
Genome-Wide Analysis of NPH3/RPT2-like (NRL) Genes in Grape (Vitis vinifera L.): Their Identification, Characterization, and Different Responses to Light Quality

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification and Characterization of VvNRLs in Grape
2.2. Phylogenetic Analysis of VvNRLs
2.3. Spatiotemporal Expression Profiling During Grapevine Development
2.4. Identification of Cis-Acting Elements in VvNRL Promoters
2.5. Chromosomal Location and Synteny Correlation Analysis
2.6. Protein–Protein Interaction Network and GO Enrichment
2.7. Three-Dimensional Structure and Protein–Protein Docking
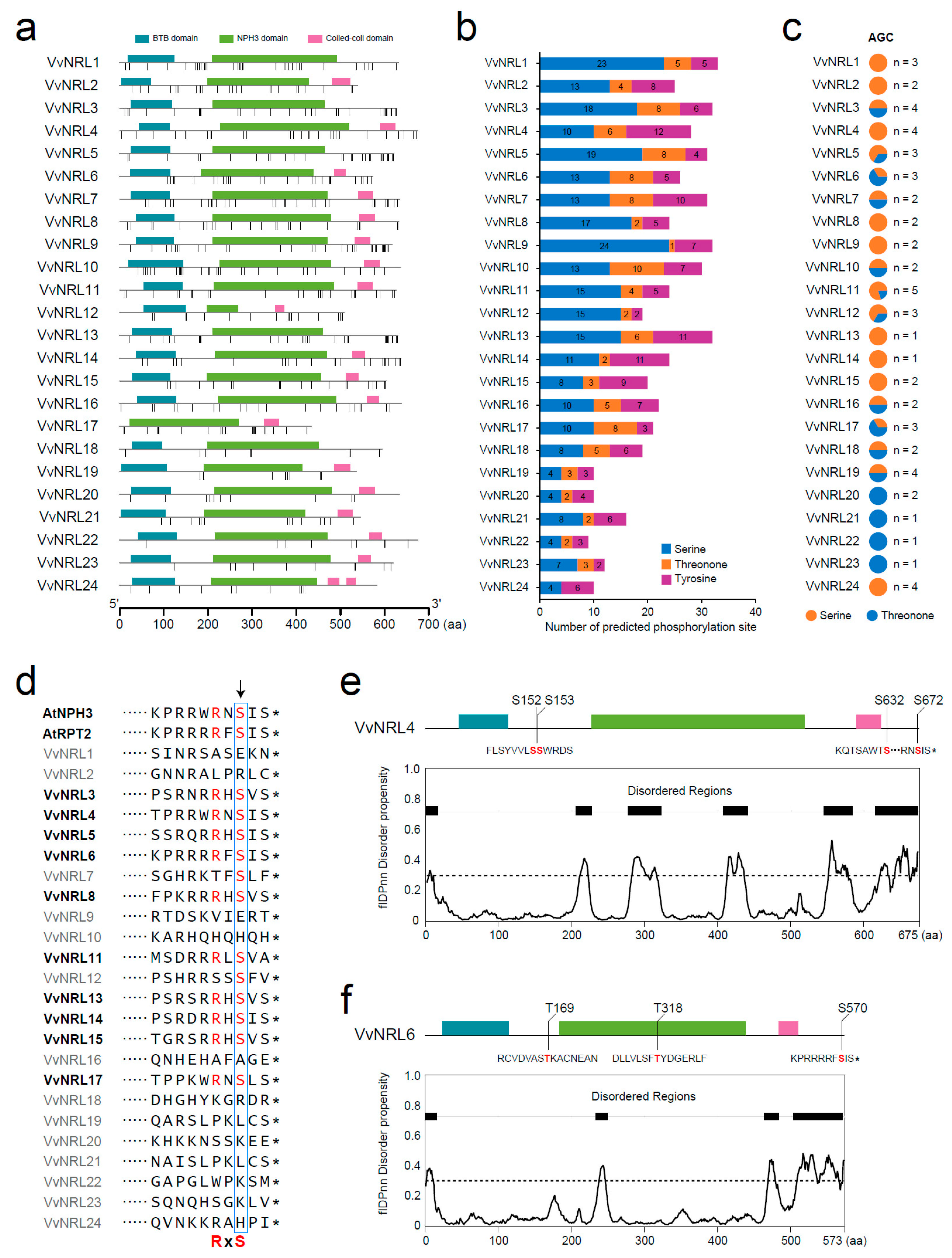
2.8. Prediction of Phosphorylation Sites and Disorder Regions in VvNRLs
3. Results
3.1. Genome-Wide Identification of the VvNRL Family in the Grape Genome
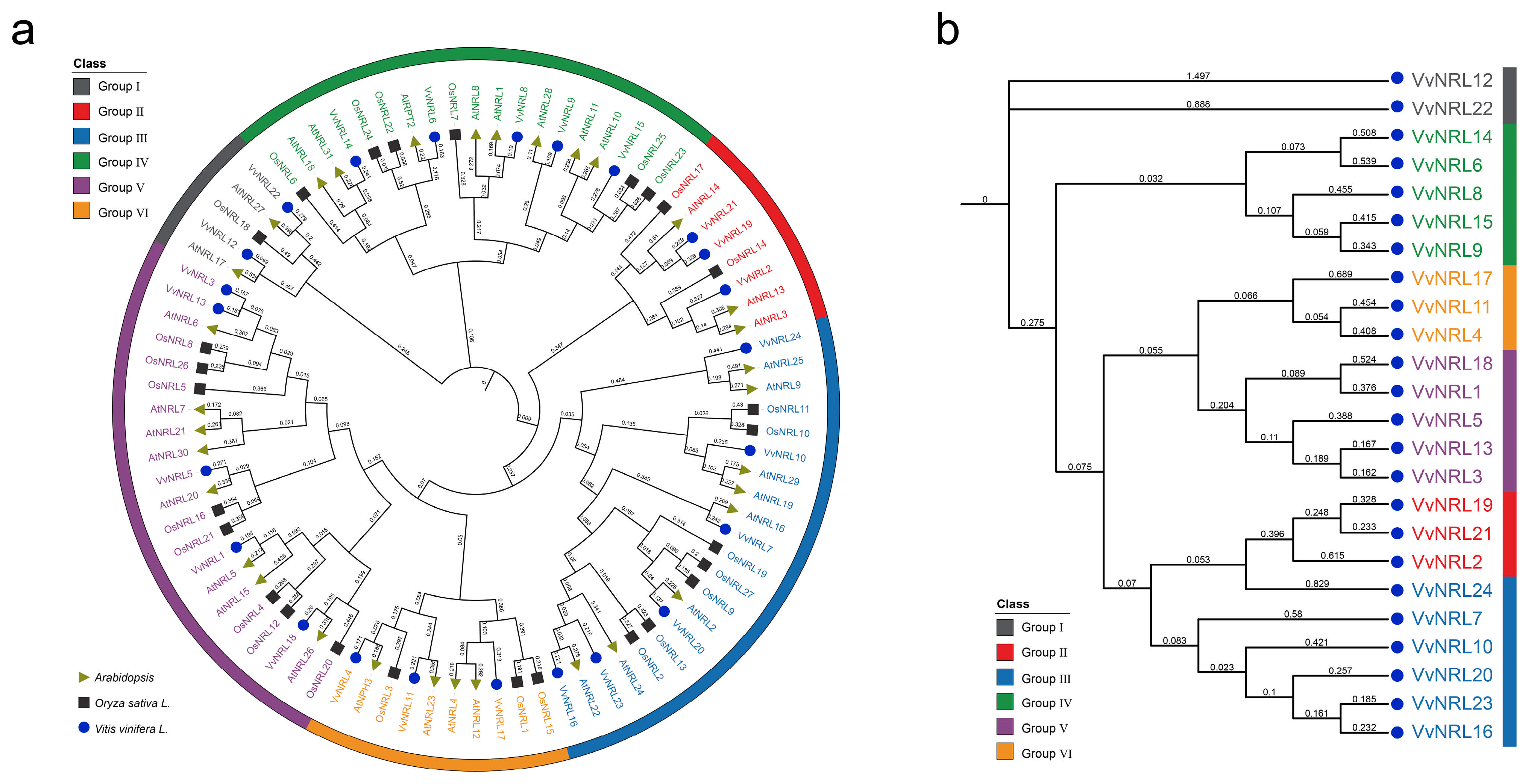
3.2. Phylogenetic Analysis of the Grapevine VvNRL Gene Family’s Evolutionary Relationships
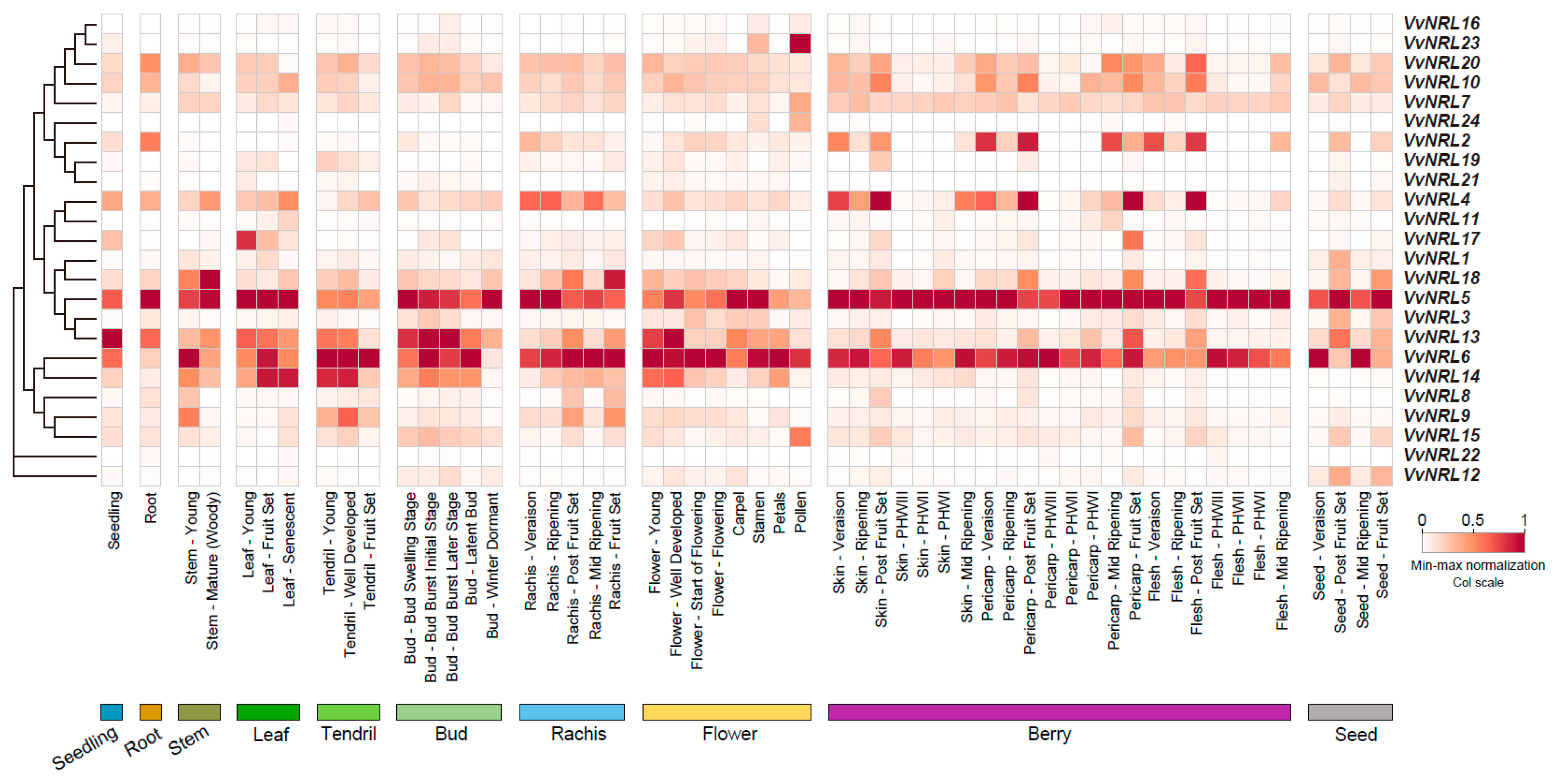
3.3. Expression Pattern Analysis of VvNRL Genes
3.4. Analysis of Cis-Acting Elements in Grapevine NRL Gene Family Promoters
3.5. Segmental Duplication Events Are Observed in VvNRLs
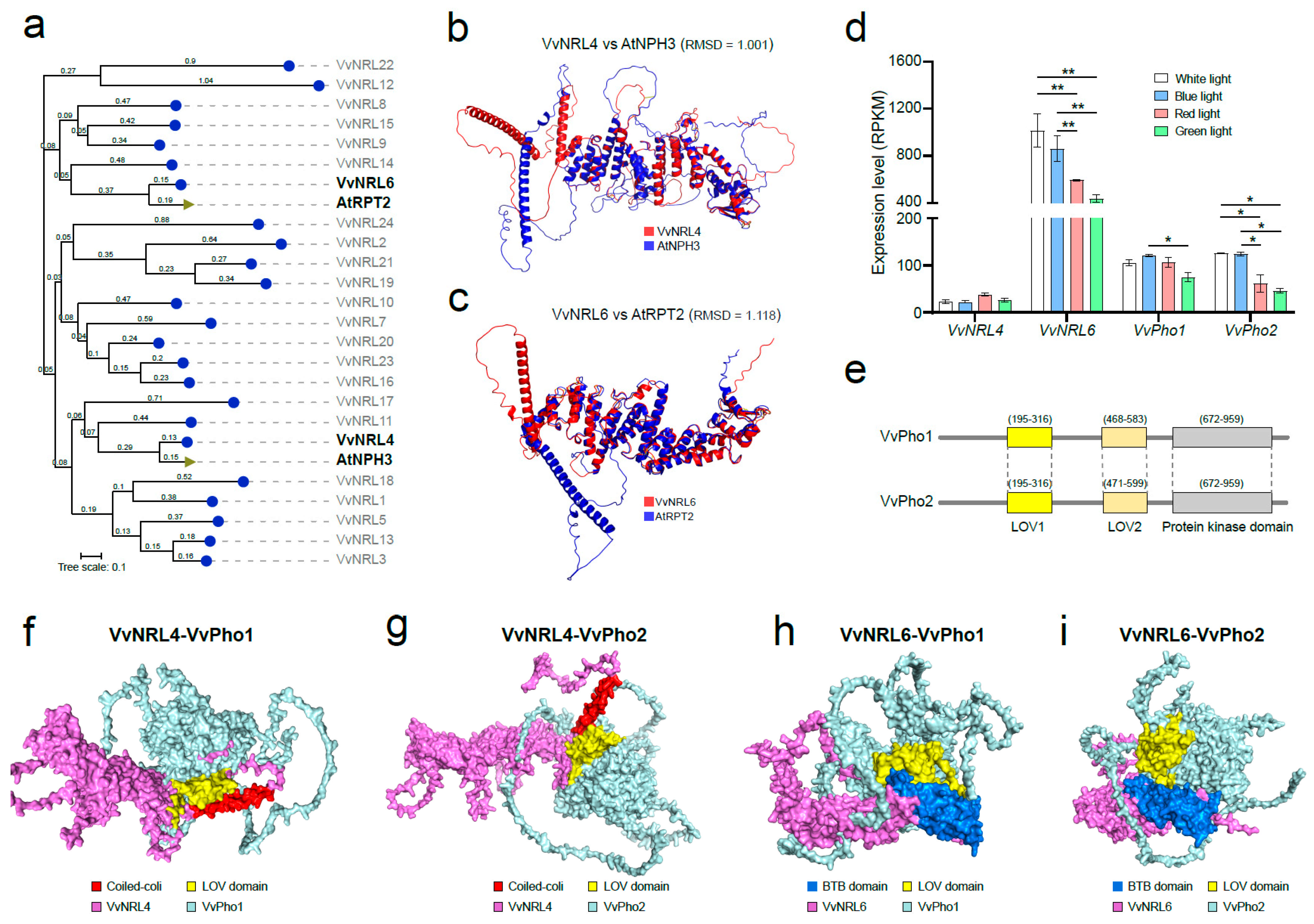
3.6. Protein Docking of VvNRL4, VvNRL6 and VvPHO1, and VvPHO2
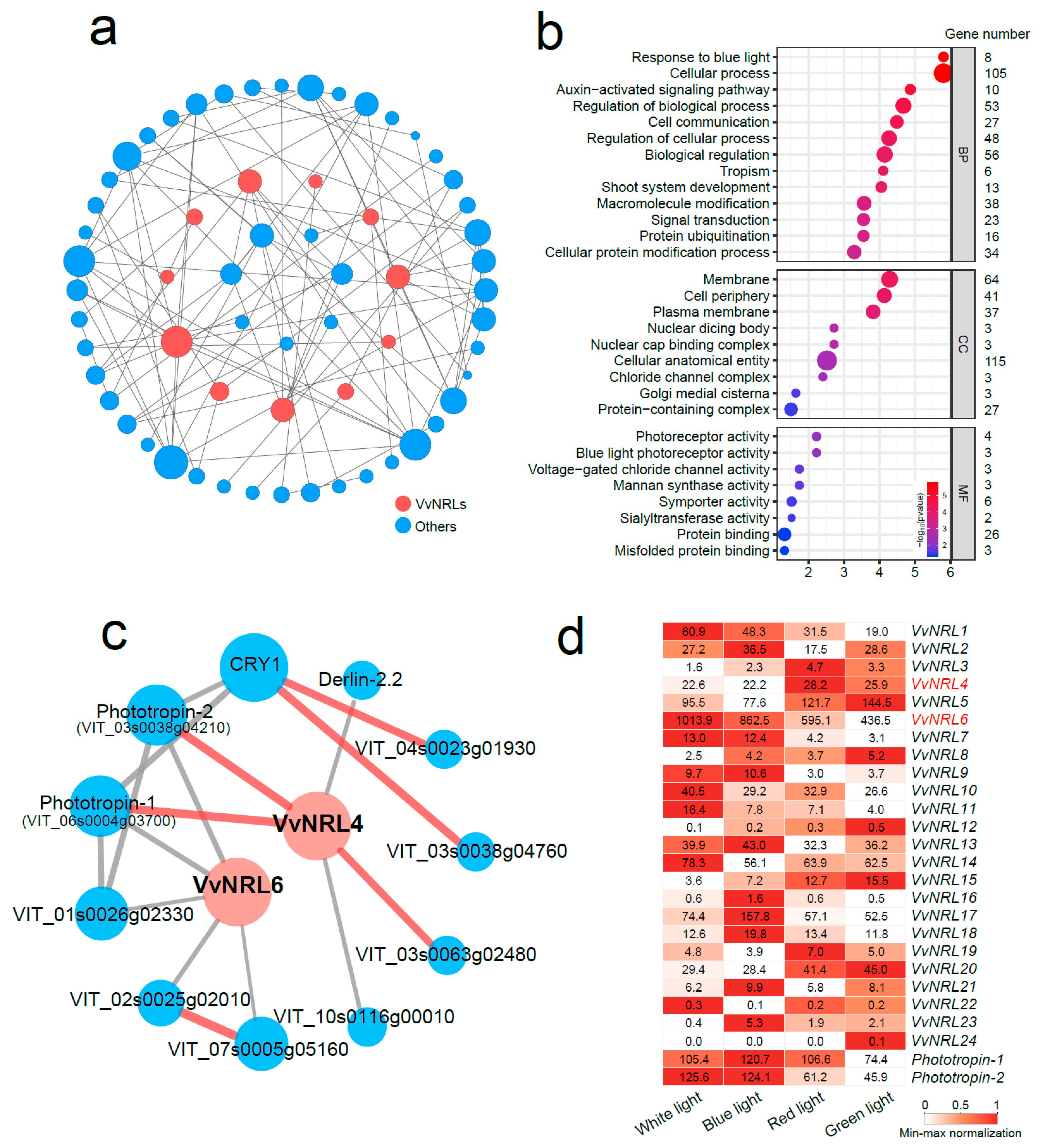
3.7. Network Analysis of VvNRL-Mediated Genes for Development and Light Responsiveness
3.8. Prediction of the Phosphorylation Site of VvNRL Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Holland, J.J.; Roberts, D.; Liscum, E. Understanding Phototropism: From Darwin to Today. J. Exp. Bot. 2009, 60, 1969–1978. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Kagawa, T.; Kasahara, M.; Swartz, T.E.; Christie, J.M.; Briggs, W.R.; Wada, M.; Okada, K. Arabidopsis nph1 and npl1: Blue Light Receptors that Mediate both Phototropism and Chloroplast Relocation. Proc. Natl. Acad. Sci. USA 2001, 98, 6969–6974. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, K.; Sakai, T.; Takagi, S. Blue Light-Dependent Nuclear Positioning in Arabidopsis thaliana Leaf Cells. Plant Cell Physiol. 2007, 48, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- Kagawa, T.; Sakai, T.; Suetsugu, N.; Oikawa, K.; Ishiguro, S.; Kato, T.; Tabata, S.; Okada, K.; Wada, M. Arabidopsis NPL1: A Phototropin Homolog Controlling the Chloroplast High-Light Avoidance Response. Science 2001, 291, 2138–2141. [Google Scholar] [CrossRef]
- Suetsugu, N.; Kagawa, T.; Wada, M. An Auxilin-Like J-Domain Protein, JAC1, Regulates Phototropin-Mediated Chloroplast Movement in Arabidopsis. Plant Physiol. 2005, 139, 151–162. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.L.; Qiao, X.R.; Wang, J.; Wang, L.D.; Xu, C.S.; Zhang, X. Phototropins Function in High-Intensity Blue Light-Induced Hypocotyl Phototropism in Arabidopsis by Altering Cytosolic Calcium. Plant Physiol. 2013, 162, 1539–1551. [Google Scholar] [CrossRef]
- Christie, J.M. Phototropin Blue-Light Receptors. Annu. Rev. Plant Biol. 2007, 58, 21–45. [Google Scholar] [CrossRef]
- Motchoulski, A.; Liscum, E. Arabidopsis NPH3: A NPH1 Photoreceptor-Interacting Protein Essential for Phototropism. Science 1999, 286, 961–964. [Google Scholar] [CrossRef]
- Inada, S.; Ohgishi, M.; Mayama, T.; Okada, K.; Sakai, T. RPT2 Is a Signal Transducer Involved in Phototropic Response and Stomatal Opening by Association with Phototropin 1 in Arabidopsis thaliana. Plant Cell 2004, 16, 887–896. [Google Scholar] [CrossRef]
- Liscum, E.; Askinosie, S.K.; Leuchtman, D.L.; Morrow, J.; Willenburg, K.T.; Coats, D.R. Phototropism: Growing towards an Understanding of Plant Movement. Plant Cell 2014, 26, 38–55. [Google Scholar] [CrossRef]
- Okada, K.; Shimura, Y. Mutational Analysis of Root Gravitropism and Phototropism of Arabidopsis thaliana Seedlings. Aust. J. Plant Physiol. 1992, 19, 439–448. [Google Scholar] [CrossRef]
