
The Volatile Composition, Biosynthesis Pathways, Breeding Strategies, and Regulation Measures of Apple Aroma: A Review

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Volatile Composition in Apple Fruit
3. Biosynthesis Pathways of Apple Volatiles
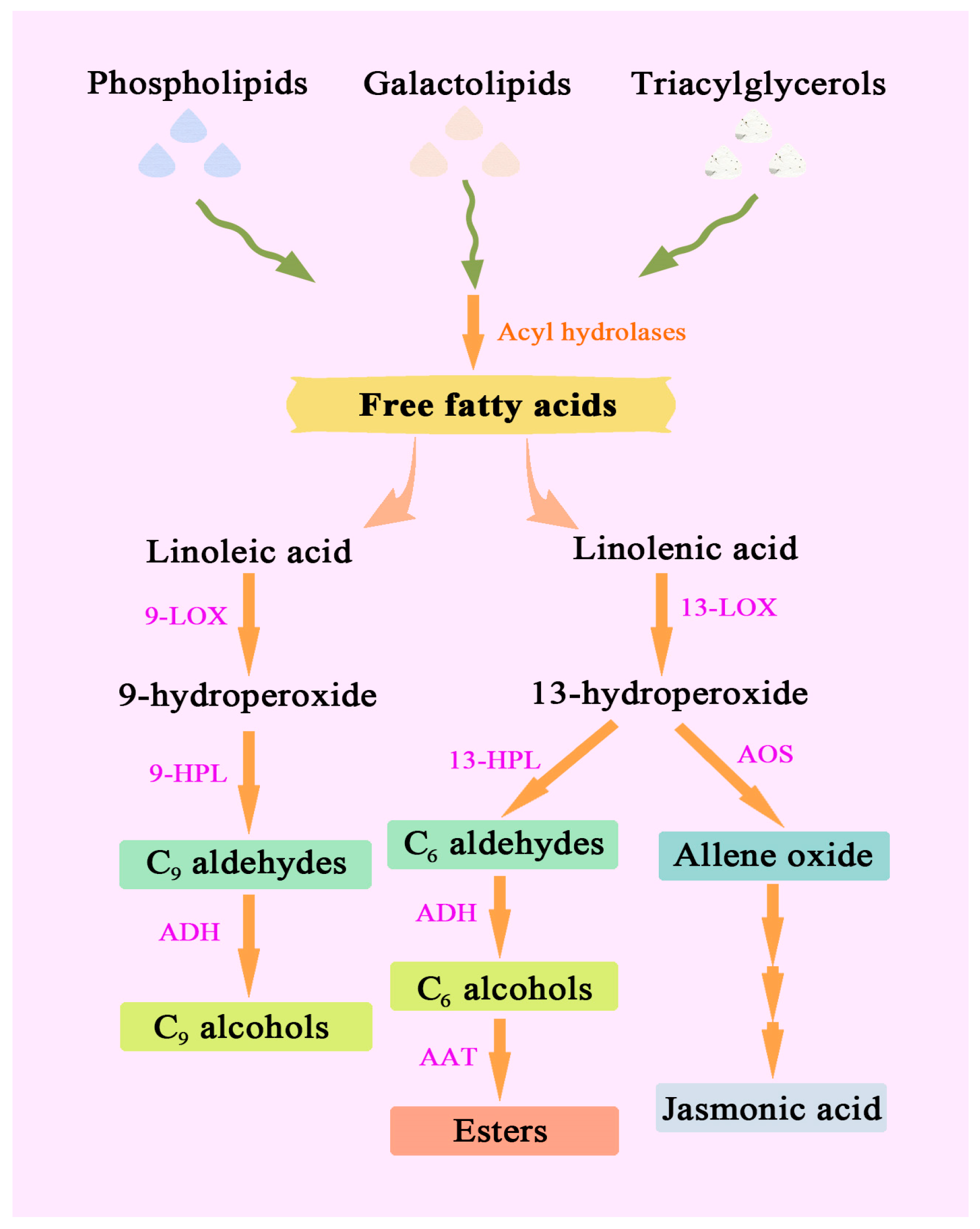
3.1. Fatty Acid-Derived Compounds
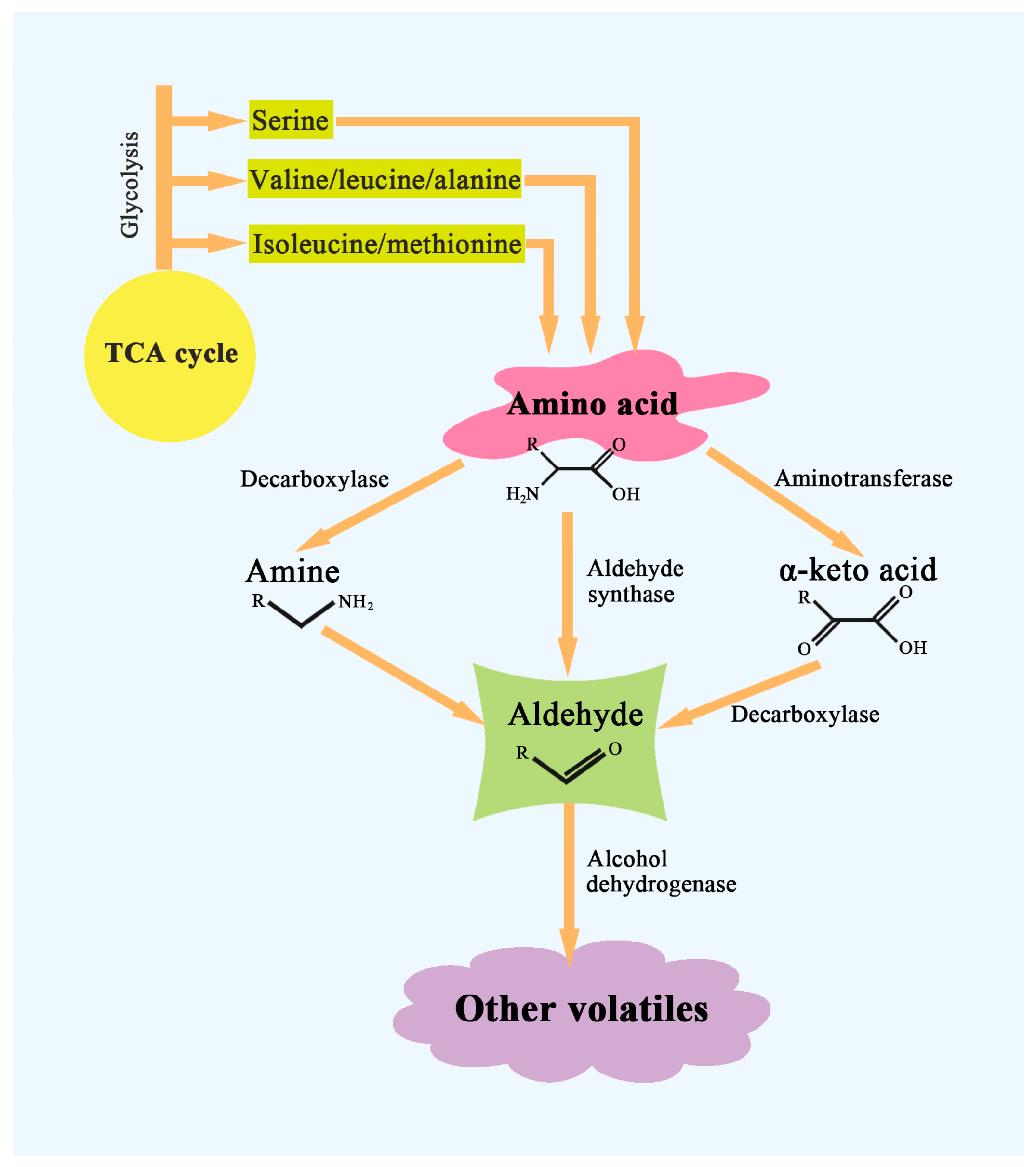
3.2. Amino Acid-Derived Compounds
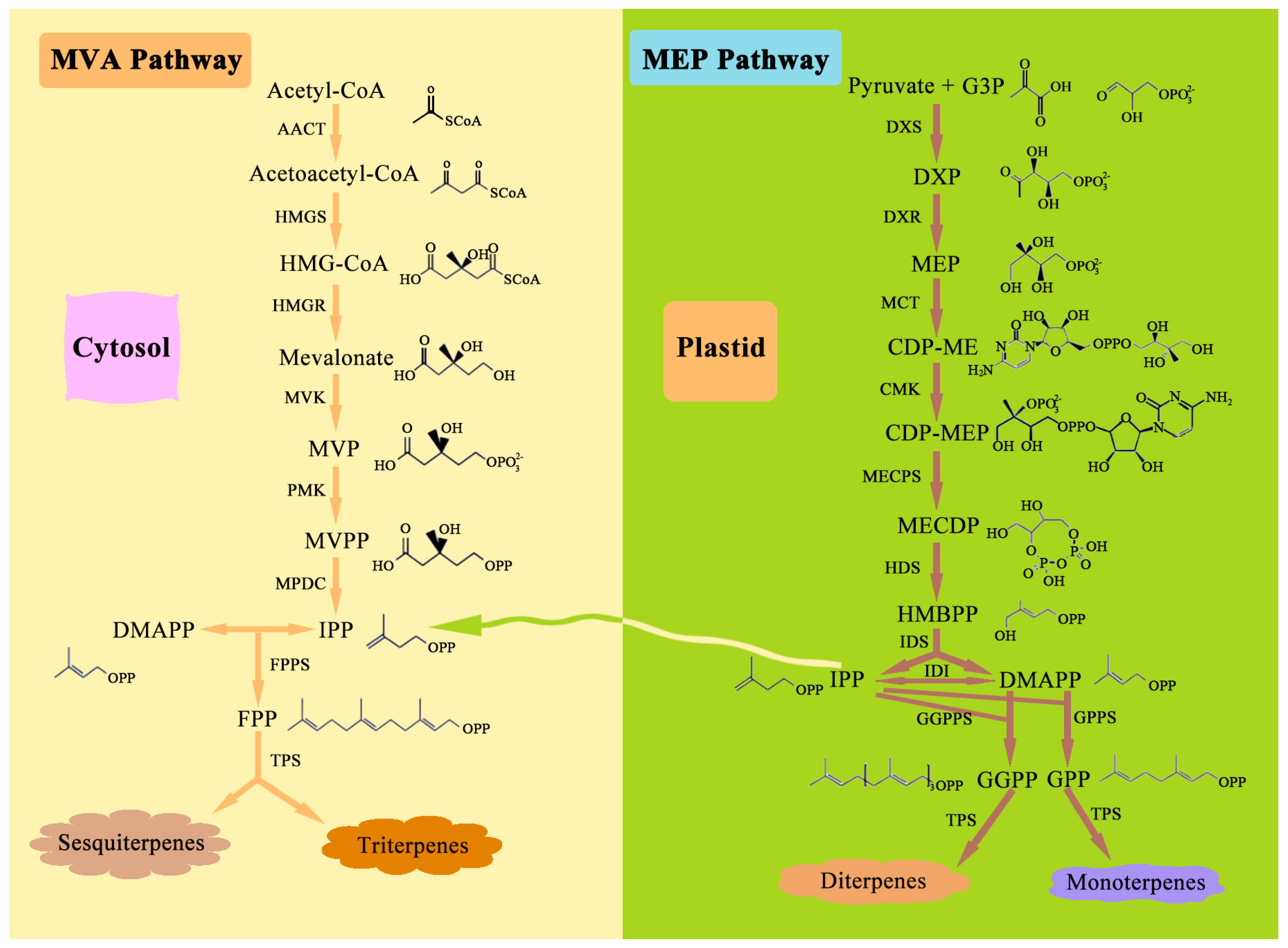
3.3. Terpenoid Pathway
3.4. Phenylpropanoid Pathway
4. Breeding Strategies on Apple Aroma
5. Regulation Measures for Apple Aroma
5.1. Environmental Condition
5.2. Agricultural Practice
5.3. Postharvest Management
6. Issues and Prospects
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Harker, F.R.; Kupferman, E.M.; Marin, A.B.; Gunson, F.A.; Triggs, C.M. Eating quality standards for apples based on consumer preferences. Postharvest Biol. Technol. 2008, 50, 70–78. [Google Scholar] [CrossRef]
- Klee, H.J. Improving the flavor of fresh fruits: Genomics, biochemistry, and biotechnology. New Phytol. 2010, 187, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Abbott, J.A.; Saftner, R.A.; Gross, K.C.; Vinyard, B.T.; Janick, J. Consumer evaluation and quality measurement of fresh-cut slices of ‘Fuji’, ‘Golden Delicious’, ‘GoldRush’, and ‘Granny Smith’ apples. Postharvest Biol. Technol. 2004, 33, 127–140. [Google Scholar] [CrossRef]
- Zhu, D.; Ren, X.; Wei, L.; Cao, X.; Ge, Y.; Liu, H.; Li, J. Collaborative analysis on difference of apple fruits flavour using electronic nose and electronic tongue. Sci. Hortic. 2020, 260, 108879. [Google Scholar] [CrossRef]
- Yan, D.; Shi, J.; Ren, X.; Tao, Y.; Ma, F.; Li, R.; Liu, X.; Liu, C. Insights into the aroma profiles and characteristic aroma of ‘Honeycrisp’ apple (Malus × domestica). Food Chem. 2020, 327, 127074. [Google Scholar] [CrossRef]
- Song, J.; Forney, C.F. Flavour volatile production and regulation in fruit. Can. J. Plant Sci. 2008, 88, 537–550. [Google Scholar] [CrossRef]
- Dixon, J.; Hewett, E.W. Factors affecting apple aroma/flavour volatile concentration: A review. N. Z. J. Crop Hortic. Sci. 2000, 28, 155–173. [Google Scholar] [CrossRef]
- Yang, S.; Meng, Z.; Fan, J.; Yan, L.; Yang, Y.; Zhao, Z. Evaluation of the volatile profiles in pulp of 85 apple cultivars (Malus domestica) by HS-SPME combined with GC-MS. J. Food Meas. Charact. 2021, 15, 4215–4225. [Google Scholar] [CrossRef]
