
Overexpression of the EuSIP5 Gene to Improve Drought Resistance in Tobacco


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
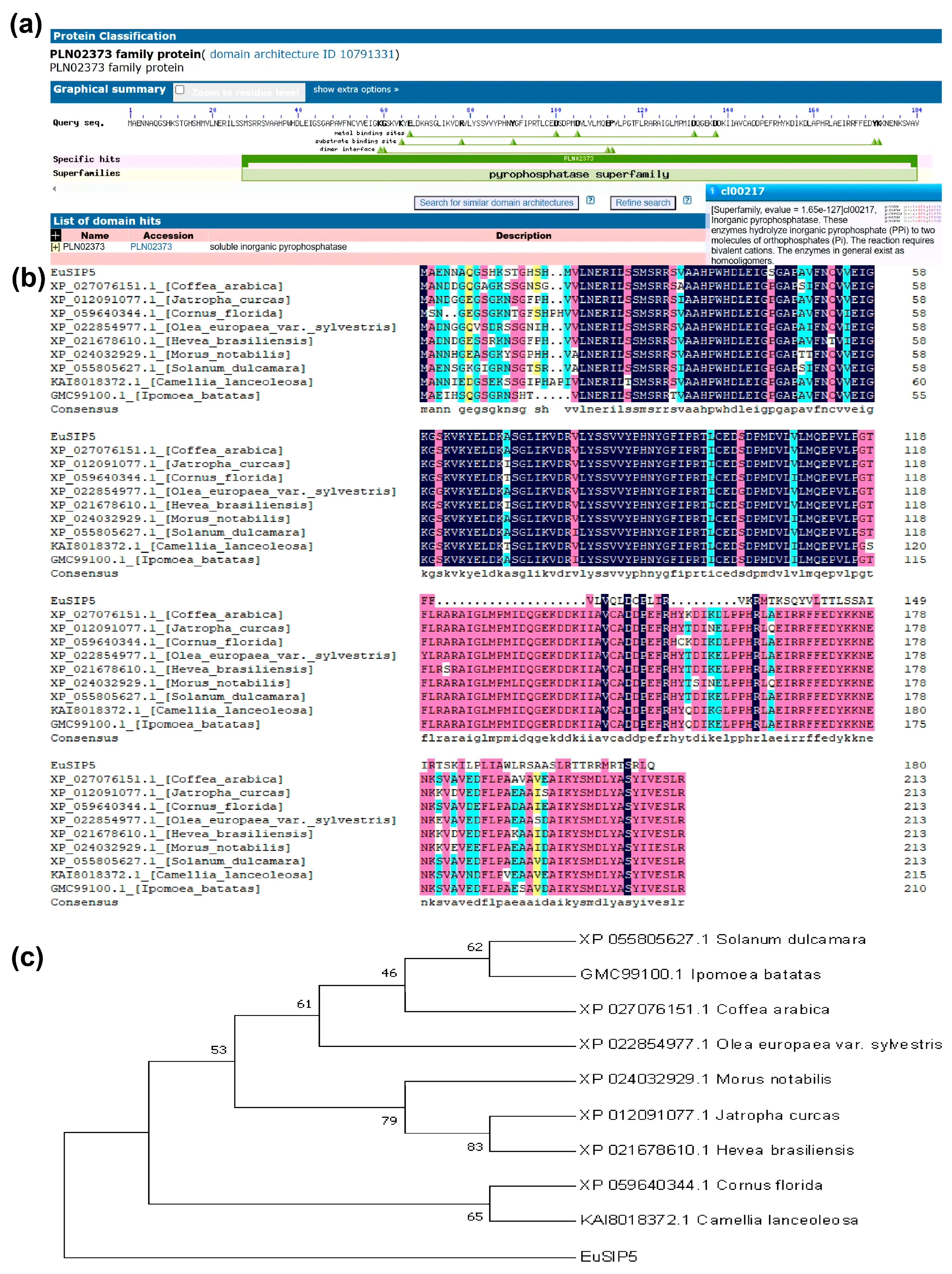
2.2.1. Process of Gene Cloning and Subsequent Bioinformatics Analysis
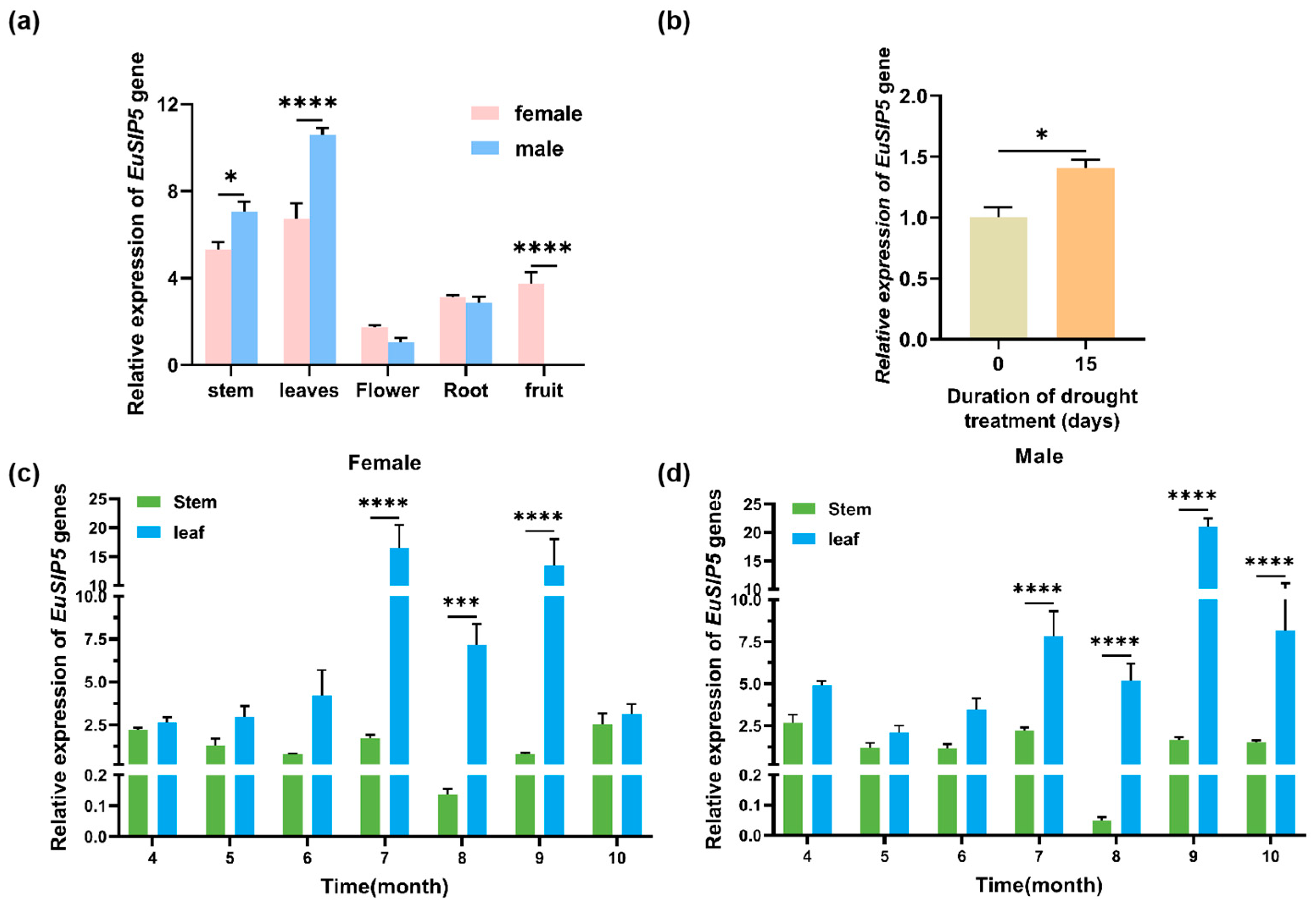
2.2.2. Expression Analysis of the EuSIP5 Gene in E. ulmoides
2.2.3. Vector Construction
2.2.4. Subcellular Localization
2.2.5. Genetic Transformation and Identification of N. tabacum L.
2.2.6. Quantitative Fluorescence Real-Time PCR (qRT-PCR)
2.2.7. Drought Stress Treatment of Transgenic N. tabacum L.
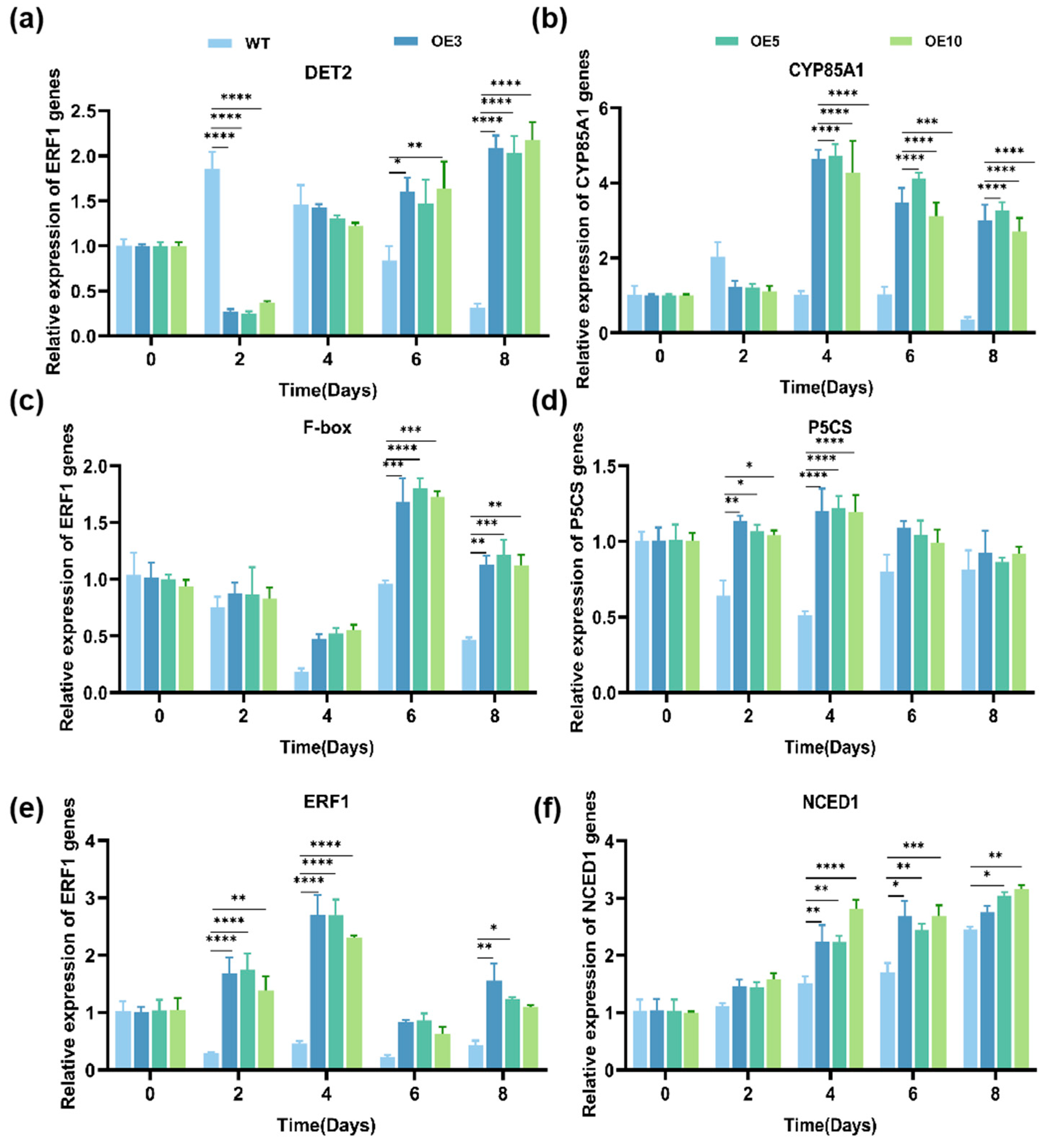
2.2.8. Expression Analysis of Genes Related to Drought Resistance in Transgenic N. tabacum L.
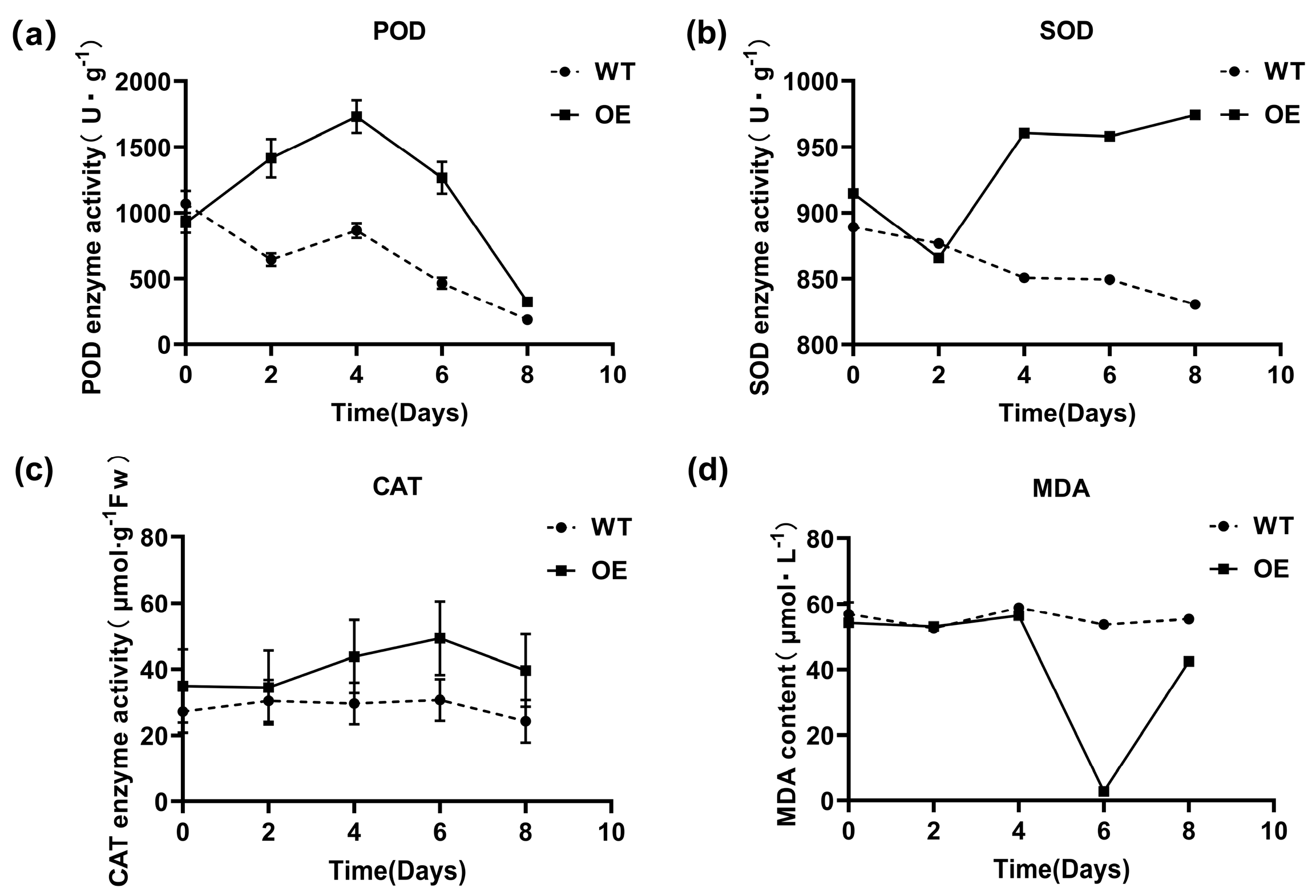
2.2.9. Determination of Protective Enzyme Activity
2.2.10. Statistical Analysis
3. Results
3.1. Cloning and Analysis of the EuSIP5 Gene
3.2. Spatiotemporal Expression Characteristics of the EuSIP5 Gene
3.3. Subcellular Localization Analysis of EuSIP5
3.4. Genetically Modified N. tabacum L. and Identification of Positive Transgenic Plants
3.5. Effect of the Trans-EuSIP5 Gene on Drought Tolerance in N. tabacum L.
