
Effects of Light Conditions on the Leaf Growth and Steviol Glycoside Yields of Hydroponically Cultivated Stevia Across Growth Stages



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
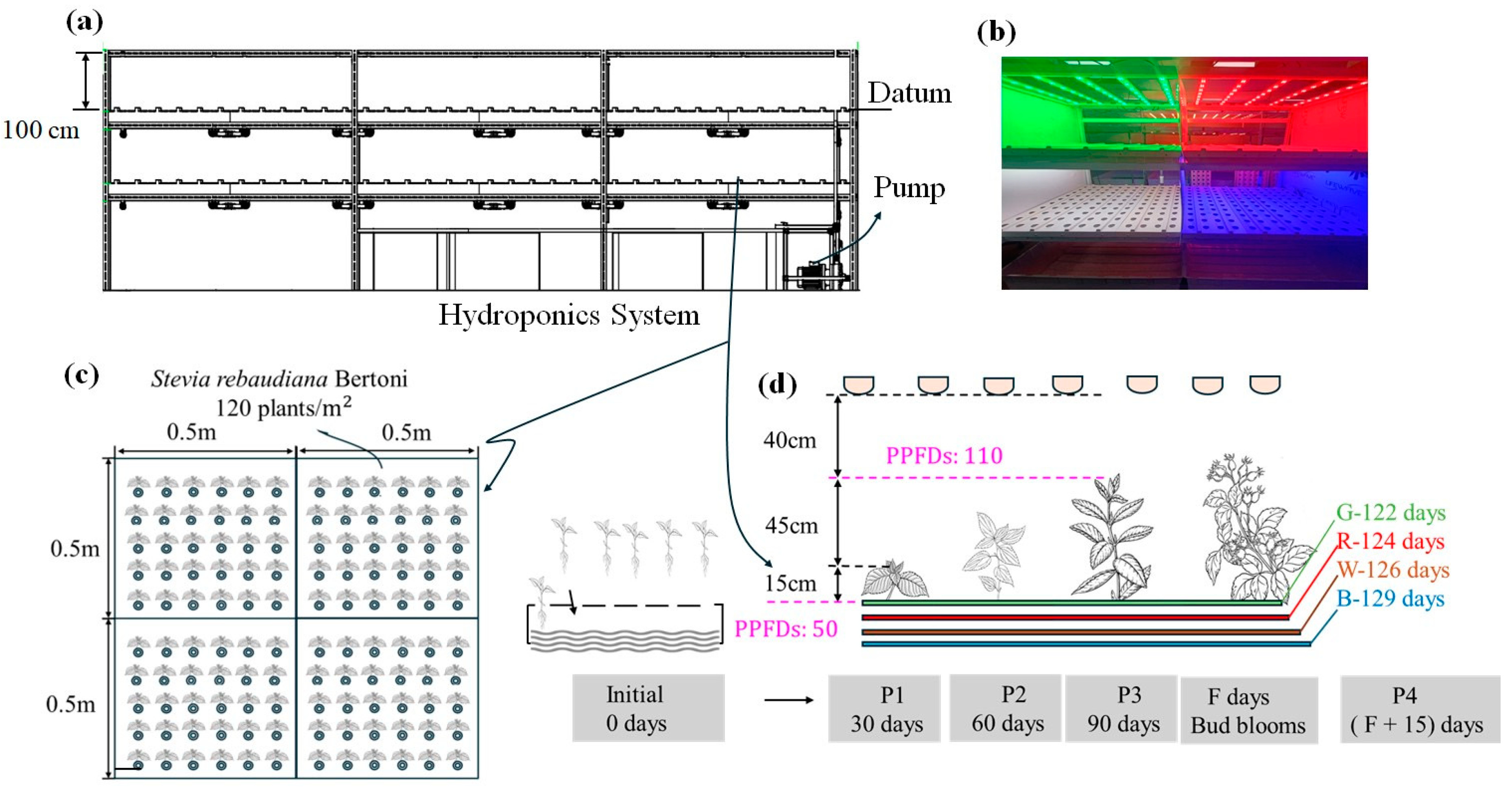
2.2. Culture System
2.3. Culture and Light Treatments
2.4. Microwave Extraction Process
2.5. Determination of Stevia Extraction Yield