- Liscum, E.; Briggs, W.R. Mutations in the nph1 Locus of Arabidopsis Disrupt the Perception of Phototropic Stimuli. Plant Cell 1995, 7, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Wada, T.; Ishiguro, S.; Okada, K. RPT2: A Signal Transducer of the Phototropic Response in Arabidopsis. Plant Cell 2000, 12, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Pedmale, U.V.; Celaya, R.B.; Liscum, E. Phototropism: Mechanism and Outcomes. Arab. Book 2010, 2010, e0125. [Google Scholar] [CrossRef] [PubMed]
- Suetsugu, N.; Takemiya, A.; Kong, S.G.; Higa, T.; Komatsu, A.; Shimazaki, K.; Kohchi, T.; Wada, M. RPT2/NCH1 Subfamily of NPH3-Like Proteins Is Essential for the Chloroplast Accumulation Response in Land Plants. Proc. Natl. Acad. Sci. USA 2016, 113, 10424–10429. [Google Scholar] [CrossRef]
- Sullivan, S.; Thomson, C.E.; Kaiserli, E.; Christie, J.M. Interaction Specificity of Arabidopsis 14-3-3 Proteins with Phototropin Receptor Kinases. Febs Lett. 2009, 583, 2187–2193. [Google Scholar] [CrossRef]
- Christie, J.M.; Blackwood, L.; Petersen, J.; Sullivan, S. Plant Flavoprotein Photoreceptors. Plant Cell Physiol. 2015, 56, 401–413. [Google Scholar] [CrossRef]
- Petricka, J.J.; Clay, N.K.; Nelson, T.M. Vein Patterning Screens and the Defectively Organized Tributaries Mutants in Arabidopsis thaliana. Plant J. 2008, 56, 251–263. [Google Scholar] [CrossRef]
- Cheng, Y.F.; Qin, G.J.; Dai, X.H.; Zhao, Y.D. NPY Genes and AGC Kinases Define Two Key Steps in Auxin-Mediated Organogenesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2008, 105, 21017–21022. [Google Scholar] [CrossRef]
- Li, Y.T.; Dai, X.H.; Cheng, Y.F.; Zhao, Y.D. NPY Genes Play an Essential Role in Root Gravitropic Responses in Arabidopsis. Mol. Plant 2011, 4, 171–179. [Google Scholar] [CrossRef]
- Lalanne, E.; Honys, D.; Johnson, A.; Borner, G.H.H.; Lilley, K.S.; Dupree, P.; Grossniklaus, U.; Twell, D. SETH1 and SETH2, Two Components of the Glycosylphosphatidylinositol Anchor Biosynthetic Pathway, Are Required for Pollen Germination and Tube Growth in Arabidopsis. Plant Cell 2004, 16, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wengier, D.; Shuai, B.; Gui, C.P.; Muschietti, J.; McCormick, S.; Tang, W.H. The Pollen Receptor Kinase LePRK2 Mediates Growth-Promoting Signals and Positively Regulates Pollen Germination and Tube Growth. Plant Physiol. 