
Locating Appropriate Reference Genes in Heteroblastic Plant Ottelia cordata for Quantitative Real-Time PCR Normalization


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Selection of Reference Genes and the Design of Primers
2.3. Extraction of RNA and Synthesis of cDNA
2.4. qRT-PCR Analysis
2.5. Gene Expression Stability Analysis
2.6. Statistical Analysis
3. Results
3.1. Selection of Candidate Reference Genes
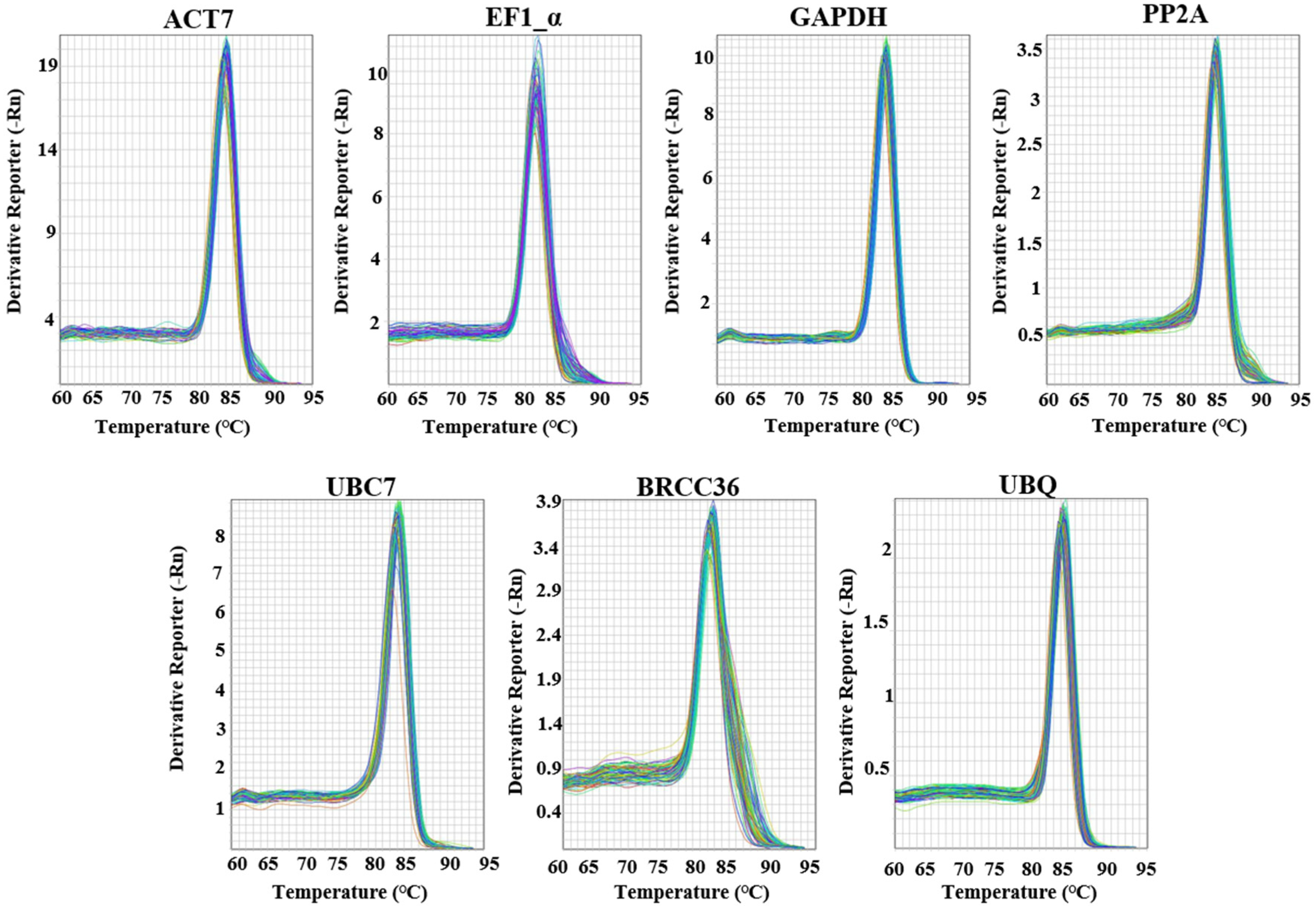
3.2. Verification of Primer Specificity and Efficacy
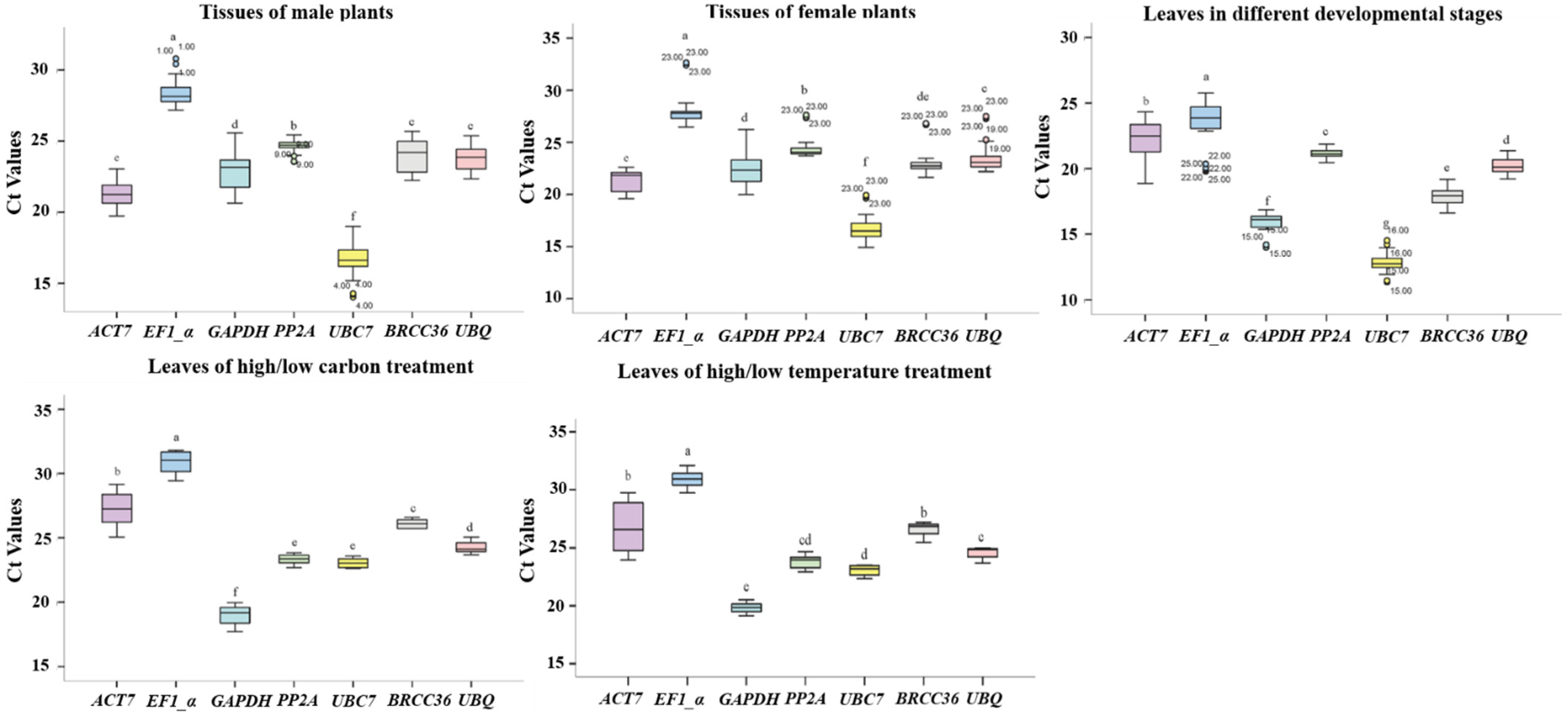
3.3. Ct Value Analyses of Seven Reference Genes
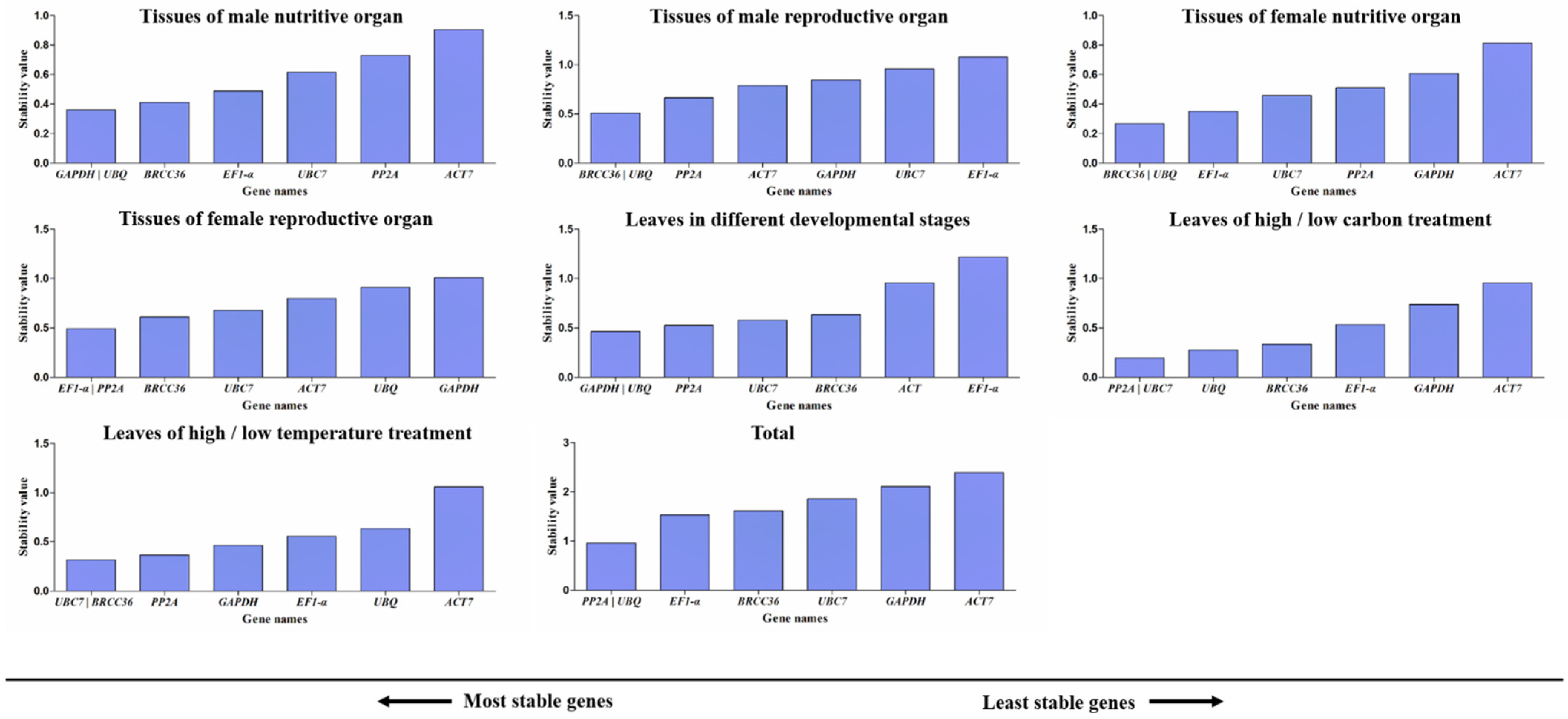
3.4. GeNorm Analysis of Candidate Reference Genes
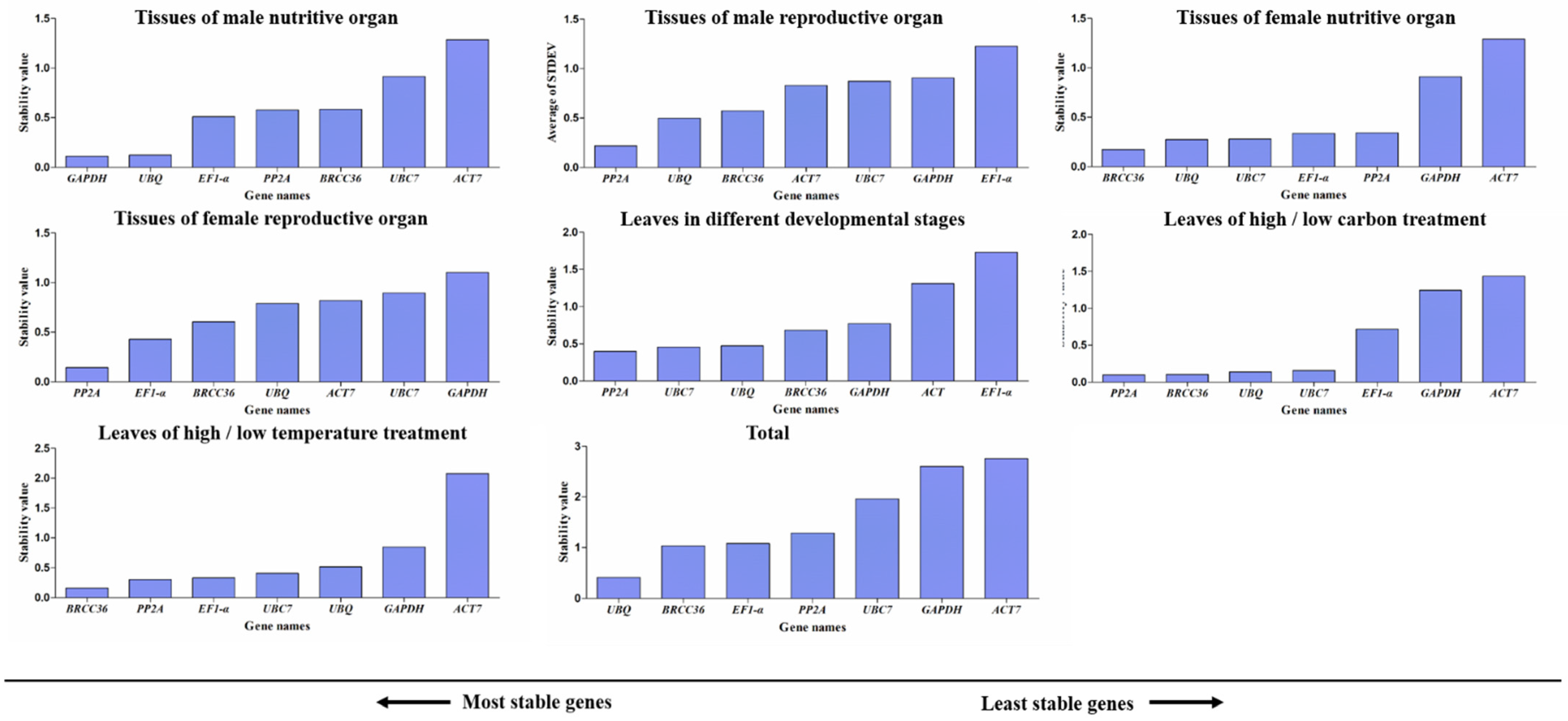
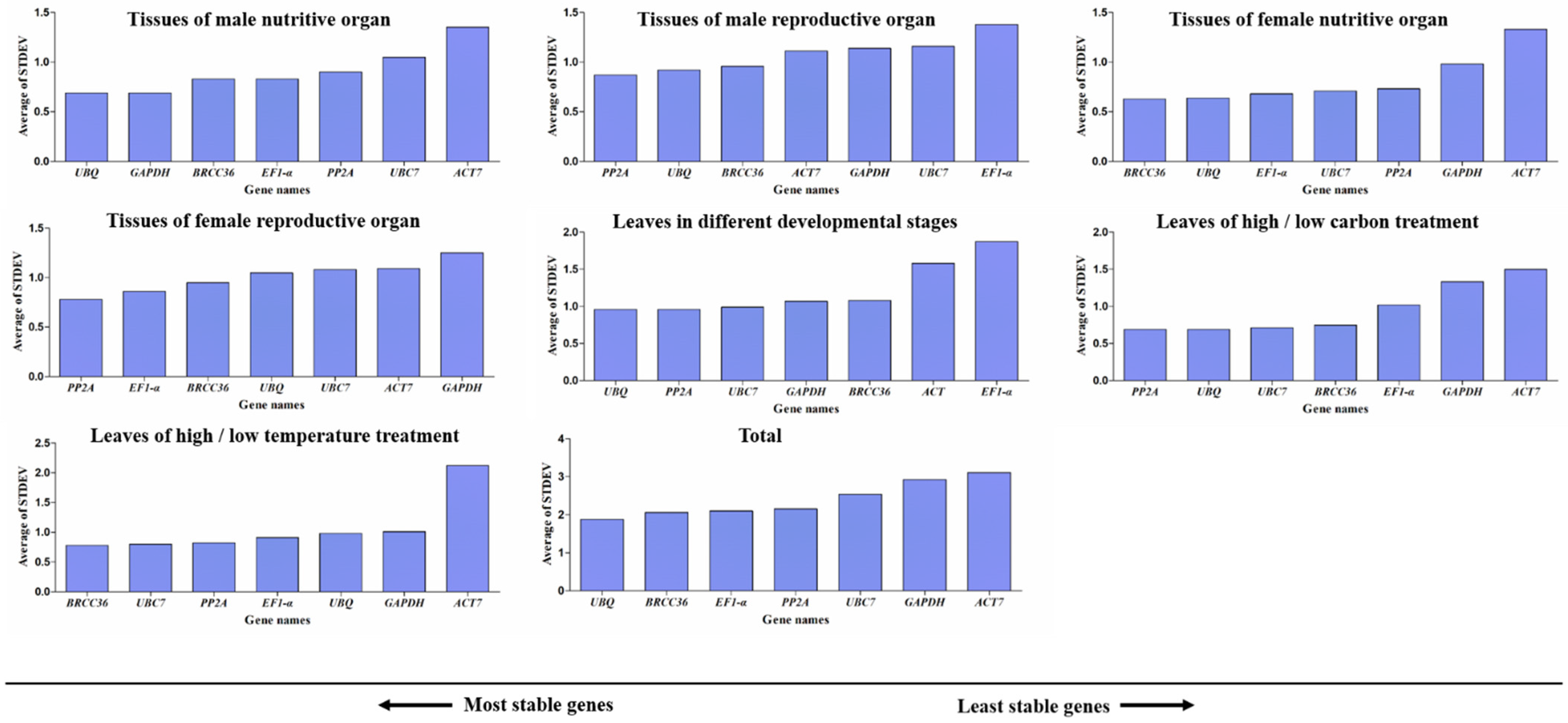
3.5. NormFinder Analysis of Candidate Reference Genes
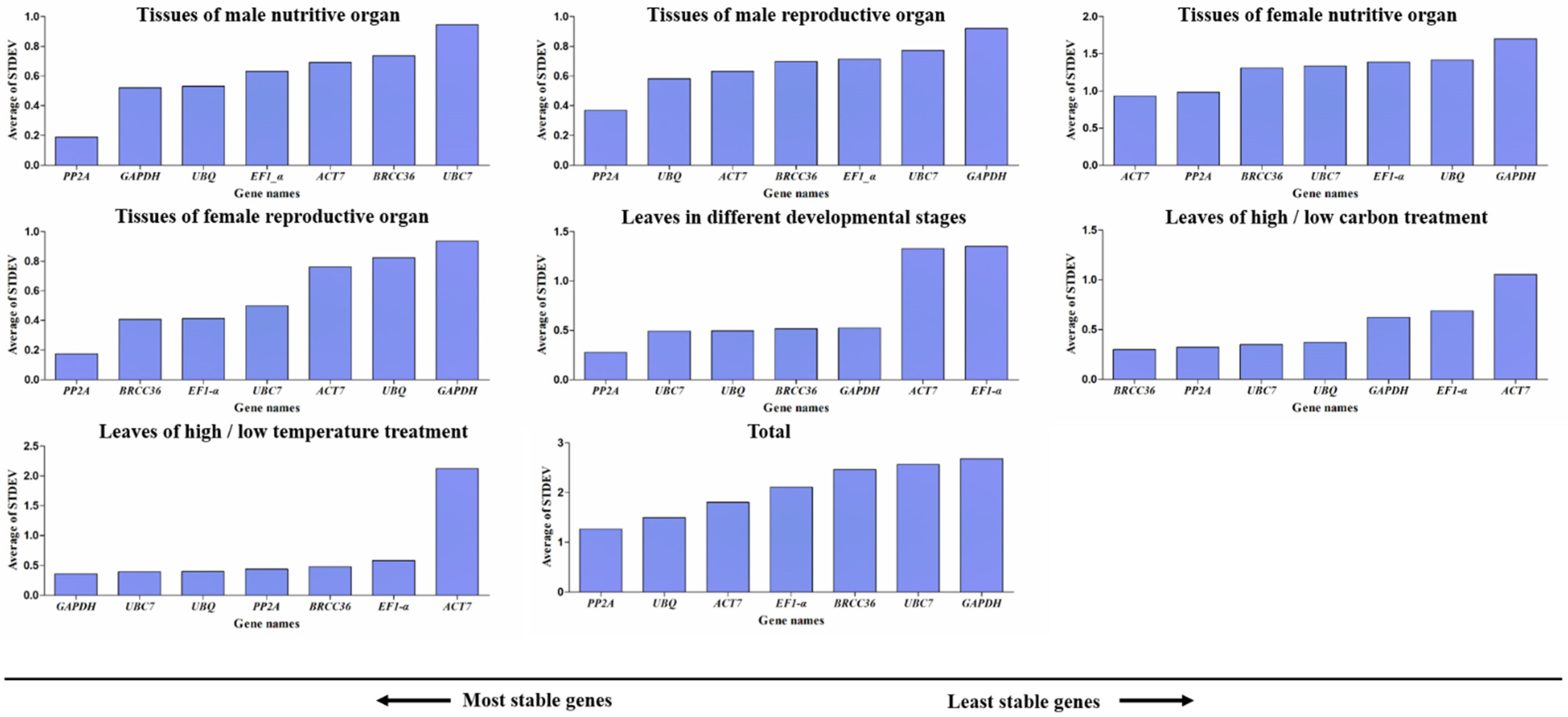
3.6. BestKeeper Analysis of Candidate Reference Genes
3.7. Delta Ct Analysis of Candidate Reference Genes
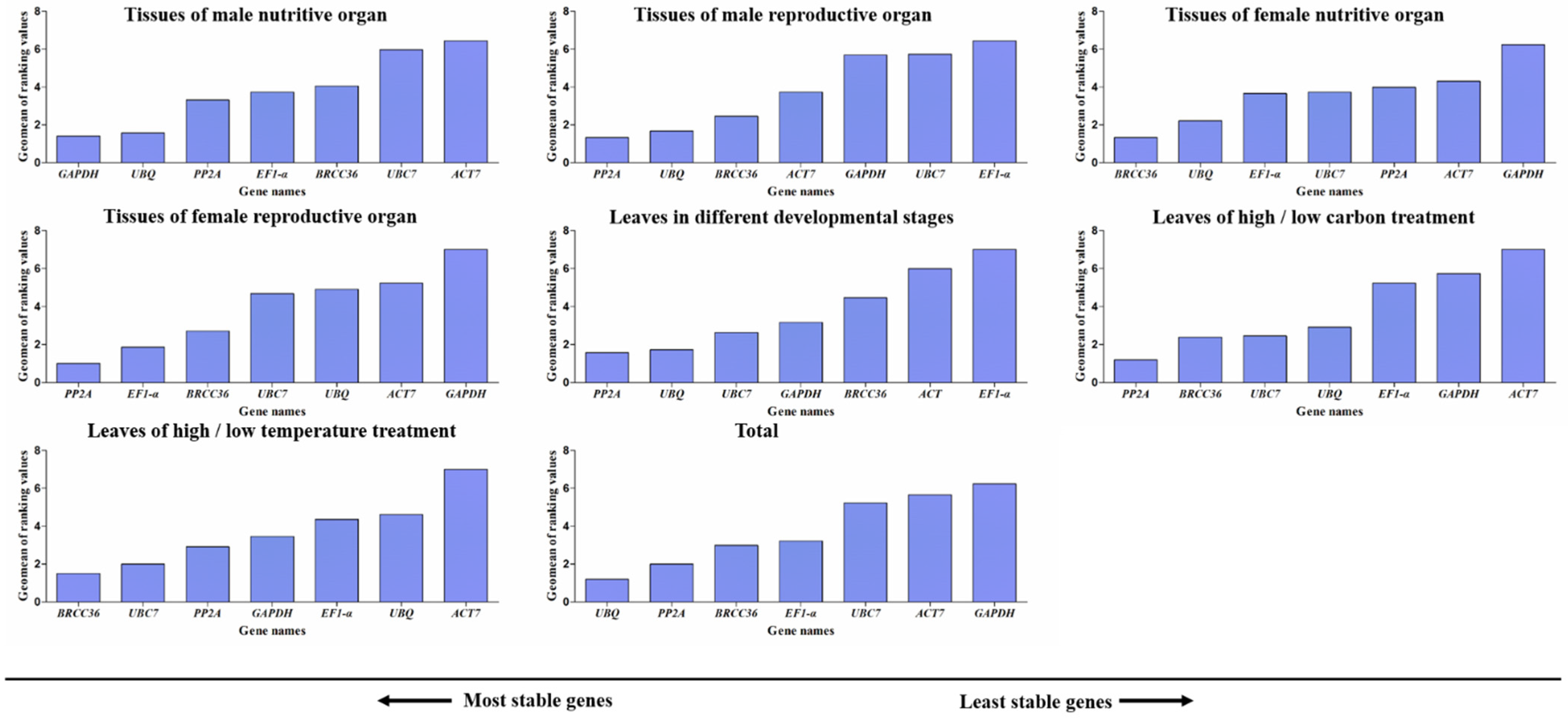
3.8. Comprehensive Analysis
