



Morphological, Physiological and Biochemical Changes in the Grape Variety “Hotan Red” Caused by the Occurrence of Stress Under the Influence of Saline–Alkaline Growing Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Measurement of Morphological Indicators
the corresponding class)/(Supreme class × Total number of plants investigated)
2.3. Determination of Chlorophyll Fluorescence and Photosynthetic Parameters
2.4. Determination of Osmotic Regulators
2.5. Determination of Oxidation Product Content and Antioxidant Enzyme Activity
2.6. Statistical Analysis and Visualisation of Data
3. Results
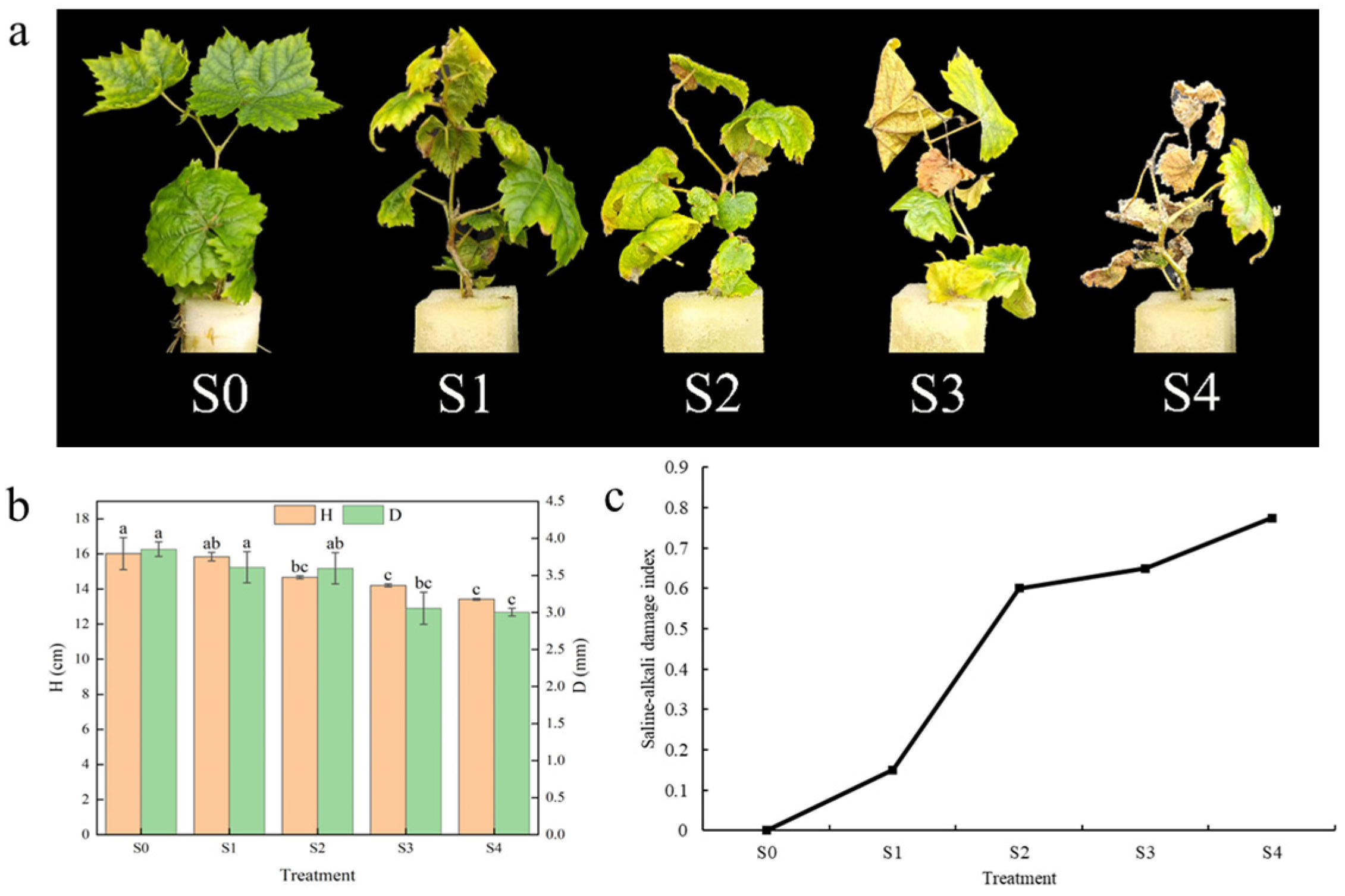
3.1. Effects of Saline–Alkali Stress on Morphological Characteristics of Hotan Red
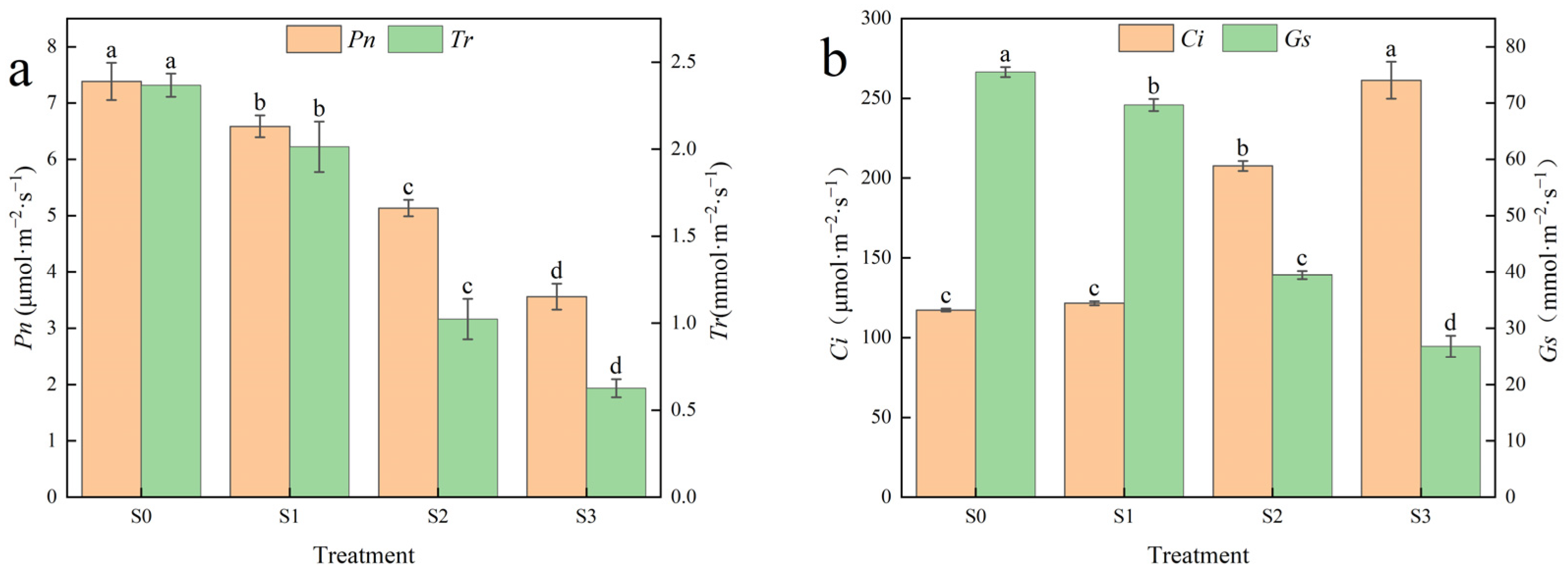
3.2. Effects of Saline–Alkali Stress on Photosynthetic Parameters of Hotan Red
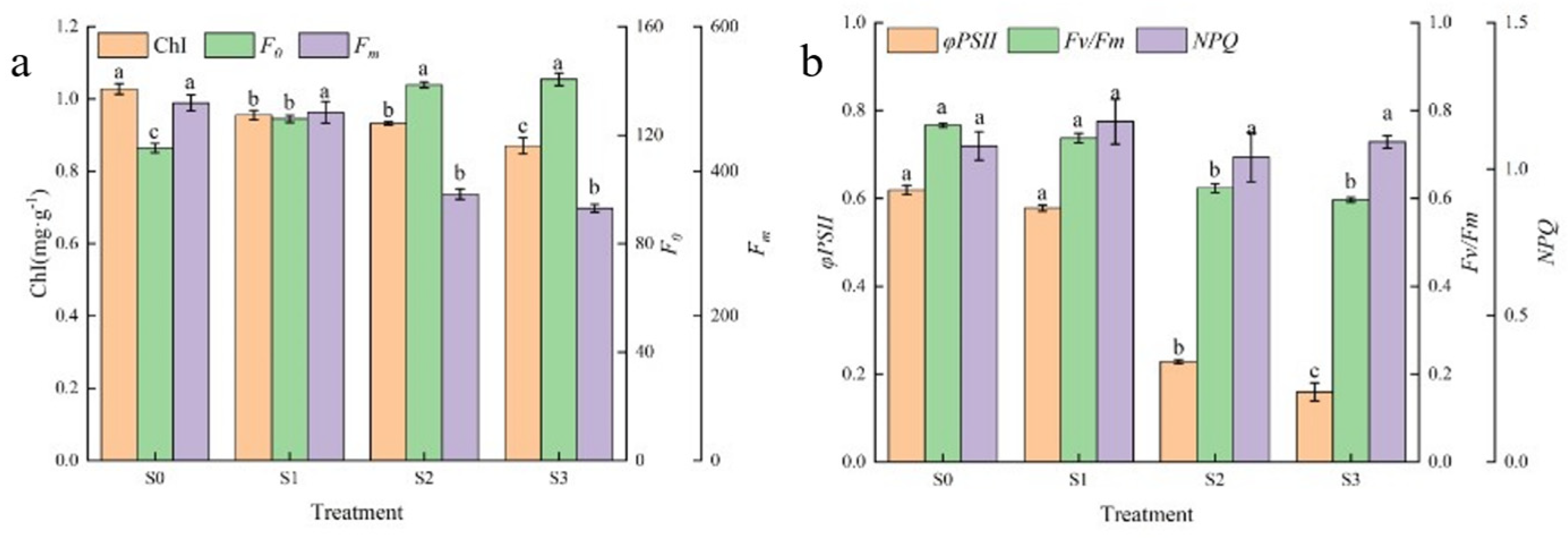
3.3. Effect of Saline–Alkali Stress on Chlorophyll Content and Fluorescence Parameters of Hotan Red
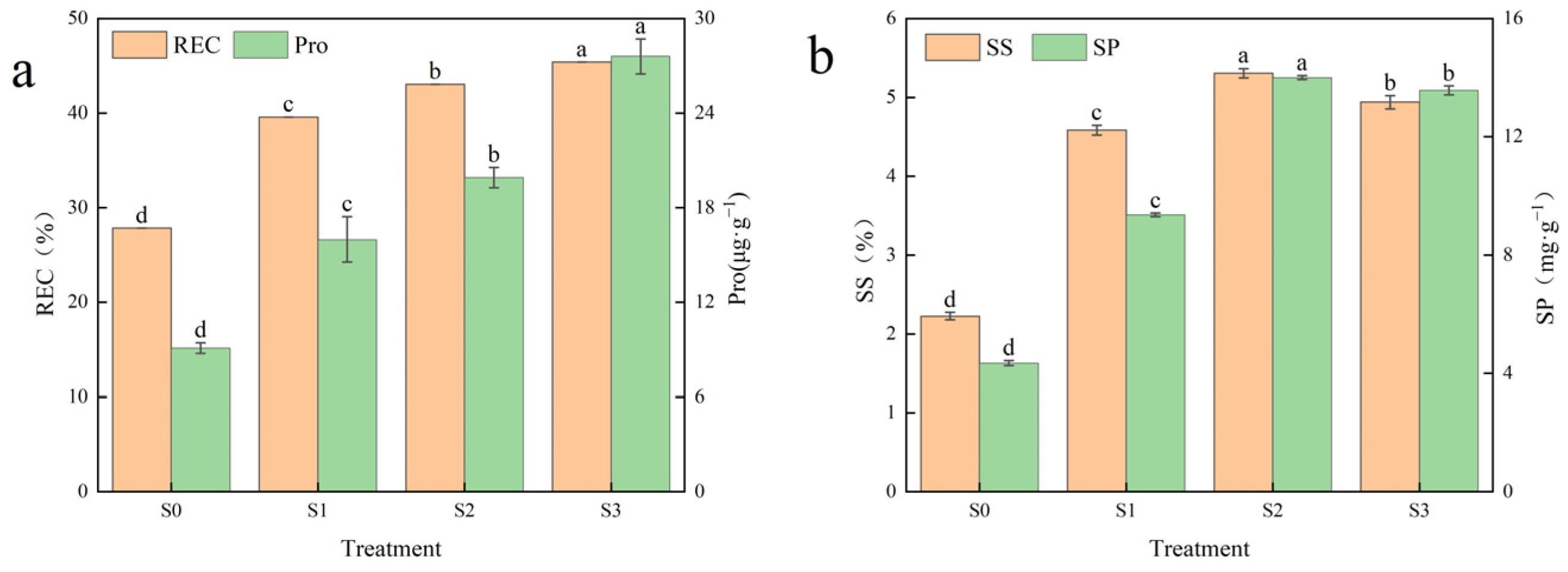
3.4. Effects of Saline–Alkali Stress on Osmoregulation in Hotan Red
3.5. Effect of Saline–Alkali Stress on Oxidation Products of Hotan Red
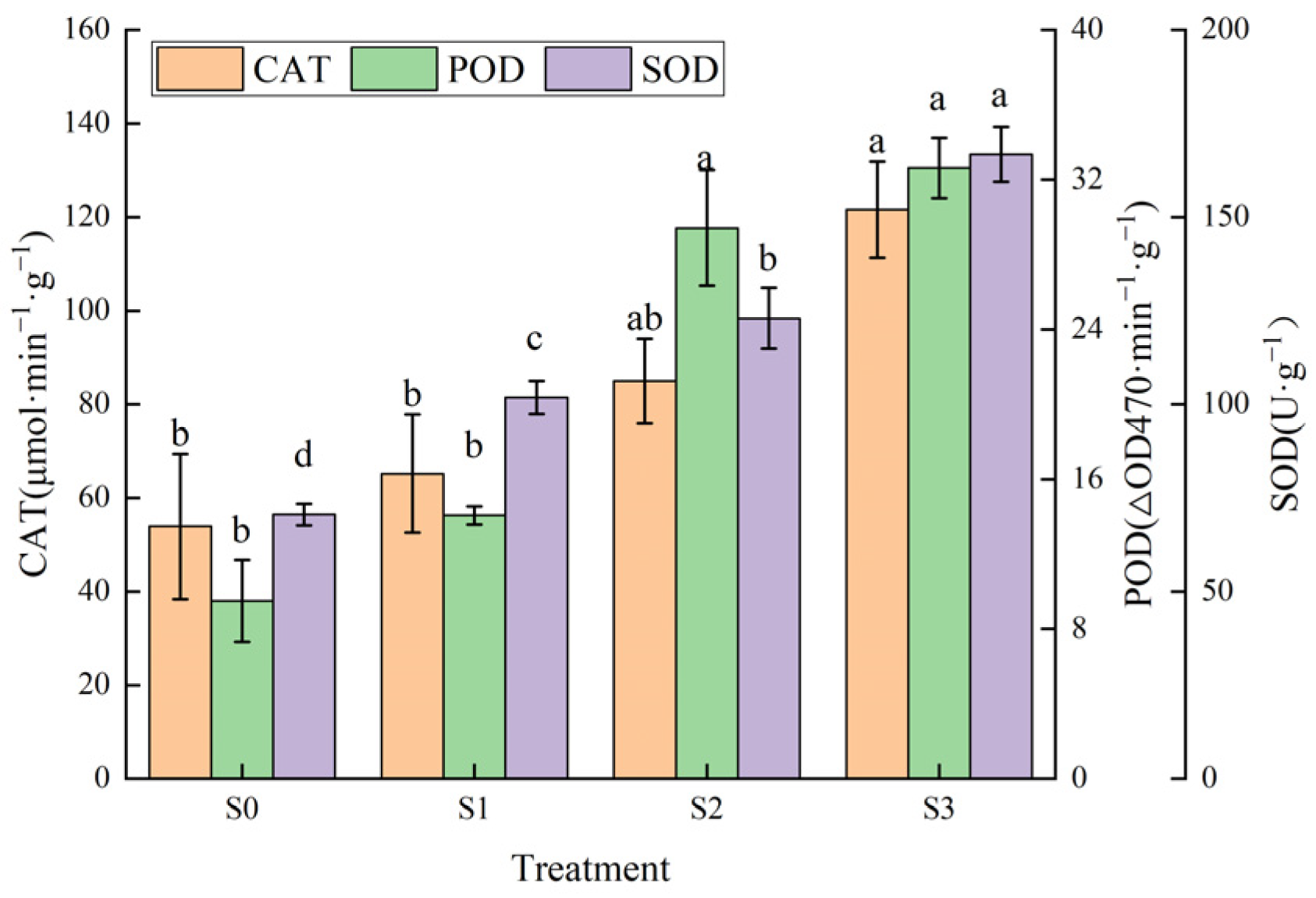
3.6. Effects of Saline–Alkali Stress on Antioxidant Enzyme Activities of Hotan Red
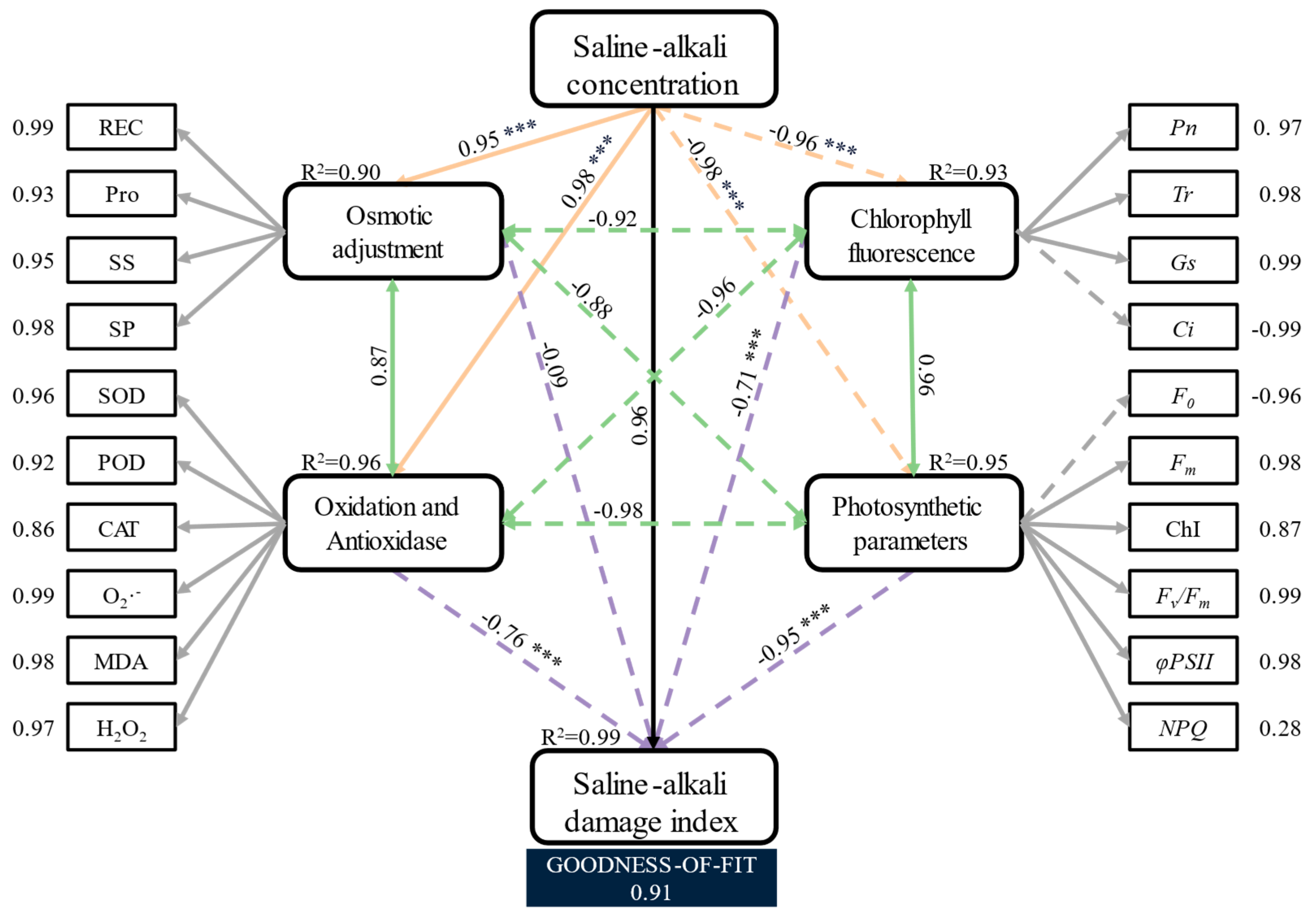
3.7. Partial Least Squares Path Modelling Analysis of the Components of the Saline–Alkali Damage Index in Hotan Red
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sun, Z.; Liu, W.; Zheng, X.; Zheng, X.; Gong, X.; Ma, C.; Liu, X.; Wang, C.; Tian, Y. Effects and Functional Mechanism of Melatonin on the Growth of Malus hupehensis Seedlings Under Saline-Alkali Stress. Acta Hortic. Sin. 2023, 50, 1697–1710. [Google Scholar] [CrossRef]
- Lim, S.J.; Shin, M.N.; Son, J.K.; Song, J.D.; Cho, J.Y. Evaluation of soil pore-water salinity using a Decagon GS3 sensor in saline-alkali reclaimed tidal lands. Comput. Electron. Agric. 2017, 132, 49–55. [Google Scholar] [CrossRef]
- Wei, B. Distribution and genetic analysis of saline-alkali soil in China. Technol. Soil Water Conserv. 2012, 6, 27–28. [Google Scholar]
- Guo, R.; Yang, Z.; Li, F.; Yan, C.; Zhong, X.; Liu, Q.; Xia, X.; Li, H.; Zhao, L. Comparative metabolic responses and adaptive strategies of wheat (Triticum aestivum) to salt and alkali stress. BMC Plant Biol. 2015, 15, 170. [Google Scholar] [CrossRef]