- Young, J.C.; Chu, C.; Lu, X.; Zhu, H. Ester variability in apple varieties as determined by solid-phase microextraction and gas chromatography-mass spectrometry. J. Agric. Food Chem. 2004, 52, 8086–8093. [Google Scholar] [CrossRef]
- Aprea, E.; Corollaro, M.L.; Betta, E.; Endrizzi, I.; Dematte, M.L.; Biasioli, F.; Gasperi, F. Sensory and instrumental profiling of 18 apple cultivars to investigate the relation between perceived quality and odour and flavour. Food Res. Int. 2012, 49, 677–686. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, M.; Wu, M.; Li, X.; Liu, H.; Niu, N.; Li, S.; Chen, L. Volatile organic compounds (VOCs) from plants: From release to detection. TrAC-Trends Anal. Chem. 2023, 158, 116872. [Google Scholar] [CrossRef]
- Haick, H. Advances in volatile organic compounds detection: From fundamental research to real-world applications. Appl. Phys. Rev. 2024, 11, 040401. [Google Scholar] [CrossRef]
- Bizzio, N.; Tieman, M.; Munoz, P. Branched-chain volatiles in fruit: A molecular perspective. Front. Plant Sci. 2022, 12, 814138. [Google Scholar] [CrossRef] [PubMed]
- Shoffe, A.; Rudell, D.; Park, D.; Algul, E.; Qin, M.; Shi, M.; Dando, R.; Watkins, B. Aroma volatiles and sensory quality of organically-grown apple cultivars after dynamic controlled atmosphere (DCA) storage and comparison with CA-stored fruit with and without 1-methylcyclopropene (1-MCP). Postharvest Biol. Technol. 2024, 218, 113162. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Rowan, D.D.; Allen, J.M.; Fielder, S.; Hunt, M.B. Biosynthesis of straight-chain ester volatiles in Red Delicious and Granny Smith apples using deuterium-labeled precursors. J. Agric. Food Chem. 1999, 47, 2553–2562. [Google Scholar] [CrossRef]
- Schwab, W.; Davidovich-Rikanati, R.; Lewinsohn, E. Biosynthesis of plant-derived flavor compounds. Plant J. 2008, 54, 712–732. [Google Scholar] [CrossRef]
- Dunemann, F.; Ulrich, D.; Malysheva-Otto, L.; Weber, W.; Longhi, S.; Velasco, R.; Costa, F. Functional allelic diversity of the apple alcohol acyl-transferase gene MdAAT1 associated with fruit ester volatile contents in apple cultivars. Mol. Breed. 2012, 29, 609–625. [Google Scholar] [CrossRef]
- McGarvey, D.J.; Croteau, R. Terpenoid metabolism. Plant Cell 1995, 7, 1015–1026. [Google Scholar]