3.9. Validation of Reference Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gadkar, V.Y.; Filion, M. New developments in quantitative real-time polymerase chain reaction technology. Curr. Issues Mol. Biol. 2014, 16, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Guénin, S.; Mauriat, M.; Pelloux, J.; Van Wuytswinkel, O.; Bellini, C.; Gutierrez, L. Normalization of qRT-PCR data: The necessity of adopting a systematic, experimental conditions-specific, validation of references. J. Exp. Bot. 2009, 60, 487–493. [Google Scholar] [CrossRef]
- Udvardi, M.K.; Czechowski, T.; Scheible, W.R. Eleven golden rules of quantitative RT-PCR. Plant Cell 2008, 20, 1736–1737. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Yin, L.Y.; Jiang, H.S.; Li, W.; Gontero, B.; Maberly, S.C. Biochemical and biophysical CO2 concentrating mechanisms in two species of freshwater macrophyte within the genus Ottelia (Hydrocharitaceae). Photosynth. Res. 2014, 121, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Bowes, G.; Rao, S.K.; Estavillo, G.M.; Reiskind, J.B. C4 mechanisms in aquatic angiosperms: Comparisons with terrestrial C4 systems. Funct. Plant Biol. 2002, 29, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Bowes, G. Chapter 5 single-cell C4 photosynthesis in aquatic plants. In C4 Photosynthesis and Related CO2 Concentrating Mechanisms, 2nd ed.; Raghavendra, A., Sage, R., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 32, pp. 63–80. [Google Scholar]
