Scholar] [CrossRef]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic stress tolerance in plants: Myriad roles of ascorbate peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef]
- Wang, N.; Liu, W.J.; Yu, L.; Guo, Z.W.; Chen, Z.J.; Jiang, S.H.; Xu, H.F.; Fang, H.C.; Wang, Y.C.; Zhang, Z.Y. HEAT SHOCK FACTOR A8a modulates flavonoid synthesis and drought tolerance. Plant Physiol. 2020, 184, 1273–1290. [Google Scholar] [CrossRef]
- Rasheed, F.; Gondal, A.; Kudus, K.A.; Zafar, Z.; Nawaz, M.F.; Khan, W.R.; Abdullah, M.; Ibrahim, F.H.; Depardieu, C.; Pazi, A.M.M. Effects of soil water deficit on three tree species of the arid environment: Variations in growth, physiology, and antioxidant enzyme activities. Sustainability 2021, 13, 3336. [Google Scholar] [CrossRef]
- Denaxa, N.K.; Damvakaris, T.; Roussos, P.A. Antioxidant defense system in young olive plants against drought stress and mitigation of adverse effects through external application of alleviating products. Sci. Hortic. 2020, 259, 108812. [Google Scholar] [CrossRef]
- Dreyer, A.; Dietz, K.J. Reactive oxygen species and the redox-regulatory network in cold stress acclimation. Antioxidants 2018, 7, 169. [Google Scholar] [CrossRef]
- Qi, Y.; Ma, L.; Ghani, M.I.; Peng, Q.; Fan, R.D.; Hu, X.J.; Chen, X.Y.L. Effects of drought stress induced by hypertonic polyethylene glycol (PEG-6000) on Passiflora edulis sims physiological properties. Plants 2023, 12, 2296. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yang, L.; Yang, X.; Zhang, T.; Lan, Y.M.; Zhao, Y.; Han, M.; Yang, L.M. Drought stress induces biosynthesis of flavonoids in leaves and saikosaponins in roots of Bupleurum chinense DC. Phytochemistry 2020, 177, 112434. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.L.; Wang, Y.; Xie, H.; Qiu, C.; Zhang, S.N.; Xiao, J.; Li, H.Y.; Chen, L.; Xinghui Li, X.H.; Ding, Z.T. Drought stress triggers proteomic changes involving lignin, flavonoids and fatty acids in tea plants. Sci. Rep. 2020, 10, 15504. [Google Scholar] [CrossRef]
- Li, P.; Ren, G.X.; Wu, F.; Chen, J.X.; Jiang, D.; Liu, C.S. Root-specific flavones and critical enzyme genes involved in their synthesis changes due to drought stress on Scutellaria baicalensis. Front. Ecol. Evol. 2023, 11, 1113823. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Time | Leaf μm | Palisade Tissue/ μm | Spongy Tissue/ μm | Stomatal Density/ Number/mm2 | Stoma Length/ μm | Stoma Width/ μm | Vessel Diameter/ μm | Wall Thickness/ μm |
|---|---|---|---|---|---|---|---|---|
| 0 h | 218.24 ± 2.14 a | 92.06 ± 6.11 a | 114.67 ± 3.54 a | 163.78 ± 15.15 d | 4.55 ± 0.37 b | 1.98 ± 0.11 b | 15.51 ± 0.51 b | 4.31 ± 0.06 bc |
| 24 h | 214.35 ± 3.89 a | 89.91 ± 1.98 a | 102.54 ± 7.24 b | 190.43 ± 6.14 c | 3.58 ± 0.19 c | 2.36 ± 0.16 a | 18.83 ± 0.89 a | 4.51 ± 0.12 b |
| 36 h | 192.25 ± 7.59 b | 74.33 ± 4.16 b | 92.38 ± 6.44 c | 206.74 ± 22.63 bc | 3.28 ± 0.07 c | 1.92 ± 0.04 bc | 19.24 ± 1.20 a | 4.97 ± 0.12 a |
| 48 h | 188.36 ± 2.57 b | 71.35 ± 3.51 b | 87.60 ± 3.58 cd | 228.47 ± 14.93 ab | 4.83 ± 0.38 ab | 1.70 ± 0.14 c | 16.58 ± 0.34 b | 4.53 ± 0.29 b |
| 72 h | 175.71 ± 6.43 c | 61.45 ± 1.42 c | 81.97 ± 3.13 d | 245.63 ± 4.73 a | 5.33 ± 0.47 a | 1.16 ± 0.19 d | 15.80 ± 0.70 b | 3.99 ± 0.30 c |