- Zhu, Y.; Rudell, D.R.; Mattheis, J.P. Characterization of cultivar differences in alcohol acyltransferase and 1-aminocyclopropane-1-carboxylate synthase gene expression and volatile ester emission during apple fruit maturation and ripening. Postharvest Biol. Technol. 2008, 49, 330–339. [Google Scholar] [CrossRef]
- Liu, X.; Hao, N.; Feng, R.; Meng, Z.; Li, Y.; Zhao, Z. Transcriptome and metabolite profiling analyses provide insight into volatile compounds of the apple cultivar ‘Ruixue’ and its parents during fruit development. BMC Plant Biol. 2021, 21, 231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Shen, J.; Wei, W.; Xi, W.; Xu, C.; Ferguson, I.; Chen, K. Expression of genes associated with aroma formation derived from the fatty acid pathway during peach fruit ripening. J. Agric. Food Chem. 2010, 58, 6157–6165. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, D.; Li, S.; Yang, H.; Zhao, Z. GC-MS metabolite and transcriptome analyses reveal the differences of volatile synthesis and gene expression profiling between two apple varieties. Int. J. Mol. Sci. 2022, 23, 2939. [Google Scholar] [CrossRef]
- Klee, H.J.; Tieman, D.M. The genetics of fruit flavour preferences. Nat. Rev. Genet. 2018, 19, 347–356. [Google Scholar] [CrossRef]
- Schaffer, R.J.; Friel, E.N.; Souleyre, E.J.; Bolitho, K.; Thodey, K.; Ledger, S.; Bowen, J.H.; Ma, J.; Nain, B.; Cohen, D.; et al. A genomics approach reveals that aroma production in apple is controlled by ethylene predominantly at the final step in each biosynthetic pathway. Plant Physiol. 2007, 144, 1899–1912. [Google Scholar] [CrossRef]
- Mattheis, J.P.; Fan, X.; Argenta, L.C. Interactive responses of gala apple fruit volatile production to controlled atmosphere storage and chemical inhibition of ethylene action. J. Agric. Food Chem. 2005, 53, 4510–4516. [Google Scholar] [CrossRef]
- Echeverria, G.; Fuentes, T.; Graell, J.; Lara, I.; Lopez, M. Aroma volatile compounds of ‘Fuji’ apples in relation to harvest date and cold storage technology: A comparison of two seasons. Postharvest Biol. Technol. 2004, 32, 29–44. [Google Scholar] [CrossRef]
- Villatoro, C.; Altisent, R.; Echeverria, G.; Graell, J.; Lopez, M.; Lara, I. Changes in biosynthesis of aroma volatile compounds during on-tree maturation of ‘Pink Lady’ apples. Postharvest Biol. Technol. 2008, 47, 286–295. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, N.; Ma, Z.; Che, F.; Mao, J.; Chen, B. The changes in color, soluble sugars, organic acids, anthocyanins and aroma components in ‘starkrimson’ during the ripening period in China. Molecules 2016, 21, 812–825. [Google Scholar] [CrossRef]