- Wang, S.; Li, P.; Liao, Z.; Wang, W.; Chen, T.; Yin, L.; Jiang, H.S.; Li, W. Adaptation of inorganic carbon utilization strategies in submerged and floating leaves of heteroblastic plant Ottelia cordata. Environ. Exp. Bot. 2022, 196, 104818. [Google Scholar] [CrossRef]
- Huang, W.; Han, S.; Xing, Z.; Li, W. Responses of leaf anatomy and CO2 concentrating mechanisms of the aquatic plant Ottelia cordata to variable CO2. Front. Plant Sci. 2020, 11, 1261. [Google Scholar] [CrossRef] [PubMed]
- Iversen, L.L.; Winkel, A.; Baastrup-Spohr, L.; Hinke, A.B.; Alahuhta, J.; Baattrup-Pedersen, A.; Birk, S.; Brodersen, P.; Chambers, P.A.; Ecke, F.; et al. Catchment properties and the photosynthetic trait composition of freshwater plant communities. Science 2019, 366, 878–881. [Google Scholar] [CrossRef]
- Maberly, S.C.; Gontero, B. Ecological imperatives for aquatic CO2-concentrating mechanisms. J. Exp. Bot. 2017, 68, 3797–3814. [Google Scholar] [CrossRef]
- Pedersen, O.; Colmer, T.D.; Sand-Jensen, K. Underwater photosynthesis of submerged plants—Recent advances and methods. Front. Plant Sci. 2013, 4, 140. [Google Scholar] [CrossRef]
- Yuan, G.; Li, Z.; Deng, J.; Pacheco, J.P.; Fu, H. Editorial: The performance and adaptation of aquatic plants under global changes. Front. Plant Sci. 2024, 15, 1380921. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, G. Alternative pathways of carbon dioxide fixation: Insights into the early evolution of life? Annu. Rev. Microbiol. 2011, 65, 631–658. [Google Scholar] [CrossRef] [PubMed]
- Von Caemmerer, S.; Ghannoum, O.; Furbank, R.T. C4 photosynthesis: 50 years of discovery and innovation. J. Exp. Bot. 2017, 68, 97–102. [Google Scholar] [CrossRef]
- Hang, H.; Wu, Y. Quantification of photosynthetic inorganic carbon utilisation via a bidirectional stable carbon isotope tracer. Acta Geochim. 2016, 35, 130–137. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Wang, H.; Shu, L.; Lv, C.; Liu, N.; Long, Y.; Peng, X.; Ling, H.; Tao, T.; Tang, J.; Cheng, Y.; et al. BRCC36 deubiquitinates HMGCR to regulate the interplay between ferroptosis and pyroptosis. Adv. Sci. 2024, 11, e2304263. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef]