- Song, L.; Yu, Y.; Chen, H.; Feng, Y.; Chen, S.; Zhang, H.; Zhou, H.; Meng, L.; Wang, Y. Response of photosynthetic characteristics and antioxidant system in the leaves of safflower to NaCl and NaHCO3. Plant Cell Rep. 2024, 43, 146. [Google Scholar] [CrossRef]
- Pang, Q.; Zhang, A.; Zang, W.; Wei, L.; Yan, X. Integrated proteomics and metabolomics for dissecting the mechanism of global responses to salt and alkali stress in Suaeda corniculata. Plant Soil 2016, 402, 379–394. [Google Scholar] [CrossRef]
- Yin, Z.; Zhang, H.; Zhao, Q.; Yoo, M.J.; Zhu, N.; Yu, J.; Yu, J.; Guo, S.; Miao, Y.; Chen, S.; et al. Physiological and comparative proteomic analyses of saline-alkali NaHCO3-responses in leaves of halophyte Puccinellia tenuiflora. Plant Soil 2019, 437, 137–158. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Q.; Feng, Y.; Dong, Y.; Zhang, Z.; Wang, Y.; Liu, W. Responsive mechanism of Hemerocallis citrina Baroni to complex saline-alkali stress revealed by photosynthetic characteristics and antioxidant regulation. Plant Cell Rep. 2024, 43, 176. [Google Scholar] [CrossRef]
- Zhao, Q.; Chen, S.; Wang, G.; Du, Y.; Zhang, Z.; Yu, G.; Ren, C.; Zhang, Y.; Du, J. Exogenous melatonin enhances soybean (Glycine max (L.) Merr.) seedling tolerance to saline-alkali stress by regulating antioxidant response and DNA damage repair. Physiol. Plant. 2022, 174, e13731. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Z.; Sui, N. Sensitivity and responses of chloroplasts to salt stress in plants. Front. Plant Sci. 2024, 15, 1374086. [Google Scholar] [CrossRef]
- Zargar, S.M.; Zargar, M.Y. Abiotic Stress-Mediated Sensing and Signaling in Plants: An Omics Perspective || Multifarious Role of ROS in Halophytes: Signaling and Defense; Springer: Berlin/Heidelberg, Germany, 2018; pp. 207–223. [Google Scholar] [CrossRef]
- Rangani, J.; Parida, A.K.; Panda, A.; Kumari, A. Coordinated Changes in Antioxidative Enzymes Protect the Photosynthetic Machinery from Salinity Induced Oxidative Damage and Confer Salt Tolerance in an Extreme Halophyte Salvadora persica L. Front. Plant Sci. 2016, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, S.; Tang, T.; Liu, Y.; Mobeen, T.M.; Wang, C.; Meng, Z.; Niu, J.; Yang, W.; Ma, J. Comparison of morphological, physiological, and related-gene expression responses to saline-alkali stress in eight apple rootstock genotypes. Sci. Hortic. 2022, 306, 111455. [Google Scholar] [CrossRef]