- Shimizu, T.; Hattori, Y.; Ogami, S.; Imayoshi, Y.; Moriya, S.; Okada, K.; Sawamura, Y.; Abe, K. Identification of aroma compounds responsible for apple flavor via palatability score prediction in tree-based regression models. LWT 2024, 192, 115737. [Google Scholar] [CrossRef]
- Matich, A.J.; Rowan, D.D. Pathway analysis of branched-chain ester biosynthesis in apple using deuterium labeling and enantioselective gas chromatography-mass spectrometry. J. Agric. Food Chem. 2007, 55, 2727–2735. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef]
- Espino-Diaz, M.; Sepulveda, D.; Gonzalez-Aguilar, G.; Olivas, G. Biochemistry of apple aroma: A review. Food Technol. Biotechnol. 2016, 54, 375–394. [Google Scholar] [CrossRef]
- Park, S.; Sugimoto, N.; Larson, M.D.; Beaudry, R.; van Nocker, S. Identification of genes with potential roles in apple fruit development and biochemistry through large-scale statistical analysis of expressed sequence tags. Plant Physiol. 2006, 141, 811–824. [Google Scholar] [CrossRef]
- Goepfert, S.; Poirier, Y. β-Oxidation in fatty acid degradation and beyond. Curr. Opin. Plant Biol. 2007, 10, 245–251. [Google Scholar] [CrossRef]
- Baker, A.; Graham, I.A.; Holdsworth, M.; Smith, S.M.; Theodoulou, F.L. Chewing the fat: β-oxidation in signalling and development. Trends Plant Sci. 2006, 11, 124–132. [Google Scholar] [CrossRef]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Floral scents and fruit aromas: Functions, compositions, biosynthesis, and regulation. Front. Plant Sci. 2022, 13, 860157. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef]
- Ramya, M.; An, H.R.; Baek, Y.S.; Reddy, K.E.; Park, P.H. Orchid floral volatiles: Biosynthesis genes and transcriptional regulations. Sci. Hortic. 2018, 235, 62–69. [Google Scholar] [CrossRef]
- Bartley, I.M.; Stoker, P.G.; Martin, A.D.E.; Hatfield, S.G.S.; Knee, M. Synthesis of aroma compounds by apples supplied with alcohols and methyl esters of fatty acids. J. Sci. Food Agric. 1985, 36, 567–574. [Google Scholar] [CrossRef]
- Jerkovic, I.; Kus, P.M.; Carbonell-Barrachina, A. Volatile organic compounds as artefacts derived from natural phytochemicals sourced form plants and honey. Phytochem. Rev. 2019, 18, 871–891. [Google Scholar] [CrossRef]