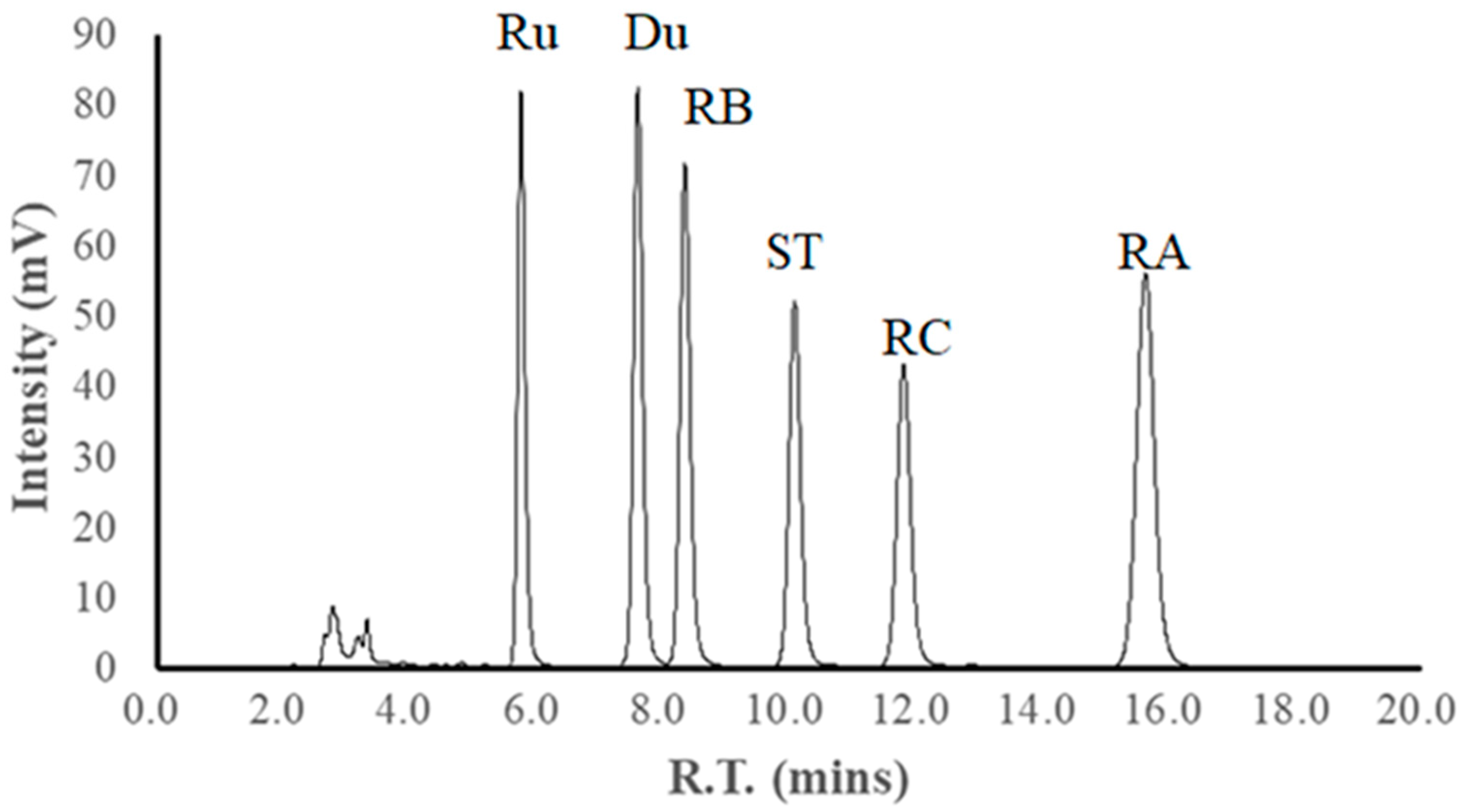
2.6. Content and Yield of Steviol Glycosides
- C: Concentration of each SG component determined by HPLC (mg·mL−1)
- V: Volume of the prepared stevia extract lyophilizate solution (mL)
- LE: Weight of the lyophilized stevia extract (g)
- HRu, HDu, HRB, HST, HRC, and HRA represent the content of Ru, Du, RB, ST, RC, and RA in the lyophilized extract, respectively.