- Zhong, Y.; Qi, X.; Chen, J.; Li, Z.; Bai, D. Growth and physiological responses of four kiwifruit genotypes to salt stress and resistance evaluation. J. Integr. Agric. 2019, 18, 87–99. [Google Scholar] [CrossRef]
- Wei, J.; Liu, D.; Liu, Y.; Wei, S. Physiological Analysis and Transcriptome Sequencing Reveal the Effects of Salt Stress on Banana (Musa acuminata cv. BD) Leaf. Front. Plant Sci. 2022, 13, 822838. [Google Scholar] [CrossRef]
- Boughalleb, F.; Abdellaoui, R.; Mahmoudi, M.; Bakhshandeh, E. Changes in phenolic profile, soluble sugar, proline, and antioxidant enzyme activities of Polygonumequisetiforme in response to salinity. Turk. J. Bot. 2020, 44, 25–35. [Google Scholar] [CrossRef]
- Amini, F.; Ehsanpour, A.A. Soluble Proteins, Proline, Carbohydrates and Na+/K+ Changes in Two Tomato (Lycopersicon esculentum Mill.) Cultivars under in vitro Salt Stress. Am. J. Biochem. Biotechnol. 2005, 1, 212–216. [Google Scholar] [CrossRef]
- Ma, Y.; Riziwangguli, A.; Bai, S.; Zhong, H.; Zhang, F.; Hu, J.; Wu, X.; Wu, J. Analyze on development of grape industry in Xinjiang. China Fruits 2024, 9, 102–108. [Google Scholar] [CrossRef]
- Zhao, J.; Ma, L.; Zhang, Y.; Bahar, G. An experimental study of antitumor effects on Xinjiang Hotan Red grape in vitro. J. Xinjiang Med. Univ. 2001, 2, 95–97. [Google Scholar]
- Wang, F.; Yang, S.; Wei, Y.; Shi, Q.; Ding, J. Characterizing soil salinity at multiple depth using electromagnetic induction and remote sensing data with random forests: A case study in Tarim River Basin of southern Xinjiang, China. Sci. Total Environ. 2021, 754, 142030. [Google Scholar] [CrossRef]
- Du, Z.; Zhai, H.; Luo, X.; Pan, Z.; Cheng, S. Salt-Tolerance Identification on Apple Rootstocks. J. Fruit Sci. 2002, 1, 4–7. [Google Scholar] [CrossRef]
- Hussain, S.; Huang, J.; Zhu, C.; Zhu, L.; Cao, X.; Hussain, S.; Ashraf, M.; Khaskheli, M.A.; Kong, Y.; Jin, Q.; et al. Pyridoxal 5’-phosphate enhances the growth and morpho-physiological characteristics of rice cultivars by mitigating the ethylene accumulation under salinity stress. Plant Physiol. Biochem. 2020, 154, 782–795. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jia, K.; Tian, Y.; Han, K.; El-Kassaby, Y.A.; Yang, H.; Si, H.; Sun, Y.; Li, Y. Time-course transcriptomics analysis reveals key responses of populus to salt stress. Ind. Crop. Prod. 2023, 194, 116278. [Google Scholar] [CrossRef]