- Dickinson, J.R.; Harrison, S.J.; Dickinson, J.A.; Hewlins, M.J.E. An investigation of the metabolism of isoleucine to active amyl alcohol in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 10937–10942. [Google Scholar] [CrossRef]
- Tavaria, F.K.; Dahl, S.; Carballo, F.J.; Malcata, F.X. Amino acid catabolism and generation of volatiles by lactic acid bacteria. J. Dairy Sci. 2002, 85, 2462–2470. [Google Scholar] [CrossRef]
- Gonda, I.; Bar, E.; Portnoy, V.; Lev, S.; Burger, J.; Schaffer, A.A.; Tadmor, Y.; Gepstein, S.; Giovannoni, J.J.; Katzir, N.; et al. Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L. fruit. J. Exp. Bot. 2010, 61, 1111–1123. [Google Scholar] [CrossRef]
- Beekwilder, J.; Alvarez-Huerta, M.; Neef, E.; Verstappen, F.W.A.; Bouwmeester, H.J.; Aharoni, A. Substrate usage by recombinant alcohol acyltransferases from various fruit species. Plant Physiol. 2004, 135, 1865–1878. [Google Scholar] [CrossRef]
- Wang, Q.; Quan, S.; Xiao, H. Towards efficient terpenoid biosynthesis: Manipulating IPP and DMAPP supply. Bioresour. Bioprocess. 2019, 6, 6. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E. Biochemical and molecular genetic aspects of floral scents. Plant Physiol. 2000, 122, 627–633. [Google Scholar] [CrossRef]
- Yang, S.; Hao, N.; Meng, Z.; Li, Y.; Zhao, Z. Identification, comparison and classification of volatile compounds in peels of 40 apple cultivars by HS-SPME with GC-MS. Foods 2021, 10, 1051. [Google Scholar] [CrossRef]
- Zhao, P.; Yang, Y.; Wang, X.; Guo, Y. Evolution of typical aromas and phenolic compounds of a red-fleshed apple throughout different fruit developmental periods in Xinjiang, China. Food Res. Int. 2021, 148, 110635. [Google Scholar] [CrossRef]
- Ma, N.; Zhu, J.; Wang, H.; Qian, M.; Xiao, Z. Comparative investigation of aroma-active volatiles in (“Ruixue”, “Liangzhi”, “Crystal Fuji”, and “Guifei”) apples by application of gas chromatography−mass spectrometry−olfactometry (GC−MS−O) and two-dimensional gas chromatography-quadrupole mass spectrometry (GC × GC-qMS) coupled with sensory molecular science. J. Agric. Food Chem. 2024, 72, 25229–25250. [Google Scholar]
- Ferrer, J.; Austin, C.S.; Noel, J.P. Structure and function of enzymes involved in the biosynthesis of phenylpropanoids. Plant Physiol. Biochem. 2008, 46, 356–370. [Google Scholar] [CrossRef]
- Neelam; Khatkar, A.; Sharma, K.K. Phenylpropanoids and its derivatives: Biological activities and its role in food, pharmaceutical and cosmetic industries. Crit. Rev. Food Sci. 2020, 60, 2655–2675. [Google Scholar] [CrossRef]