2008, 148, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Christie, J.M.; Suetsugu, N.; Sullivan, S.; Wada, M. Shining Light on the Function of NPH3/RPT2-Like Proteins in Phototropin Signaling. Plant Physiol. 2018, 176, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.T.; Meng, X.C.; Zhang, L.; Zhang, Y.Y.; Cai, Z.R.; Huang, Z.Q.; Su, M.; Wang, Y.; Li, M.Z.; Li, F.Y.; et al. A Mechanically Robust Conducting Polymer Network Electrode for Efficient Flexible Perovskite Solar Cells. Joule 2019, 3, 2205–2218. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Fasoli, M.; Dal Santo, S.; Zenoni, S.; Tornielli, G.B.; Farina, L.; Zamboni, A.; Porceddu, A.; Venturini, L.; Bicego, M.; Murino, V.; et al. The Grapevine Expression Atlas Reveals a Deep Transcriptome Shift Driving the Entire Plant into a Maturation Program. Plant Cell 2012, 24, 3489–3505. [Google Scholar] [CrossRef]
- Li, C.X.; Xu, Z.G.; Dong, R.Q.; Chang, S.X.; Wang, L.Z.; Khalil-Ur-Rehman, M.; Tao, J.M. An RNA-Seq Analysis of Grape Plantlets Grown in Vitro Reveals Different Responses to Blue, Green, Red LED Light, and White Fluorescent Light. Front. Plant Sci. 2017, 8, 78. [Google Scholar] [CrossRef]
- Wang, Y.P.; Tang, H.B.; DeBarry, J.D.; Tan, X.; Li, J.P.; Wang, X.Y.; Lee, T.H.; Jin, H.Z.; Marler, B.; Guo, H.; et al. MCScanX: A Toolkit for Detection and Evolutionary Analysis of Gene Synteny and Collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Yan, Y.M.; Tao, H.Y.; He, J.H.; Huang, S.Y. The HDOCK Server for Integrated Protein–Protein Docking. Nat. Protoc. 2020, 15, 1829–1852. [Google Scholar] [CrossRef]
- Chen, M.M.; Zhang, W.Z.; Gou, Y.J.; Xu, D.Y.; Wei, Y.X.; Liu, D.; Han, C.; Huang, X.H.; Li, C.Z.; Ning, W.S.; et al. GPS 6.0: An Updated Server for Prediction of Kinase-Specific Phosphorylation Sites in Proteins. Nucleic Acids Res. 2023, 51, W243–W250. [Google Scholar] [CrossRef]