- Vieira, S.M.A.; Silva, T.M.; Glória, M.B.A. Influence of processing on the levels of amines and proline and on the physicochemical characteristics of concentrated orange juice. Food Chem. 2010, 119, 7–11. [Google Scholar] [CrossRef]
- Campion, E.M.; Walls, D.; Loughran, S.T. Protein Quantitation and Analysis of Purity. Methods Mol. Biol. 2023, 2699, 305–347. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Wang, X.; Li, Y.; Peng, F.; Wang, L. Regulation of POD activity by pelargonidin during vegetative growth in radish (Raphanus sativus L.). Sci. Hortic. 2014, 174, 105–111. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, R.; Gao, S.; Pan, Z.; Guo, Z.; Yu, S.; Wang, Y.; Jin, Q.; Chen, X.; Zhang, L. Transcriptome analysis and phenotyping of walnut seedling roots under nitrogen stresses. Sci. Rep. 2022, 12, 12066. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Y.; Zhu, J.K. Thriving under Stress: How Plants Balance Growth and the Stress Response. Dev. Cell 2020, 55, 529–543. [Google Scholar] [CrossRef]
- Yu, X.; Yue, Q.; Yu, M.; Du, Y.; Yao, Y. Physiological responses of grape rootstocks SA15, SA17 and 1103P to salt-alkali stress. Plant Physiol. J. 2020, 56, 57–65. [Google Scholar] [CrossRef]
- Lu, Y.; Li, M.; Chen, X. Effect of saline-alkali stress on the growth and related physiological indicators of ‘Cabernet Sauvignon’ grape seedlings. Acta Bot. Boreali-Occident. Sin. 2024, 44, 182–193. [Google Scholar]
- Ji, X.; Tang, J.; Zhang, J. Effects of Salt Stress on the Morphology, Growth and Physiological Parameters of Juglansmicrocarpa L. Seedlings. Plants 2022, 11, 2381. [Google Scholar] [CrossRef]
- Xu, J.; Sui, C.; Ge, J.; Ren, R.; Pang, Y.; Gan, H.; Du, Y.; Cao, H.; Sun, Q. Exogenous spermidine improved the salinity-alkalinity stress tolerance of grapevine (Vitis vinifera) by regulating antioxidant system, Na+/K+ homeostasis and endogenous polyamine contents. Sci. Hortic. 2024, 326, 112725. [Google Scholar] [CrossRef]
- Qin, L.; Kang, W.; Qi, Y.; Cai, A. Effects of Salt Stress on Mesophyll Cell Structures and Photosynthetic Characteristics in Leaves of Wine Grape (Vitis spp.). Sci. Agric. Sin. 2012, 45, 4233–4241. [Google Scholar]
- Xiong, H.; Tian, X.; Mao, P.; Zheng, L.; Fu, J.; Meng, L. Effects of Exogenous Melatonin on Photosynthetic and Physiological Characteristics of White Clover Seedlings under Salt Stress. Pratacultural Sci. 2024, 1–20. [Google Scholar]