- Yauk, Y.K.; Chagne, D.; Tomes, S.; Matich, A.J.; Wang, M.Y.; Chen, X.Y.; Maddumage, R.; Hunt, M.B.; Rowan, D.D.; Atkinson, R.G. The O-methyltransferase gene MdoOMT1 is required for biosynthesis of methylated phenylpropenes in ripe apple fruit. Plant J. 2015, 82, 937–950. [Google Scholar] [CrossRef]
- Yang, S.; Meng, Z.; Li, Y.; Chen, R.; Yang, Y.; Zhao, Z. Evaluation of physiological characteristics, soluble sugars, organic acids and volatile compounds in ‘Orin’ apples (Malus domestica) at different ripening stages. Molecules 2021, 26, 807. [Google Scholar] [CrossRef]
- Zini, E.; Biasioli, F.; Gasperi, F.; Mott, D.; Mark, T.D.; Patocchi, A.; Gessler, C.; Komjanc, M. QTL mapping of volatile compounds in ripe apples detected by proton transfer reaction-mass spectrometry. Euphytica 2005, 145, 269–279. [Google Scholar] [CrossRef]
- Dunemann, F.; Boudichevskaia, A.; Grafe, C.; Weber, W.; Ulrich, D. QTL and candidate gene mapping for aroma compounds in the apple progeny discovery × prima. Acta Hortic. 2009, 839, 433–440. [Google Scholar] [CrossRef]
- Costa, F.; Cappellin, L.; Zini, E.; Patocchi, A.; Kellerhals, M.; Komjanc, M.; Gessler, C.; Biasioli, F. QTL validation and stability for volatile organic compounds (VOCs) in apple. Plant Sci. 2013, 211, 1–7. [Google Scholar] [CrossRef]
- Souleyre, E.; Chagne, D.; Chen, X.; Tomes, S.; Turner, R.; Wang, M.; Maddumage, R.; Hunt, M.; Winz, R.; Wiedow, C.; et al. The AAT1 locus is critical for the biosynthesis of esters contributing to ‘ripe apple’ flavour in ‘Royal gala’ and ‘Granny smith’ apples. Plant J. 2014, 78, 903–915. [Google Scholar] [CrossRef]
- Daccord, N.; Celton, J.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E.; Geest, H.; Bianco, L.; Micheletti, D.; Velasco, R.; et al. High-quality De Novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 2017, 49, 1099–1106. [Google Scholar] [CrossRef]
- Yang, S.; Yu, J.; Yang, H.; Zhao, Z. Genetic analysis and QTL mapping of aroma volatile compounds in the apple progeny ‘Fuji’ × ‘Cripps Pink’. Front. Plant Sci. 2023, 14, 1048846. [Google Scholar] [CrossRef]
- Schiller, D.; Contreras, C.; Vogt, J.; Dunemann, F.; Defilippi, B.G.; Beaudry, R.; Schwab, W. A dual positional specific lipoxygenase functions in the generation of flavor compounds during climacteric ripening of apple. Hortic. Res. 2015, 2, 15003. [Google Scholar] [CrossRef] [PubMed]