2.7. Statistical Analysis
3. Results and Discussion
3.1. Effects of Light Quality on Biomass
3.1.1. Stevia Leaves Fresh Weight (FW)
3.1.2. Stevia Leaves Dry Weight (DW)
3.2. Effects of Different Light Qualities and Growth Stages on Stevia Extraction Yield
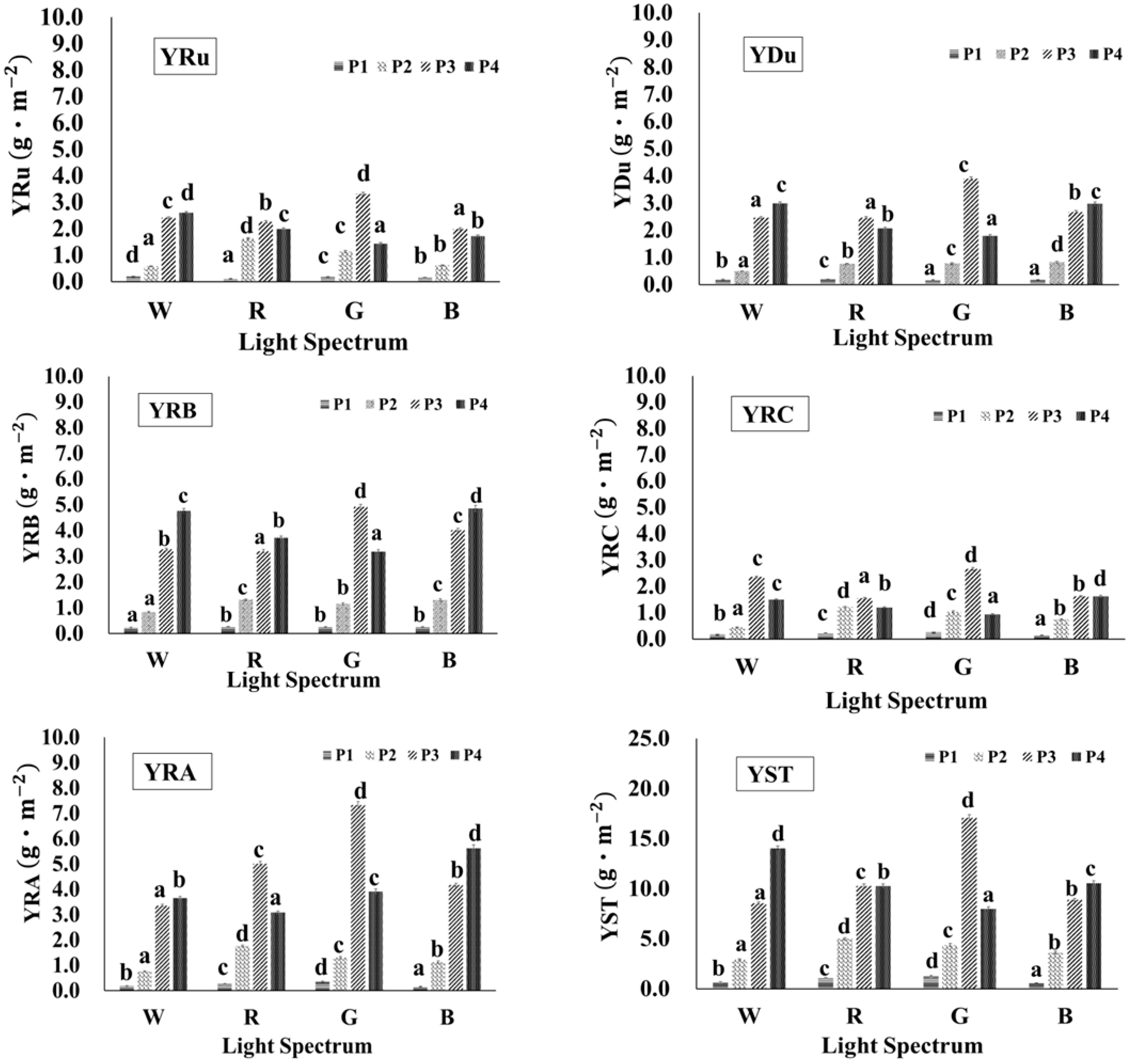
3.3. Effects of Different Light Qualities on the Content of SG in Stevia Leaves
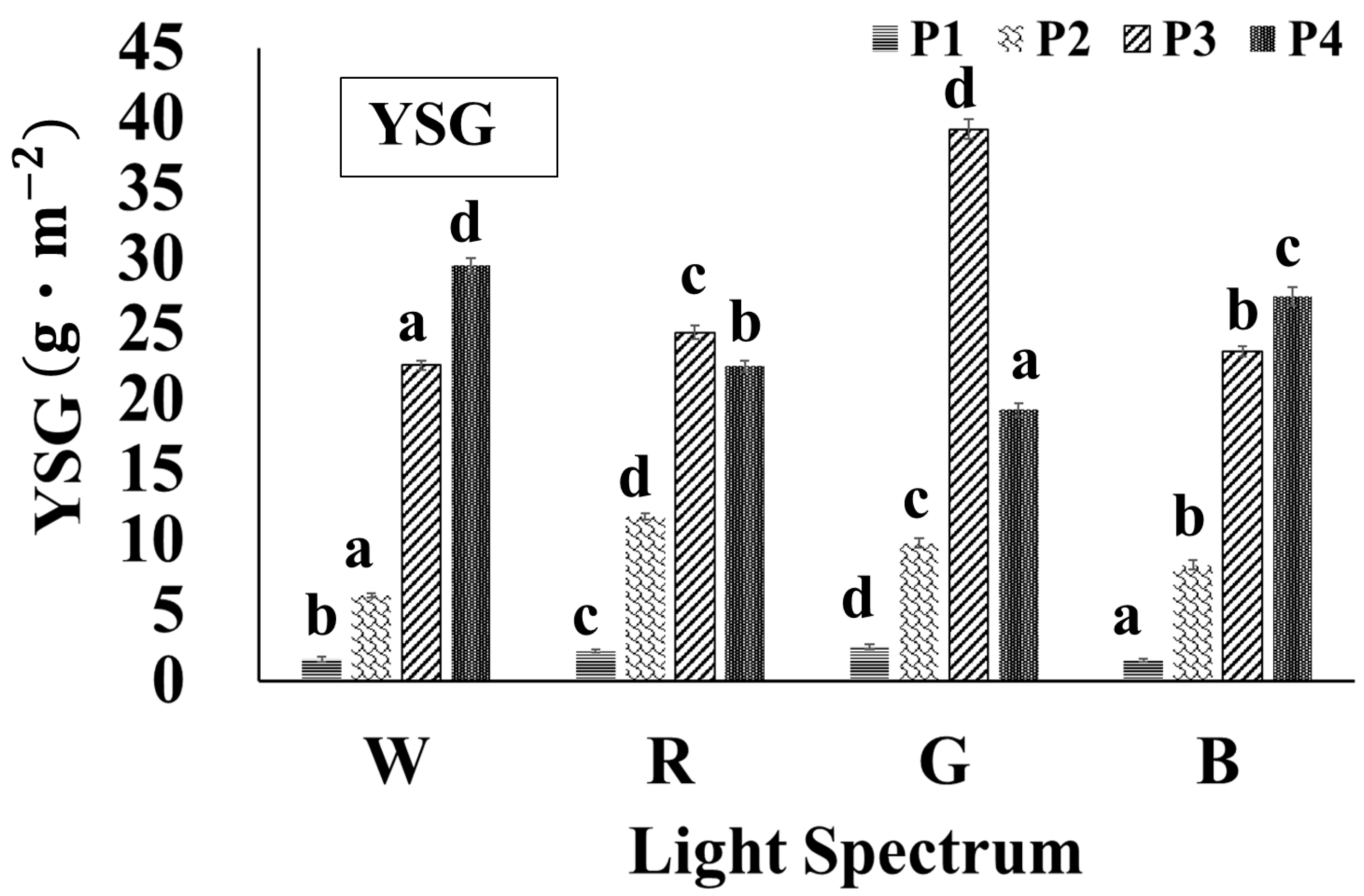
3.4. Effects of Different Light Qualities on the Yield of SG
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| CRYs | Cryptochrome family |
| D | Planting density (plants·m−2) |
| DW | Stevia leaves dry weight (g·plant−1) |
| E | Extraction yield of the stevia extract (%) |
| EC | Electrical conductivity (mS·cm−1) |
| F | The number of days required for the first flower bud to appear (days) |
| FW | Stevia leaves fresh weight (g·plant−1) |
| HPLC | High-performance liquid chromatography |
| KA13H | Ent-kaurenoic acid 13-hydroxylase |
| LE | The weight of the lyophilized stevia leaf extract (g) |
| LED | Light-emitting diode |
| PPFD | Photosynthetic photon flux density (μmol·m−2·s−1) |
| PHOTs | Phototropin family |
| PHYs | Phytochrome family |
| RA–RF | Rebaudioside A–F |
| SP | Stevia powder weight (g) |
| STB | Steviolbioside |
| UGTs | A series of uridine 5′-diphospho-glucuronosyltransferases |
| UVR8 | UV-resistance locus 8 |
| 1st code | |
| Concentration, content, yield | |
| C | Concentration of each SG component determined by HPLC. |
| H | Content of each SG component in the lyophilized stevia extract. |
| L | Content of each SG component in each gram of dried stevia leaf. |
| Y | Yield of each SG component per m2 of cultivation area. |
| 2nd code | |
| Steviol glycosides type | |
| Ru | Rubusoside |
| Du | Dulcoside A |
| RB | Rebaudioside B |
| ST | Stevioside |
| RC | Rebaudioside C |
| RA | Rebaudioside A |
| SG | Total SG |
| 3rd code | |
| Light condition | |
| W | White |
| R | Red |
| G | Green |
| B | Blue |
| 4th code | |
| Cultivation period | |
| P1 | 30 days |
| P2 | 60 days |
| P3 | 90 days |
| P4 | (F + 15) days |
References
- Budeguer, C.J.; Camadro, E.L.; Erazzú, L.E. Reproductive biology and pollen-pistil compatibility relationships in an Argentinian collection of Stevia rebaudiana Bertoni. J. Basic Appl. Genet. 2024, 35, 65–72. [Google Scholar] [CrossRef]
- Basharat, S.; Huang, Z.; Gong, M.; Lv, X.; Ahmed, A.; Hussain, I.; Li, J.; Du, G.; Liu, L. A review on current conventional and biotechnical approaches to enhance biosynthesis of steviol glycosides in Stevia rebaudiana. Chin. J. Chem. Eng. 2021, 30, 92–104. [Google Scholar] [CrossRef]
- Muñoz-Labrador, A.; Hernandez-Hernandez, O.; Moreno, F.J. A review of the state of sweeteners science: The natural versus artificial non-caloric sweeteners debate. Stevia rebaudiana and Siraitia grosvenorii into the spotlight. Crit. Rev. Biotechnol. 2024, 44, 1080–1102. [Google Scholar]