- Hu, G.; Katuwawala, A.; Wang, K.; Wu, Z.H.; Ghadermarzi, S.; Gao, J.Z.; Kurgan, L. flDPnn: Accurate Intrinsic Disorder Prediction with Putative Propensities of Disorder Functions. Nat. Commun. 2021, 12, 4438. [Google Scholar] [CrossRef] [PubMed]
- Pedmale, U.V.; Liscum, E. Regulation of Phototropic Signaling in Arabidopsis Via Phosphorylation State Changes in the Phototropin 1-Interacting Protein NPH3. J. Biol. Chem. 2007, 282, 19992–20001. [Google Scholar] [CrossRef] [PubMed]
- Christie, J.M.; Salomon, M.; Nozue, K.; Wada, M.; Briggs, W.R. LOV (Light, Oxygen, or Voltage) Domains of the Dlue-Light Photoreceptor Phototropin (nph1): Binding Sites for the Chromophore Flavin Mononucleotide. Proc. Natl. Acad. Sci. USA 1999, 96, 8779–8783. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, S.; Thomson, C.E.; Lamont, D.J.; Jones, M.A.; Christie, J.M. In Vivo Phosphorylation Site Mapping and Functional Characterization of Arabidopsis Phototropin 1. Mol. Plant 2008, 1, 178–194. [Google Scholar] [CrossRef]
- Inoue, S.; Kinoshita, T.; Takemiya, A.; Doi, M.; Shimazaki, K. Leaf Positioning of Arabidopsis in Response to Blue Light. Mol. Plant 2008, 1, 15–26. [Google Scholar] [CrossRef]
- Kimura, T.; Tsuchida-Mayama, T.; Imai, H.; Okajima, K.; Ito, K.; Sakaia, T. Arabidopsis ROOT PHOTOTROPISM2 Is a Light-Dependent Dynamic Modulator of Phototropin1. Plant Cell 2020, 32, 2004–2019. [Google Scholar] [CrossRef]
- Roberts, D.; Pedmale, U.V.; Morrow, J.; Sachdev, S.; Lechner, E.; Tang, X.B.; Zheng, N.; Hannink, M.; Genschik, P.; Liscum, E. Modulation of Phototropic Responsiveness in Arabidopsis through Ubiquitination of Phototropin 1 by the CUL3-Ring E3 Ubiquitin Ligase CRL3NPH3. Plant Cell 2011, 23, 3627–3640. [Google Scholar] [CrossRef]
- Haga, K.; Tsuchida-Mayama, T.; Yamada, M.; Sakai, T. Arabidopsis ROOT PHOTOTROPISM2 Contributes to the Adaptation to High-Intensity Light in Phototropic Responses. Plant Cell 2015, 27, 1098–1112. [Google Scholar] [CrossRef]