- Vogt, J.; Schiller, D.; Ulrich, D.; Schwab, W.; Dunemann, F. Identification of lipoxygenase (LOX) genes putatively involved in fruit flavour formation in apple (Malus × domestica). Tree Genet. Genomes. 2013, 9, 1493–1511. [Google Scholar] [CrossRef]
- Souleyre, E.; Greenwood, D.; Friel, E.; Karunairetnam, S.; Newcomb, R. An alcohol acyl transferase from apple (cv. Royal Gala), MpAAT1, produces esters involved in apple fruit flavor. FEBS J. 2005, 272, 3132–3144. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xu, Y.; Xu, G.; Gu, L.; Li, D.; Shu, H. Molecular cloning and expression of a gene encoding alcohol acyltransferase (MdAAT2) from apple (cv. golden delicious). Phytochemistry 2006, 67, 658–667. [Google Scholar] [CrossRef]
- Li, R.; Yan, D.; Tan, C.; Li, C.; Song, M.; Zhao, Q.; Yang, Y.; Yin, W.; Liu, Z.; Ren, X.; et al. Transcriptome and metabolomics integrated analysis reveals MdMYB94 associated with esters biosynthesis in apple (Malus × domestica). J. Agric. Food Chem. 2023, 71, 7904–7920. [Google Scholar] [CrossRef]
- Cao, X.; Wei, C.; Duan, W.; Gao, Y.; Kuang, J.; Liu, M.; Chen, K.; Klee, H.; Zhang, B. Transcriptional and epigenetic analysis reveals that NAC transcription factors regulate fruit flavor ester biosynthesis. Plant J. 2021, 106, 785–800. [Google Scholar] [CrossRef]
- Li, L.X.; Fang, Y.; Li, D.; Zhu, Z.H.; Zhang, Y.; Tang, Z.Y.; Li, T.; Chen, X.S.; Feng, S.Q. Transcription factors MdMYC2 and MdMYB85 interact with ester aroma synthesis gene MdAAT1 in apple. Plant Physiol. 2023, 193, 2442–2458. [Google Scholar] [CrossRef]
- Qin, L.; Wei, Q.; Kang, W.; Zhang, Q.; Sun, J.; Liu, S. Comparison of volatile compounds in ‘Fuji’ apples in the different regions in China. Food Sci. Technol. Res. 2017, 23, 79–89. [Google Scholar] [CrossRef]
- Wang, J.; Sun, Q.; Ma, C.; Meng, W.; Wang, C.; Zhao, Y.; Wang, W.; Hu, D. MdWRKY31-MdNAC7 regulatory network: Orchestrating fruit softening by modulating cell wall-modifying enzyme MdXTH2 in response to ethylene signalling. Plant Biotechnol. J. 2024, 22, 3244–3261. [Google Scholar] [CrossRef]
- Zhang, B.; Yan, S.; Wu, S.; Feng, H.; Siddique, K. Organic and inorganic fertilizers combined with a water-saving technique increased soil fertilities and apple production in a rainfed hilly orchard. J. Clean. Prod. 2023, 414, 137647. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, L.; Wang, Y.; Tao, H.; Fan, J.; Zhao, Z.; Guo, Y. Effects of soil water stress on fruit yield, quality and their relationship with sugar metabolism in ‘Gala’ apple. Sci. Hortic. 2019, 258, 108753. [Google Scholar] [CrossRef]