- Sundaram, V.; Sampathkumar, N.; Massaad, C.; Grenier, J. Optimal use of statistical methods to validate reference gene stability in longitudinal studies. PLoS ONE 2019, 14, e0219440. [Google Scholar] [CrossRef]
- Chan, O.; Keng, B.; Ling, M. Correlation and variation-based method for identifying reference genes from large datasets. Electron. Physician 2014, 6, 719–727. [Google Scholar]
- Curis, E.; Nepost, C.; Laroche, G.; Courtin, C.; Laplanche, J.; Étain, B.; Marie-Claire, C. Selecting reference genes in RT-qPCR based on equivalence tests: A network based approach. Sci. Rep. 2019, 9, 16231. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Wang, J.; Zhang, B. RefFinder: A web-based tool for comprehensively analyzing and identifying reference genes. Funct. Integr. Genom. 2023, 15, 23, 125. [Google Scholar] [CrossRef]
- Ling, D.; Salvaterra, P. Robust RT-qPCR data normalization: Validation and selection of internal reference genes during post-experimental data analysis. PLoS ONE 2011, 6, e17762. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, K.D.; Fernandes de Melo, D.; Morais, V.D.; Vasconcelos, I.M.; Costa, J.H. Selection of suitable soybean EF1_α genes as internal controls for real-time PCR analyses of tissues during plant development and under stress conditions. Plant Cell Rep. 2014, 33, 1453–1465. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.; Lu, H.; Tian, C.; Xu, T.; Song, C.; Wang, W.; Wei, P.; Gu, F.; Liu, D.; Cai, Y.; et al. Selection of suitable reference genes for gene expression normalization studies in Dendrobium huoshanense. Genes 2022, 13, 1486. [Google Scholar] [CrossRef]
- Wang, B.; Duan, H.; Chong, P.; Su, S.; Shan, L.; Yi, D.; Wang, L.; Li, Y. Systematic selection and validation of suitable reference genes for quantitative real-time PCR normalization studies of gene expression in Nitraria tangutorum. Sci. Rep. 2020, 10, 15891. [Google Scholar] [CrossRef]
- Scarabel, L.; Milani, A.; Panozzo, S.; Rasori, A. Suitable reference genes for accurate gene expression analysis in Papaver rhoeas under 2,4-D herbicide stress. Pestic. Biochem. Physiol. 2017, 143, 66–72. [Google Scholar] [CrossRef]
- Zeqiraj, E.; Tian, L.; Piggott, C.A.; Pillon, M.C.; Duffy, N.M.; Ceccarelli, D.F.; Keszei, A.F.; Lorenzen, K.; Kurinov, I.; Orlicky, S.; et al. Higher-order assembly of BRCC36-KIAA0157 is required for DUB activity and biological function. Mol. Cell 2015, 59, 970–983. [Google Scholar] [CrossRef]