- Liscum, E.; Briggs, W.R. Mutations of Arabidopsis in Potential Transduction and Response Components of the Phototropic Signaling Pathway. Plant Physiol. 1996, 112, 291–296. [Google Scholar] [CrossRef]
- Fankhauser, C.; Christie, J.M. Plant Phototropic Growth. Curr. Biol. 2015, 25, R384–R389. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, Q.P.; Xu, C.Y.; Wang, J.; Zhu, J.D.; Shang, B.S.; Zhang, X. Phot2-Regulated Relocation of NPH3 Mediates Phototropic Response to High-Intensity Blue Light in Arabidopsis thaliana. J. Integr. Plant Biol. 2018, 60, 562–577. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, S.; Waksman, T.; Henderson, L.; Paliogianni, D.; Lütkemeyer, M.; Suetsugu, N.; Christie, J.M. Regulation of Plant Phototropic Growth by NPH3/RPT2-Like Substrate Phosphorylation and 14-3-3 Binding. Nat. Commun. 2021, 12, 6129. [Google Scholar] [CrossRef] [PubMed]
- Fankhauser, C.; Chory, J. Light Control of Plant Development. Annu. Rev. Cell Dev. Biol. 1997, 13, 203–229. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.E.; Dyson, H.J. Intrinsically Disordered Proteins in Cellular Signalling and Regulation. Nat. Rev. Mol. Cell Biol. 2015, 16, 18–29. [Google Scholar] [CrossRef]
- Bah, A.; Forman-Kay, J.D. Modulation of Intrinsically Disordered Protein Function by Post-Translational Modifications. J. Biol. Chem. 2016, 291, 6696–6705. [Google Scholar] [CrossRef]
- Cuevas-Velazquez, C.L.; Dinneny, J.R. Organization out of Disorder: Liquid–Liquid Phase Separation in Plants. Curr. Opin. Plant Biol. 2018, 45, 68–74. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Chr | Start | Stop | Gene ID (Release 56) | Gene ID (12 X) | Protein Size (aa) | pI | MW (KDa) | Localization Prediction |
|---|---|---|---|---|---|---|---|---|---|
| VvNRL1 | 1 | 7,262,915 | 7,266,769 | Vitvi01g00644 | VIT_01s0137g00820 | 631 | 8.78 | 70.08 | Cell membrane. |
| VvNRL2 | 1 | 18,575,511 | 18,577,731 | Vitvi01g01404 | VIT_01s0010g01390 | 539 | 8.82 | 60.30 | Cell membrane. Nucleus. |