- Watanabe, T.; Fujikawa, K.; Urai, S.; Iwaki, K.; Hirai, T.; Miyagawa, K.; Uratani, H.; Yamagaki, T.; Nagao, K.; Yokoo, Y.; et al. Identification, chemical synthesis, and sweetness evaluation of rhamnose or xylose containing steviol glycosides of stevia (Stevia rebaudiana) leaves. J. Agric. Food Chem. 2023, 71, 11158–11169. [Google Scholar] [CrossRef]
- Tian, X.; Zhong, F.; Xia, Y. Dynamic characteristics of sweetness and bitterness and their correlation with chemical structures for six steviol glycosides. Food Res. Int. 2022, 151, 110848. [Google Scholar] [CrossRef]
- Rivera-Avilez, J.A.; Jarma-Orozco, A.; Pompelli, M.F. Stevia rebaudiana Bertoni: The interaction of night interruption on gas exchange, flowering delay, and steviol glycosides synthesis. Horticulturae 2021, 7, 543. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Zhou, C.; Fan, X.; Sun, Q.; Han, J.; Hua, C.; Li, Y.; Niu, Y.; Okonkwo, C.E.; et al. Properties, extraction and purification technologies of Stevia rebaudiana steviol glycosides: A review. Food Chem. 2024, 453, 139622. [Google Scholar] [CrossRef]
- Zewail, R.M.; Ali, M.; El-Gamal, I.S.; Al-Maracy, S.H.; Islam, K.R.; Elsadek, M.; Azab, E.; Gobouri, A.A.; ElNahhas, N.; Mohamed, M.H.M.; et al. Interactive effects of arbuscular mycorrhizal inoculation with nano boron, zinc, and molybdenum fertilization on stevioside contents of stevia (Stevia rebaudiana L.) plants. Horticulturae 2021, 7, 260. [Google Scholar] [CrossRef]
- Khiraoui, A.; Bakha, M.; Boulli, A.; Hasib, A.; Faiz, C.A. The productivity of Stevia rebaudiana (Bertoni) on dry leaves and steviol glycosides of four varieties grown in six regions of Morocco. Biocatal. Agric. Biotechnol. 2021, 37, 102151. [Google Scholar] [CrossRef]
- Wu, D.; Hu, Q.; Yan, Z.; Chen, W.; Yan, C.; Huang, X.; Zhang, J.; Yang, P.; Deng, H.; Wang, J.; et al. Structural basis of ultraviolet B perception by UVR8. Nature 2012, 484, 214–219. [Google Scholar] [PubMed]
- Paradiso, R.; Proietti, S. Light-quality manipulation to control plant growth and photomorphogenesis in greenhouse horticulture: The state of the art and opportunities of modern LED systems. J. Plant Growth Regul. 2022, 41, 742–780. [Google Scholar] [CrossRef]
- Konrad, K.R.; Gao, S.; Zurbriggen, M.D.; Nagel, G. Optogenetic methods in plant biology. Annu. Rev. Plant Biol. 2023, 74, 313–339. [Google Scholar] [CrossRef] [PubMed]
- Casal, J.J. Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol. 2013, 64, 403–427. [Google Scholar] [CrossRef] [PubMed]
- Lei, K.; Hu, H.; Chang, M.; Sun, C.; Ullah, A.; Yu, J.; Dong, C.; Gao, Q.; Jiang, D.; Cao, W.; et al. A low red/far-red ratio restricts nitrogen assimilation by inhibiting nitrate reductase associated with downregulated TaNR1. 2 and upregulated TaPIL5 in wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2024, 206, 107850. [Google Scholar] [CrossRef] [PubMed]