3.6. Effect of Trans-EuSIP5 on the Activity of N. tabacum L. Protective Enzymes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Fujita, M. Drought Stress Responses in Plants, Oxidative Stress, and Antioxidant Defense. In Climate Change and Plant Abiotic Stress Tolerance; John Wiley & Sons: New York, NY, USA, 2013; pp. 209–250. [Google Scholar]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Razi, K.; Muneer, S. Drought stress-induced physiological mechanisms, signaling pathways and molecular response of chloroplasts in common vegetable crops. Crit. Rev. Biotechnol. 2021, 41, 669–691. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.X.; Shi, J.W.; Li, R.; An, Y.C.; Yang, B.L. Effects of extreme drought on plant species in Karst area of Guizhou Province, Southwest China. Chin. J. Appl. Ecol. 2011, 22, 1127–1134. [Google Scholar] [CrossRef]
- Ren, N.; Gong, W.W.; Zhao, Y.C.; Zhao, D.G.; Xu, Y.W. Innovation in sweet rice wine with high antioxidant activity: Eucommia ulmoides leaf sweet rice wine. Front. Nutr. 2023, 9, 1108843. [Google Scholar] [CrossRef]
- Wang, J.; Chen, X.; Huang, X.; Zhao, Y.; Zhao, D. Cloning and Function Identification of a Phytoene Desaturase Gene from Eucommia ulmoides. Phyton 2023, 92, 1377–1389. [Google Scholar] [CrossRef]
- Park, S.A.; Choi, M.S.; Kim, M.J.; Jung, U.J.; Kim, H.J.; Park, K.K.; Noh, H.J.; Park, H.M.; Park, Y.B.; Lee, J.S.; et al. Hypoglycemic and hypolipidemic action of Du-zhong (Eucommia ulmoides Oliver) leaves water extract in C57BL/KsJ-db/db mice. J. Ethnopharmacol. 