| VvNRL3 | 3 | 7,127,155 | 7,138,732 | Vitvi03g00629 | VIT_03s0091g00680 | 627 | 6.85 | 68.89 | Cell membrane. Chloroplast. |
| VvNRL4 | 3 | 13,350,055 | 13,353,833 | Vitvi03g00987 | VIT_03s0038g00270 | 675 | 8.15 | 75.72 | Cell membrane. |
| VvNRL5 | 4 | 6,566,953 | 6,574,324 | Vitvi04g00599 | VIT_04s0008g06630 | 621 | 9.02 | 69.46 | Cell membrane. |
| VvNRL6 | 6 | 8,945,384 | 8,947,855 | Vitvi06g00814 | VIT_06s0004g08230 | 573 | 6.88 | 63.72 | Cell membrane. |
| VvNRL7 | 6 | 18,792,864 | 18,803,839 | Vitvi06g01942 | VIT_06s0061g01140 | 634 | 8.12 | 70.90 | Cell membrane. |
| VvNRL8 | 7 | 2,839,057 | 2,843,199 | Vitvi07g00260 | VIT_07s0104g01440 | 632 | 8.95 | 70.48 | Cell membrane. |
| VvNRL9 | 7 | 11,773,033 | 11,779,092 | Vitvi07g00905 | VIT_07s0005g06300 | 617 | 6.51 | 69.52 | Cell membrane. Nucleus. |
| VvNRL10 | 7 | 20,370,199 | 20,372,852 | Vitvi07g01277 | VIT_00s0207g00230 | 636 | 5.12 | 70.74 | Cell membrane. Nucleus. |
| VvNRL11 | 7 | 21,537,862 | 21,551,645 | Vitvi07g01380 | VIT_00s0665g00050 | 626 | 6 | 70.78 | Cell membrane. Nucleus. |
| VvNRL12 | 7 | 23,884,253 | 23,886,726 | Vitvi07g01591 | VIT_07s0129g00150 | 508 | 5.8 | 55.90 | Cell membrane. Nucleus. |
| VvNRL13 | 7 | 23,977,616 | 23,985,606 | Vitvi07g01603 | VIT_07s0129g00250 | 630 | 6.37 | 69.69 | Cell membrane. |
| VvNRL14 | 7 | 24,100,049 | 24,103,874 | Vitvi07g01617 | VIT_07s0129g00380 | 637 | 6.47 | 70.96 | Cell membrane. |
| VvNRL15 | 8 | 903,840 | 907,145 | Vitvi08g00052 | VIT_08s0056g00610 | 603 | 6.19 | 66.97 | Cell membrane. |
| VvNRL16 | 8 | 12,930,734 | 12,936,077 | Vitvi08g01032 | VIT_08s0040g01800 | 638 | 5.69 | 71.57 | Cell membrane. Nucleus. |
| VvNRL17 | 9 | 5,439,739 | 5,441,660 | Vitvi09g00488 | VIT_09s0002g05700 | 434 | 9.17 | 48.42 | Cell membrane. Chloroplast. |
| VvNRL18 | 10 | 2,389,744 | 2,392,079 | Vitvi10g00220 | VIT_00s0194g00080 | 594 | 8.73 | 67.31 | Cell membrane. Nucleus. |