- D’Amico-Damião, V.; Barreto, R.F.; de Oliveira Garcia, L.F.; Porto, J.S.; de Mello Prado, R.; Carvalho, R.F. Cryptochrome 1a of tomato modulates nutritional deficiency responses. Sci. Hortic. 2022, 291, 110577. [Google Scholar] [CrossRef]
- Battle, M.W.; Jones, M.A. Cryptochromes integrate green light signals into circadian system. Plant Cell Environ. 2020, 43, 16–27. [Google Scholar] [CrossRef]
- Karki, N.; Vergish, S.; Zoltowski, B.D. Cryptochromes: Photochemical and structural insight into magnetoreception. Protein Sci. 2021, 30, 1521–1534. [Google Scholar] [CrossRef]
- Mawphlang, O.I.L.; Kharshiing, E.V. Photoreceptor mediated plant growth responses: Implications for photoreceptor engineering toward improved performance in crops. Front. Plant Sci. 2017, 8, 1181. [Google Scholar] [CrossRef]
- Hart, J.E.; Gardner, K.H. Lighting the way: Recent insights into the structure and regulation of phototropin blue light receptors. J. Biol. Chem. 2021, 296, 100594. [Google Scholar] [CrossRef]
- Christie, J.M.; Blackwood, L.; Petersen, J.; Sullivan, S. Plant flavoprotein photoreceptors. Plant Cell Physiol. 2015, 56, 401–413. [Google Scholar] [CrossRef]
- Boccalandro, H.E.; Giordano, C.V.; Ploschuk, E.L.; Piccoli, P.N.; Bottini, R.; Casal, J.J. Phototropins but not cryptochromes mediate the blue light-specific promotion of stomatal conductance, while both enhance photosynthesis and transpiration under full sunlight. Plant Physiol. 2012, 158, 1475–1484. [Google Scholar] [CrossRef]
- Lopez, L.; Fasano, C.; Perrella, G.; Facella, P. Cryptochromes and the circadian clock: The story of a very complex relationship in a spinning world. Genes 2021, 12, 672. [Google Scholar] [CrossRef]
- Jenkins, G.I. Structure and function of the UV-B photoreceptor UVR8. Curr. Opin. Struct. Biol. 2014, 29, 52–57. [Google Scholar] [CrossRef]
- Ahmad, A.; Ali, H.; Khan, H.; Begam, A.; Khan, S.; Ali, S.S.; Ahmad, N.; Fazal, H.; Ali, M.; Hano, C.; et al. Effect of gibberellic acid on production of biomass, polyphenolics and steviol glycosides in adventitious root cultures of Stevia rebaudiana (Bert.). Plants 2020, 9, 420. [Google Scholar] [CrossRef]
- Zhou, X.; Gong, M.; Lv, X.; Liu, Y.; Li, J.; Du, G.; Liu, L. Metabolic engineering for the synthesis of steviol glycosides: Current status and future prospects. Appl. Microbiol. Biotechnol. 2021, 105, 5367–5381. [Google Scholar] [CrossRef]