2006, 107, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Chiba, A.; Murakami, M.; Sekihashi, K.; Tanaka, M.; Takahoko, M.; Moribayashi, S.; Kudou, C.; Hara, Y.; Nakazawa, Y.; et al. Antimutagenicity of Tochu tea (an aqueous extract of Eucommia ulmoides leaves): 2. Suppressing effect of Tochu tea on the urine mutagenicity after ingestion of raw fish and cooked beef. Mutat. Res. Genet. Toxicol. 1996, 371, 203–214. [Google Scholar] [CrossRef]
- Yen, G.C.; Hsieh, C.L. Reactive Oxygen Species Scavenging Activity of Du-zhong (Eucommia ulmoides Oliv.) and Its Active Compounds. J. Agric. Food Chem. 2000, 48, 3431–3436. [Google Scholar] [CrossRef]
- Dong, X.; Zhao, Y.C.; Ran, X.; Guo, L.X.; Zhao, D.G. Overexpression of a New Chitinase Gene EuCHIT2 Enhances Resistance to Erysiphe cichoracearum DC. in Tobacco Plants. Int. J. Mol. Sci. 2017, 18, 2361. [Google Scholar] [CrossRef]
- Li, Z.Y.; Li, B.; Zhao, Y.C.; Zhao, D.G. Cloning and characterization of the DIR1 promoter from Eucommia ulmoides Oliv and its response to hormonal and abiotic stress. Plant Cell Tissue Organ Cult. 2021, 146, 313–322. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, D.G. Cloning, Characterization, and Functional Analysis of EuTIL1, a Gene-Encoding Temperature-Induced Lipocalin in Eucommia ulmoides Oliv. Horticulturae 2023, 9, 950. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Liu, Y.Q.; Dong, X.; Liu, J.J.; Zhao, D.G. Identification of a novel laccase gene EuLAC1 and its potential resistance against Botrytis cinerea. Transgenic Res. 2022, 31, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.X.; Dong, X.; Huang, X.Z.; Zhao, D.G.; Zhao, Y.C.; Peng, L. Combined analysis of the transcriptome and proteome of Eucommia ulmoides Oliv. (Duzhong) in response to Fusarium oxysporum. Front. Chem. 2022, 10, 1053227. [Google Scholar] [CrossRef]
- Wang, L.; Wang, R.; Li, Y.; Zhao, Y.; Zhao, D. Overexpression of β-1,4-Glucanase Gene EuEG1 Improves Micrografting of Eucommia ulmoides. Phyton 2023, 92, 3063–3075. [Google Scholar] [CrossRef]
- Gloria, S.B.; Agustín, H.; Guillermo, L.L.; José Román, P.C.; Plácido, N.; Aurelio, S. Inorganic Pyrophosphatase Defects Lead to Cell Cycle Arrest and Autophagic Cell Death through NAD+ Depletion in Fermenting Yeast*. J. Biol. Chem. 2013, 288, 13082–13092. [Google Scholar] [CrossRef]
- Li, X.Y.; Zeng, R.Z.; Xiao, X.Z. Advances on the Research of Pyrophosphatase in Plants. Life Sci. Res. 2004, 8, 83–87. [Google Scholar] [CrossRef]
- Xian, J.H.; Zhang, M.P.; Sun, C.Y.; Wang, Y.F.; Wang, K.Y.; Chen, J.; Zhao, M.Z.; Wang, Y. Research Progress of Soluble Pyrophosphatase. Genom. Appl. Biol. 2019, 38, 4030–4035. [Google Scholar]