| Time | Chlorophyll mg/g | Pn μmol/m2·s | Tr μmol/m2·s | Gs mol/m2·s | Ci μmol/mol | WUE % | Fv/Fm | qP | qN | ΦPSII |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 2.50 ± 0.11 a | 13.53 ± 0.43 a | 3.23 ± 0.03 a | 0.15 ± 0.03 a | 183.30 ± 11.23 d | 4.31 ± 0.33 a | 0.67 ± 0.02 a | 0.83 ± 0.03 a | 0.80 ± 0.01 b | 0.32 ± 0.02 a |
| 24 h | 2.09 ± 0.01 b | 10.52 ± 0.33 b | 2.92 ± 0.05 ab | 0.09 ± 0.00 b | 215.33 ± 12.76 c | 4.05 ± 0.14 a | 0.61 ± 0.06 ab | 0.79 ± 0.04 a | 0.82 ± 0.06 b | 0.24 ± 0.04 b |
| 48 h | 1.62 ± 0.05 c | 7.65 ± 0.34 c | 2.72 ± 0.09 bc | 0.08 ± 0.01 bc | 337.37 ± 22.46 b | 2.95 ± 0.01 b | 0.51 ± 0.01 c | 0.59 ± 0.01 b | 0.88 ± 0.01 a | 0.11 ± 0.01 c |
| 72 h | 1.40 ± 0.03 d | 3.63 ± 0.36 d | 2.42 ± 0.29 c | 0.05 ± 0.01 c | 378.77 ± 5.60 a | 1.57 ± 0.25 c | 0.44 ± 0.02 d | 0.33 ± 0.03 c | 0.89 ± 0.00 a | 0.05 ± 0.01 d |
| Time | IAA ng/g FW | CTK ng/g FW | GA ng/g FW | ABA ng/g FW |
|---|---|---|---|---|
| 0 h | 763.00 ± 5.30 a | 306.89 ± 12.16 a | 434.99 ± 4.14 a | 502.46 ± 10.76 e |
| 24 h | 666.02 ± 4.99 b | 236.54 ± 9.16 b | 418.29 ± 24.08 a | 583.01 ± 14.26 d |
| 36 h | 584.01 ± 5.03 c | 230.04 ± 4.37 b | 380.83 ± 18.28 b | 642.72 ± 4.39 c |
| 48 h | 544.30 ± 9.95 d | 224.26 ± 4.26 b | 382.64 ± 12.03 b | 706.83 ± 21.56 b |
| 72 h | 483.36 ± 5.30 e | 206.83 ± 7.18 c | 365.04 ± 12.92 b | 742.96 ± 11.68 a |
| Time | SS mg/g | SP mg/g | Pro ug/g FW | BA ug/g FW |
|---|---|---|---|---|
| 0 h | 22.69 ± 0.20 d | 24.77 ± 0.75 e | 122.51 ± 1.91 e | 101.94 ± 2.28 e |
| 24 h | 24.08 ± 1.40 d | 30.93 ± 0.37 d | 389.44 ± 6.85 d | 136.72 ± 2.35 d |
| 36 h | 27.35 ± 1.01 c | 36.10 ± 0.64 c | 647.38 ± 6.38 c | 184.71 ± 2.67 c |
| 48 h | 32.78 ± 0.62 b | 37.99 ± 0.75 b | 818.05 ± 5.78 b | 245.19 ± 2.96 b |
| 72 h | 36.45 ± 0.31 a | 42.74 ± 0.34 a | 975.64 ± 4.87 a | 340.79 ± 4.71 a |
| Time | SOD (U/g FW) | POD (U/g FW) | CAT (U/g FW) | APX (U/g FW) | Total Flavonoids (mg/g FW) |
|---|---|---|---|---|---|
| 0 h | 63.40 ± 0.80 c | 29,108.86 ± 545.79 e | 286.39 ± 4.12 e | 1.66 ± 0.04 c | 11.66 ± 0.32 a |
| 24 h | 146.78 ± 3.73 a | 112,988.12 ± 1679.93 a | 601.91 ± 10.91 a | 2.42 ± 0.07 a | 8.26 ± 0.06 bc |
| 36 h | 94.38 ± 2.33 b | 104,194.50 ± 4206.33 b | 550.07 ± 2.45 b | 2.46 ± 0.09 a | 8.27 ± 0.11 bc |
| 48 h | 57.75 ± 1.13 d | 92,450.27 ± 1561.88 c | 436.53 ± 4.22 c | 2.09 ± 0.10 b | 7.96 ± 0.21 c |
| 72 h | 25.71 ± 1.36 e | 79,067.91 ± 1843.06 d | 346.84 ± 11.18 d | 1.51 ± 0.01 d | 8.55 ± 0.32 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Z.; Cai, H.; Guo, G.; Zeng, H.; Wang, W.; Tu, X. Physiological Response of Macadamia (Macadamia integrifolia) Seedlings to Drought Stress. Horticulturae 2025, 11, 347. https://doi.org/10.3390/horticulturae11040347
Kang Z, Cai H, Guo G, Zeng H, Wang W, Tu X. Physiological Response of Macadamia (Macadamia integrifolia) Seedlings to Drought Stress. Horticulturae. 2025; 11(4):347. https://doi.org/10.3390/horticulturae11040347
Chicago/Turabian StyleKang, Zhuanmiao, Hu Cai, Guangzheng Guo, Hui Zeng, Wenlin Wang, and Xinghao Tu. 2025. "Physiological Response of Macadamia (Macadamia integrifolia) Seedlings to Drought Stress" Horticulturae 11, no. 4: 347. https://doi.org/10.3390/horticulturae11040347
APA StyleKang, Z., Cai, H., Guo, G., Zeng, H., Wang, W., & Tu, X. (2025). Physiological Response of Macadamia (Macadamia integrifolia) Seedlings to Drought Stress. Horticulturae, 11(4), 347. https://doi.org/10.3390/horticulturae11040347