- Richman, A.; Swanson, A.; Humphrey, T.; Chapman, R.; McGarvey, B.; Pocs, R.; Brandle, J. Functional genomics uncovers three glucosyltransferases involved in the synthesis of the major sweet glucosides of Stevia rebaudiana. Plant J. 2005, 41, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Biswas, P.; Kumari, A.; Modi, A.; Kumar, N. Improvement and regulation of steviol glycoside biosynthesis in Stevia rebaudiana Bertoni. Gene 2024, 891, 147809. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Kaul, K.; Bajpai-Gupta, S.; Kaul, V.K.; Kumar, S. A comprehensive analysis of fifteen genes of steviol glycosides biosynthesis pathway in Stevia rebaudiana (Bertoni). Gene 2012, 492, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Hernández, K.V.; Moreno-Romero, J.; de la Torre, M.H.; Manríquez, C.P.; Leal, D.R.; Martínez-Garcia, J.F. Effect of light intensity on steviol glycosides production in leaves of Stevia rebaudiana plants. Phytochchemistry 2022, 194, 113027. [Google Scholar] [CrossRef] [PubMed]
- Trumpler, K.; Wu, B.; Addo, P.W.; MacPherson, S.; Lefsrud, M. Plant growth optimization using amber light supplemented with different blue light spectra. Horticulturae 2024, 10, 1097. [Google Scholar] [CrossRef]
- Kaiser, E.; Weerheim, K.; Schipper, R.; Dieleman, J.A. Partial replacement of red and blue by green light increases biomass and yield in tomato. Sci. Hortic. 2019, 249, 271–279. [Google Scholar] [CrossRef]
- Zoratti, L.; Karppinen, K.; Escobar, A.L.; Häggman, H.; Jaakola, L. Light-controlled flavonoid biosynthesis in fruits. Front. Plant Sci. 2014, 5, 534. [Google Scholar] [CrossRef]
- Ahmad, N.; Rab, A.; Ahmad, N. Light-induced biochemical variations in secondary metabolite production and antioxidant activity in callus cultures of stevia rebaudiana (Bert). J. Photochem. Photobiol. B 2016, 154, 51–56. [Google Scholar] [CrossRef]
- Yoneda, Y.; Nakashima, H.; Miyasaka, J.; Ohdoi, K.; Shimizu, H. Impact of blue, red, and far-red light treatments on gene expression and steviol glycoside accumulation in Stevia rebaudiana. Phytochchemistry 2017, 137, 57–65. [Google Scholar] [CrossRef]
- Shulgina, A.A.; Kalashnikova, E.A.; Tarakanov, I.G.; Kirakosyan, R.N.; Cherednichenko, M.Y.; Polivanova, O.B.; Baranova, E.N.; Khaliluev, M.R. Influence of light conditions and medium composition on morphophysiological characteristics of Stevia rebaudiana Bertoni in vitro and in vivo. Horticulturae 2021, 7, 195. [Google Scholar] [CrossRef]
- Semenova, N.A.; Ivanitskikh, A.S.; Uyutova, N.I.; Smirnov, A.A.; Proshkin, Y.A.; Burynin, D.A.; Kachan, S.A.; Sokolov, A.V.; Dorokhov, A.S.; Chilingaryan, N.O. Effect of UV stress on the antioxidant capacity, photosynthetic activity, flavonoid and steviol glycoside accumulation of stevia rebaudiana Bertoni. Horticulturae 2024, 10, 210. [Google Scholar] [CrossRef]
- Kang, L.K.; Huang, Y.J.; Lim, W.T.; Hsu, P.H.; Hwang, P.A. Growth, pigment content, antioxidant activity, and phytoene desaturase gene expression in Caulerpa lentillifera grown under different combinations of blue and red light-emitting diodes. J. Appl. Phycol. 2020, 32, 1971–1982. [Google Scholar] [CrossRef]