- Wang, W.; Gao, L.; Zhang, R.; Zhao, T.; Zhang, Z.; Wang, S.; Wang, Y. Effects of 2,4 ep brassinolide on photosynthetic and physiological characteristics of Malusa halliana under saline-alkali stress. J. Fruit Sci. 2021, 38, 1479–1490. [Google Scholar] [CrossRef]
- Lu, Q.; Chen, L.; Ma, Y.; Liu, Y.; Zhao, Y.; Zhao, B.; Sun, J. Effects of saline-alkali stress on photosynthetic and chlorophyll fluorescence characteristics of different grape rootstocks. J. Fruit Sci. 2022, 39, 773–783. [Google Scholar] [CrossRef]
- Niu, M.; Xie, J.; Chen, C.; Cao, H.; Sun, J.; Kong, Q.; Shabala, S.; Shabala, L.; Huang, Y.; Bie, Z. An early ABA-induced stomatal closure, Na+ sequestration in leaf vein and K+ retention in mesophyll confer salt tissue tolerance in Cucurbita species. J. Exp. Bot. 2018, 69, 4945–4960. [Google Scholar] [CrossRef]
- Geilfus, C.M.; Mithöfer, A.; Ludwig-Müller, J.; Zörb, C.; Muehling, K.H. Chloride-inducible transient apoplastic alkalinizations induce stomata closure by controlling abscisic acid distribution between leaf apoplast and guard cells in salt-stressed Vicia faba. New Phytol. 2015, 208, 803–816. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Shabala, L.; Zhou, M.; Brodribb, T.J.; Corkrey, R.; Shabala, S. Factors determining stomatal and non-stomatal (residual) transpiration and their contribution towards salinity tolerance in contrasting barley genotypes. Environ. Exp. Bot. 2018, 153, 10–20. [Google Scholar] [CrossRef]
- Nefissi, O.R.; Arasappan, D.; Ruhlman, T.A.; Ben, C.M.; Abid, G.; Mejri, S.; Ghorbel, A.; Jansen, R.K. Effects of Salt Stress on Transcriptional and Physiological Responses in Barley Leaves with Contrasting Salt Tolerance. Int. J. Mol. Sci. 2022, 23, 5006. [Google Scholar] [CrossRef]
- Abdelgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High Salinity Induces Different Oxidative Stress and Antioxidant Responses in Maize Seedlings Organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef]
- Martin, R.B.F. Rootstocks increase grapevine tolerance to NaCl through ion compartmentalization and exclusion. Acta Physiol. Plant 2020, 42, 1–11. [Google Scholar] [CrossRef]
- Song, C.; Dong, S.; Schlisser, A.; Lupo, Y.; Rachmilevitch, S.; Lazarovitch, N.; Fait, A. Rootstock varietal ability in accumulation of chloride ions underpins improved physiology and metabolism of grapevine exposed to salinity. Sci. Hortic. 2024, 328, 112964. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, Y.; Hu, Y.; Jia, X.; Zhao, T.; Cheng, L.; Wang, Y. Tolerance of two apple rootstocks to short-term salt stress: Focus on chlorophyll degradation, photosynthesis, hormone and leaf ultrastructures. Acta Physiol. Plant 2019, 41, 1–14. [Google Scholar] [CrossRef]