- Huang, H.; Patskovsky, Y.; Toro, R.; Farelli, J.D.; Pandya, C.; Almo, S.C.; Allen, K.N.; Dunaway-Mariano, D. Divergence of structure and function in the haloacid dehalogenase enzyme superfamily: Bacteroides thetaiotaomicron BT2127 is an inorganic pyrophosphatase. Biochemistry 2011, 50, 8937–8949. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, Y.R.; Wang, X.L.; Fang, T.; Zhen, S.H.; Lu, J.W.; Zhang, J.; Fu, J.J. Cloning and Expression Analysis of Soluble Inorganic Pyrophosphatase Family Genes in Maize (Zea mays L.). J. Plant Genet. Resour. 2021, 22, 455–465. [Google Scholar] [CrossRef]
- Zhu, J.H.; Xu, J.; Yu, X.H.; Chang, W.J.; Zhang, Z.L. Prokaryotic Expression for Three Soluble Inorganic Pyrophosphatase Genes from Hevea brasiliensis. Chin. J. Trop. Crops 2013, 34, 41–45. [Google Scholar]
- George, G.M.; van der Merwe, M.J.; Nunes-Nesi, A.; Bauer, R.; Fernie, A.R.; Kossmann, J.; Lloyd, J.R. Virus-induced gene silencing of plastidial soluble inorganic pyrophosphatase impairs essential leaf anabolic pathways and reduces drought stress tolerance in Nicotiana benthamiana. Plant Physiol. 2010, 154, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Song, Q.; Wei, H.; Wang, Y.; Lin, M.; Sun, K.; Zhang, Y.; Yang, J.; Li, C.; Luo, K.J.N.P. The AP2/ERF transcription factor PtoERF15 confers drought tolerance via JA-mediated signaling in Populus. New Phytol. 