| VvNRL19 | 10 | 3,546,476 | 3,549,309 | Vitvi10g02177 | VIT_00s0361g00090 | 536 | 8.57 | 59.37 | Cell membrane. Nucleus. |
| VvNRL20 | 10 | 12,613,320 | 12,619,423 | Vitvi10g00892 | VIT_10s0003g04490 | 633 | 6.16 | 69.95 | Cell membrane. Nucleus. |
| VvNRL21 | 12 | 7,540,897 | 7,544,207 | Vitvi12g00572 | VIT_12s0134g00010 | 545 | 8.81 | 60.79 | Cell membrane. Nucleus. |
| VvNRL22 | 12 | 8,719,997 | 8,722,329 | Vitvi12g00658 | VIT_12s0057g00510 | 675 | 8.97 | 76.31 | Cell membrane. Nucleus. |
| VvNRL23 | 13 | 5,909,622 | 5,914,496 | Vitvi13g00618 | VIT_13s0019g04420 | 620 | 5.46 | 68.89 | Cell membrane. Nucleus. |
| VvNRL24 | 15 | 18,253,865 | 18,256,619 | Vitvi15g00905 | VIT_15s0048g02810 | 582 | 5.75 | 65.39 | Cell membrane. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Gao, H.; Li, X.; Liu, Y.; Zhao, H.; Qiu, N.; Zhang, H. Genome-Wide Analysis of NPH3/RPT2-like (NRL) Genes in Grape (Vitis vinifera L.): Their Identification, Characterization, and Different Responses to Light Quality. Horticulturae 2025, 11, 274. https://doi.org/10.3390/horticulturae11030274
Li S, Gao H, Li X, Liu Y, Zhao H, Qiu N, Zhang H. Genome-Wide Analysis of NPH3/RPT2-like (NRL) Genes in Grape (Vitis vinifera L.): Their Identification, Characterization, and Different Responses to Light Quality. Horticulturae. 2025; 11(3):274. https://doi.org/10.3390/horticulturae11030274
Chicago/Turabian StyleLi, Shiyang, Hongsheng Gao, Xiaoyan Li, Yihua Liu, Hang Zhao, Nianwei Qiu, and Hongxia Zhang. 2025. "Genome-Wide Analysis of NPH3/RPT2-like (NRL) Genes in Grape (Vitis vinifera L.): Their Identification, Characterization, and Different Responses to Light Quality" Horticulturae 11, no. 3: 274. https://doi.org/10.3390/horticulturae11030274
APA StyleLi, S., Gao, H., Li, X., Liu, Y., Zhao, H., Qiu, N., & Zhang, H. (2025). Genome-Wide Analysis of NPH3/RPT2-like (NRL) Genes in Grape (Vitis vinifera L.): Their Identification, Characterization, and Different Responses to Light Quality. Horticulturae, 11(3), 274. https://doi.org/10.3390/horticulturae11030274