- Shankar, K.; Awasthi, O.P.; Dubey, A.K. Rootstock mediated alteration in morphology and photosystem in sweet orange (Citrus sinensis) scion cv. Pusa Sharad under NaCl stress. Indian J. Agric. Sci. 2023, 93, 1103–1107. [Google Scholar] [CrossRef]
- Stefanov, M.A.; Rashkov, G.D.; Apostolova, E.L. Assessment of the Photosynthetic Apparatus Functions by Chlorophyll Fluorescence and P700 Absorbance in C3 and C4 Plants under Physiological Conditions and under Salt Stress. Int. J. Mol. Sci. 2022, 23, 3768. [Google Scholar] [CrossRef]
- Moustakas, M. Harnessing Chlorophyll Fluorescence for Phenotyping Analysis of Wild and Cultivated Tomato for High Photochemical Efficiency under Water Deficit for Climate Change Resilience. Climate 2021, 9, 154. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Q.; Lu, B.; Wang, M.; Li, Y.; Lu, B. Comparison of Photosynthetic Parameters Between Pyrus betulaefolia and Pyrus calleryana Under NaCl Stress. North. Hortic. 2019, 22, 97–104. [Google Scholar]
- Galicia-Campos, E.; Garcia-Villaraco, A.; Montero-Palmero, M.B.; Gutierrez-Manero, F.J.; Ramos-Solano, B. Bacillus G7 improves adaptation to salt stress in Olea europaea L. plantlets, enhancing water use efficiency and preventing oxidative stress. Sci. Rep. 2023, 13, 22507. [Google Scholar] [CrossRef]
- Lu, X.; Ma, L.; Zhang, C.; Yan, H.; Bao, J.; Gong, M.; Wang, W.; Li, S.; Ma, S.; Chen, B. Grapevine (Vitis vinifera) responses to salt stress and alkali stress: Transcriptional and metabolic profiling. Bmc Plant Biol. 2022, 22, 528. [Google Scholar] [CrossRef]
- Liu, B.; Wang, X.; Li, K.; Cai, Z. Spatially Resolved Metabolomics and Lipidomics Reveal Salinity and Drought-Tolerant Mechanisms of Cottonseeds. J. Agric. Food Chem. 2021, 69, 8028–8037. [Google Scholar] [CrossRef]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones Regulate Accumulation of Osmolytes Under Abiotic Stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Shi, X.; Wang, R.; Wu, X.; Wang, N.; Wang, Z. Osmotic Regulation and Antioxidant Capacity of Different Grapevine Rootstocks under Salt Stress. Acta Bot. Boreali-Occident. Sin. 2022, 42, 1880–1891. [Google Scholar]