2023, 240, 1848–1867. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Hu, L.; Jiang, W. Understanding AP2/ERF Transcription Factor Responses and Tolerance to Various Abiotic Stresses in Plants: A Comprehensive Review. Int. J. Mol. Sci. 2024, 25, 893. [Google Scholar] [CrossRef]
- Wang, H.; Ni, D.; Shen, J.; Deng, S.; Xuan, H.; Wang, C.; Xu, J.; Zhou, L.; Guo, N.; Zhao, J.J.F.i.P.S. Genome-wide identification of the AP2/ERF gene family and functional analysis of GmAP2/ERF144 for drought tolerance in soybean. Front. Plant Sci. 2022, 13, 848766. [Google Scholar] [CrossRef]
- Yu, Y.; Yu, M.; Zhang, S.; Song, T.; Zhang, M.; Zhou, H.; Wang, Y.; Xiang, J.; Zhang, X. Transcriptomic Identification of Wheat AP2/ERF Transcription Factors and Functional Characterization of TaERF-6-3A in Response to Drought and Salinity Stresses. Int. J. Mol. Sci. 2022, 23, 3272. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yu, X.; Chen, L.; Zhao, G.; Li, S.; Zhou, H.; Dai, Y.; Sun, N.; Xie, Y.; Gao, J.; et al. Genome-wide identification and expression analysis of the NCED family in cotton (Gossypium hirsutum L.). PLoS ONE 2021, 16, e0246021. [Google Scholar] [CrossRef]
- Avico, E.H.; Acevedo, R.M.; Duarte, M.J.; Rodrigues Salvador, A.; Nunes-Nesi, A.; Ruiz, O.A.; Sansberro, P.A. Integrating Transcriptional, Metabolic, and Physiological Responses to Drought Stress in Ilex paraguariensis Roots. Plants 2023, 12, 2404. [Google Scholar] [CrossRef]