- Mpelasoka, S.; Behboudian, H. Production of aroma volatiles in response to deficit irrigation and to crop load in relation to fruit maturity for ‘Braeburn’ apple. Postharvest Biol. Technol. 2002, 24, 1–11. [Google Scholar] [CrossRef]
- Wang, Z.; Feng, Y.; Wang, H.; Liu, X.; Zhao, Z. Effects of different pre-harvest bagging times on fruit quality of apple. Foods 2024, 13, 1243. [Google Scholar] [CrossRef] [PubMed]
- Watkins, B. Storage technology for apples and pears. Postharvest Biol. Technol. 2002, 24, 193–194. [Google Scholar] [CrossRef]
- Argenta, L.C.; Mattheis, J.P.; Fan, X.; Finger, F.L. Production of volatile comp ounds by Fuji apples following exposure to high CO2 or low O2. J. Agric. Food Chem. 2004, 52, 5957–5963. [Google Scholar] [CrossRef]
- Dias, C.; Ribeiro, T.; Rodrigues, A.C.; Ferrante, A.; Vasconcelos, M.W.; Pintado, M. Improving the ripening process after 1-MCP application: Implications and strategies. Trends Food Sci. Tech. 2021, 113, 382–396. [Google Scholar] [CrossRef]
- Yan, R.; Yun, J.; Gurtler, B.; Fan, X. Radiochromic film dosimetry for UV-C treatments of apple fruit. Postharvest Biol. Technol. 2017, 127, 14–20. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Yao, Y.; Wu, Y.; Yang, S. The Volatile Composition, Biosynthesis Pathways, Breeding Strategies, and Regulation Measures of Apple Aroma: A Review. Horticulturae 2025, 11, 310. https://doi.org/10.3390/horticulturae11030310
Tang Y, Yao Y, Wu Y, Yang S. The Volatile Composition, Biosynthesis Pathways, Breeding Strategies, and Regulation Measures of Apple Aroma: A Review. Horticulturae. 2025; 11(3):310. https://doi.org/10.3390/horticulturae11030310
Chicago/Turabian StyleTang, Yuying, Yijun Yao, Yangyun Wu, and Shunbo Yang. 2025. "The Volatile Composition, Biosynthesis Pathways, Breeding Strategies, and Regulation Measures of Apple Aroma: A Review" Horticulturae 11, no. 3: 310. https://doi.org/10.3390/horticulturae11030310
APA StyleTang, Y., Yao, Y., Wu, Y., & Yang, S. (2025). The Volatile Composition, Biosynthesis Pathways, Breeding Strategies, and Regulation Measures of Apple Aroma: A Review. Horticulturae, 11(3), 310. https://doi.org/10.3390/horticulturae11030310