- Oliveira, L.M.; Mendonca, V.; Moura, E.A.; Irineu, T.; Figueiredo, F.; Melo, M.F.; Celedonio, W.F.; Rego, A.; Mendonca, L.; Andrade, A. Salt stress and organic fertilization on the growth and biochemical metabolism of Hylocereus costaricensis (red pitaya) seedlings. Braz. J. Biol. 2022, 84, e258476. [Google Scholar] [CrossRef]
- Guo, X.; Ahmad, N.; Zhao, S.; Zhao, C.; Zhong, W.; Wang, X.; Li, G. Effect of Salt Stress on Growth and Physiological Properties of Asparagus Seedlings. Plants 2022, 11, 2836. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, J.; Yan, B.; Wei, Y.; Ge, S.; Li, J.; Han, Y.; Li, Z.; Zhao, C.; Xu, J. The Adjustment of Membrane Lipid Metabolism Pathways in Maize Roots Under Saline-Alkaline Stress. Front. Plant Sci. 2021, 12, 635327. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, X.; Zhang, R.; Yuan, H.; Wang, M.; Yang, H.; Ma, H.; Liu, D.; Jiang, C.; Liang, Z. Root Damage under Alkaline Stress Is Associated with Reactive Oxygen Species Accumulation in Rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 1580. [Google Scholar] [CrossRef]
- Li, H.; Guo, X. Influence of NaCl on activities of protective enzymes and MDA content in grape rootstock leaves. J. Fruit Sci. 2008, 25, 240–243. [Google Scholar]
- Bonnin, M.; Favreau, B.; Soriano, A.; Leonhardt, N.; Oustric, J.; Lourkisti, R.; Ollitrault, P.; Morillon, R.; Berti, L.; Santini, J. Insight into Physiological and Biochemical Determinants of Salt Stress Tolerance in Tetraploid Citrus. Antioxidants 2023, 12, 1640. [Google Scholar] [CrossRef]
- Wang, D.; Gao, Y.; Sun, S.; Lu, X.; Li, Q.; Li, L.; Wang, K.; Liu, J. Effects of Salt Stress on the Antioxidant Activity and Malondialdehyde, Solution Protein, Proline, and Chlorophyll Contents of Three Malus Species. Life 2022, 12, 1929. [Google Scholar] [CrossRef]
- Tirani, M.M.; Haghjou, M.M. Reactive oxygen species (ROS), Total antioxidant capacity (AOC) and Malondialdehyde (MDA) make a triangle in the evaluation of Zinc stress extension. J. Anim. Plant Sci. 2019, 29, 1100–1111. [Google Scholar]
- Tan, W.; Li, X.; Dong, Z.; Tan, M.; Tang, X. Principal component analysis on the alleviating effects of different concentrations of CaCl2 on ‘Tamina’ grape under NaCl stress. Plant Physiol. J. 2018, 54, 574–580. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, Y.; Geng, G.; Yang, R.; Yang, Z.; Yang, C.; Xu, R.; Zhang, Q.; Kakar, K.U.; Li, Z.; et al. Comparative Analysis of Physiological, Enzymatic, and Transcriptomic Responses Revealed Mechanisms of Salt Tolerance and Recovery in Tritipyrum. Front. Plant Sci. 2021, 12, 800081. [Google Scholar] [CrossRef] [PubMed]