- Rabl, J.; Bunker, R.D.; Schenk, A.D.; Cavadini, S.; Gill, M.E.; Abdulrahman, W.; Andrés-Pons, A.; Luijsterburg, M.S.; Ibrahim, A.F.M.; Branigan, E.; et al. Structural basis of BRCC36 function in DNA repair and immune regulation. Mol. Cell 2019, 75, 483–497. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Gene Description | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Length (bp) | Efficiency (%) | R2 |
|---|---|---|---|---|---|---|
| ACT7 | actin 7 | CCCTTTGGAGCATTCG | GCCCTCGGAGCATCAT | 136 | 103.3 | 0.997 |
| EF1_α | elongation factor 1-alpha | GAAGCACTGCCAAAGG | GGAAGCAACGGAAGAT | 137 | 94.2 | 0.990 |
| BRCC36 | lys-63-specific deubiquitinase BRCC36 | CCACCGAGACCGAAGA | CCAGATTAGAGCGACAGG | 83 | 96.7 | 0.993 |
| PP2A | serine/threonine protein phosphatase 2A | GCAGTCCAGAGCCTAACACT | CCAGCCGCTTCCAAAT | 148 | 99.9 | 0.999 |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase | AGGTCACCGTCTTTGGA | AACGAACATGGGAGCAT | 179 | 103.2 | 0.999 |
| UBC7 | ubiquitin-conjugating enzyme E2 7 | AGATAGGCGGGATGAGTT | TGACCTGCTTTACATTAGACA | 176 | 90.1 | 0.999 |