- Yılmaz, F.M.; Görgüç, A.; Uygun, Ö.; Bircan, C. Steviol glycosides and polyphenols extraction from Stevia rebaudiana Bertoni leaves using maceration, microwave and ultrasound-assisted techniques. Sep. Sci. Technol. 2020, 56, 936–948. [Google Scholar] [CrossRef]
- FAO/WHO. Steviol Glycosides from Stevia rebaudiana Bertoni. In Proceedings of the FAO Joint FAO/WHO Expert Committee on Food Additives (JECFA), Rome, Italy, 6–15 June 2017. [Google Scholar]
- Son, K.H.; Oh, M.M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Serfaty, M.; Ibdah, M.; Fischer, R.; Chaimovitsh, D.; Saranga, Y.; Dudai, N. Dynamics of yield components and stevioside production in Stevia rebaudiana grown under different planting times, plant stands and harvest regime. Ind. Crops Prod. 2013, 50, 731–736. [Google Scholar] [CrossRef]
- Ohashi-Kaneko, K.; Takase, M.; Kon, N.; Fujiwara, K.; Kurata, K. Effect of light quality on growth and vegetable quality in leaf lettuce, spinach amd komatsuna. Environ. Control Biol. 2007, 45, 189–198. [Google Scholar] [CrossRef]
- Frank, S.R. The effectiveness of the spectrum in chlorophyll formation. J. Gen. Physiol. 1946, 29, 157–179. [Google Scholar] [CrossRef]
- Zhang, J.; Han, C.; Liu, Z. Absorption spectrum estimating rice chlorophyll concentration: Perliminary investigations. J. Plant Breed. Crop Sci. 2009, 1, 223–229. [Google Scholar]
- Ceunen, S.; Geuns, J. Influene of photoperiodism on the spatio-temporal accumulation of steviol glycosides in Stevia rebaudiana Bertoni. Plant Sci. 2013, 198, 72–82. [Google Scholar] [CrossRef]
- Kalandia, A.; Papunidze, G.; Vanidze, M.; Papunidze, S. HPLC of stevia (Stevia rebaudiana Bertoni) diterpene glycosides. Bull. Georgian Acad. Sci. 2004, 169, 147–150. [Google Scholar]
- Kienle, U. Welches Stevia hätten sie den gerne? Anbau und herstellung-perspektiven weltweit. J. Verbr. Lebensm. 2010, 5, 241–250. [Google Scholar] [CrossRef]
- Madore, M.A. Synthesis and degradation of diterpene glycosides in source leaves of Stevia rebaudiana (Bert.) Bertoni. Plant Biol. 2000, 129. [Google Scholar]
- Ptak, A.; Szewczyk, A.; Simlat, M.; Pawłowska, B.; Warchoł, M. LED light improves shoot multiplication, steviol glycosides and phenolic compounds biosynthesis in Stevia rebaudiana Bertoni in vitro culture. Sci. Rep. 2024, 14, 30860. [Google Scholar] [CrossRef]
- Wang, P.; Chen, S.; Gu, M.; Chen, X.; Chen, X.; Yang, J.; Zhao, F.; Ye, N. Exploration of the effects of different blue LED light intensities on flavonoid and lipid metabolism in tea plants via transcriptomics and metabolomics. Int. J. Mol. Sci. 2020, 21, 4606. [Google Scholar] [CrossRef]
- Thoma, F.; Somborn-Schulz, A.; Schlehuber, D.; Keuter, V.; Deerberg, G. Effects of light on secondary metabolites in selected leafy greens: A review. Front. Plant Sci. 2020, 11, 497. [Google Scholar] [CrossRef]