- Gohari, G.; Zareei, E.; Rostami, H.; Panahirad, S.; Kulak, M.; Farhadi, H.; Amini, M.; Martinez-Ballesta, M.; Fotopoulos, V. Protective effects of cerium oxide nanoparticles in grapevine (Vitis vinifera L.) cv. Flame Seedless under salt stress conditions. Ecotox Environ. Safe 2021, 220, 112402. [Google Scholar] [CrossRef]
- Cai, Z.; Liu, X.; Chen, H.; Yang, R.; Chen, J.; Zou, L.; Wang, C.; Chen, J.; Tan, M.; Mei, Y.; et al. Variations in morphology, physiology, and multiple bioactive constituents of Lonicerae Japonicae Flos under salt stress. Sci. Rep. 2021, 11, 3939. [Google Scholar] [CrossRef]
- Lin, J.; Wang, Y.; Wang, G. Salt stress-induced programmed cell death in tobacco protoplasts is mediated by reactive oxygen species and mitochondrial permeability transition pore status. J. Plant Physiol. 2006, 163, 731–739. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.; Masud, A.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of Reactive Oxygen Species and Antioxidant Defense in Plants under Salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Li, R.; Zhou, L.; Jiang, L.; Wang, X. Morphological, Physiological and Biochemical Changes in the Grape Variety “Hotan Red” Caused by the Occurrence of Stress Under the Influence of Saline–Alkaline Growing Conditions. Horticulturae 2025, 11, 69. https://doi.org/10.3390/horticulturae11010069
Song Y, Li R, Zhou L, Jiang L, Wang X. Morphological, Physiological and Biochemical Changes in the Grape Variety “Hotan Red” Caused by the Occurrence of Stress Under the Influence of Saline–Alkaline Growing Conditions. Horticulturae. 2025; 11(1):69. https://doi.org/10.3390/horticulturae11010069
Chicago/Turabian StyleSong, Yan, Ruxue Li, Long Zhou, Lili Jiang, and Xiong Wang. 2025. "Morphological, Physiological and Biochemical Changes in the Grape Variety “Hotan Red” Caused by the Occurrence of Stress Under the Influence of Saline–Alkaline Growing Conditions" Horticulturae 11, no. 1: 69. https://doi.org/10.3390/horticulturae11010069
APA StyleSong, Y., Li, R., Zhou, L., Jiang, L., & Wang, X. (2025). Morphological, Physiological and Biochemical Changes in the Grape Variety “Hotan Red” Caused by the Occurrence of Stress Under the Influence of Saline–Alkaline Growing Conditions. Horticulturae, 11(1), 69. https://doi.org/10.3390/horticulturae11010069