| UBQ | E3 ubiquitin-protein ligase RNF167 | GCGTCTTCGCATTCAG | TTCACAAGCCAACAGC | 270 | 101.3 | 0.998 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, P.; Li, R.; Huang, J.; Yin, L. Locating Appropriate Reference Genes in Heteroblastic Plant Ottelia cordata for Quantitative Real-Time PCR Normalization. Horticulturae 2025, 11, 313. https://doi.org/10.3390/horticulturae11030313
Guo P, Li R, Huang J, Yin L. Locating Appropriate Reference Genes in Heteroblastic Plant Ottelia cordata for Quantitative Real-Time PCR Normalization. Horticulturae. 2025; 11(3):313. https://doi.org/10.3390/horticulturae11030313
Chicago/Turabian StyleGuo, Panyang, Runan Li, Jiaquan Huang, and Liyan Yin. 2025. "Locating Appropriate Reference Genes in Heteroblastic Plant Ottelia cordata for Quantitative Real-Time PCR Normalization" Horticulturae 11, no. 3: 313. https://doi.org/10.3390/horticulturae11030313
APA StyleGuo, P., Li, R., Huang, J., & Yin, L. (2025). Locating Appropriate Reference Genes in Heteroblastic Plant Ottelia cordata for Quantitative Real-Time PCR Normalization. Horticulturae, 11(3), 313. https://doi.org/10.3390/horticulturae11030313