- Hashim, M.; Ahmad, B.; Drouet, S.; Hano, C.; Abbasi, B.H.; Anjum, S. Comparative effects of different light sources on the production of key secondary metabolites in plants in vitro cultures. Plants 2021, 10, 1521. [Google Scholar] [CrossRef] [PubMed]
- Rengasamy, N.; Othman, R.Y.; Che, H.S.; Harikrishna, J.A. Beyond the PAR spectra: Impact of light quality on the germination, flowering, and metabolite content of Stevia rebaudiana (Bertoni). J. Sci. Food Agric. 2022, 102, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Condition | P1 | P2 | P3 | P4 |
|---|---|---|---|---|
| W * | 2.38 ± 0.30 b | 7.51 ± 0.18 c | 18.59 ± 0.28 c | 17.79 ± 0.32 c |
| R | 2.58 ± 0.13 c | 8.97 ± 0.18 d | 23.53 ± 0.47 d | 22.52 ± 0.43 d |
| G | 2.10 ± 0.19 a | 5.58 ± 0.20 a | 17.15 ± 0.31 a | 14.46 ± 0.37 a |
| B | 2.09 ± 0.17 a | 7.12 ± 0.28 b | 17.36 ± 0.28 b | 16.62 ± 0.41 b |
| Light Condition | P1 | P2 | P3 | P4 |
|---|---|---|---|---|
| W * | 0.24 ± 0.03 b | 0.75 ± 0.02 c | 1.86 ± 0.03 c | 1.78 ± 0.03 c |
| R | 0.26 ± 0.01 c | 0.90 ± 0.02 d | 2.35 ± 0.05 d | 2.25 ± 0.04 d |
| G | 0.21 ± 0.02 a | 0.56 ± 0.02 a | 1.72 ± 0.03 a | 1.45 ± 0.04 a |
| B | 0.21 ± 0.03 a | 0.71 ± 0.03 b | 1.74 ± 0.03 b | 1.66 ± 0.04 b |
| Light Condition | P1 | P2 | P3 | P4 |
|---|---|---|---|---|
| W * | 18.97 ± 0.78 a | 21.76 ± 1.32 b | 22.89 ± 1.53 c | 23.61 ± 2.04 d |
| R | 18.34 ± 0.43 a | 21.39 ± 1.44 b | 21.47 ± 1.36 c | 23.47 ± 0.91 d |
| G | 19.42 ± 0.98 a | 22.62 ± 1.00 b | 22.92 ± 0.30 c | 23.13 ± 1.04 c |
| B | 19.85 ± 0.50 a | 22.98 ± 0.43 b | 23.65 ± 0.69 c | 23.77 ± 0.60 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, C.-T.; Christabel, V.; Le, M.A.; Tsai, M.-L.; Wang, S.-T. Effects of Light Conditions on the Leaf Growth and Steviol Glycoside Yields of Hydroponically Cultivated Stevia Across Growth Stages. Horticulturae 2025, 11, 316. https://doi.org/10.3390/horticulturae11030316
Chou C-T, Christabel V, Le MA, Tsai M-L, Wang S-T. Effects of Light Conditions on the Leaf Growth and Steviol Glycoside Yields of Hydroponically Cultivated Stevia Across Growth Stages. Horticulturae. 2025; 11(3):316. https://doi.org/10.3390/horticulturae11030316
Chicago/Turabian StyleChou, Cheng-Tai, Vivian Christabel, Mai Anh Le, Min-Lang Tsai, and Shang-Ta Wang. 2025. "Effects of Light Conditions on the Leaf Growth and Steviol Glycoside Yields of Hydroponically Cultivated Stevia Across Growth Stages" Horticulturae 11, no. 3: 316. https://doi.org/10.3390/horticulturae11030316
APA StyleChou, C.-T., Christabel, V., Le, M. A., Tsai, M.-L., & Wang, S.-T. (2025). Effects of Light Conditions on the Leaf Growth and Steviol Glycoside Yields of Hydroponically Cultivated Stevia Across Growth Stages. Horticulturae, 11(3), 316. https://doi.org/10.3390/horticulturae11030316