- Lee, S.-U.; Mun, B.-G.; Bae, E.-K.; Kim, J.-Y.; Kim, H.-H.; Shahid, M.; Choi, Y.-I.; Hussain, A.; Yun, B.-W. Drought Stress-Mediated Transcriptome Profile Reveals NCED as a Key Player Modulating Drought Tolerance in Populus davidiana. Front. Plant Sci. 2021, 12, 755539. [Google Scholar] [CrossRef]
- Chen, A.; Li, J.; Wang, H.; Zhao, P. Identification and Expression Profile of NCED Genes in Arachis hypogaea L. during Drought Stress. Int. J. Mol. Sci. 2024, 25, 5564. [Google Scholar] [CrossRef]
- Yang, D.; Ni, R.; Yang, S.; Pu, Y.; Qian, M.; Yang, Y.; Yang, Y. Functional Characterization of the Stipa purpurea P5CS Gene under Drought Stress Conditions. Int. J. Mol. Sci. 2021, 22, 9599. [Google Scholar] [CrossRef]
- Duan, F.M.; Ding, J.; Lee, D.S.; Lu, X.L.; Feng, Y.Q.; Song, W.W. Overexpression of SoCYP85A1, a spinach cytochrome p450 gene in transgenic tobacco enhances root development and drought stress tolerance. Front. Plant Sci. 2017, 8, 1909. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.F.; Ma, Y.Z.; Lu, H.D.; An, W.K.; Zhang, F.C. Effects of Spraying Brassinolide on Expression of Steroid 5-Alpha Reductase Gene (Gh DET2) in Cotton under Drought Stress. Genom. Appl. Biol. 2018, 37, 859–866. [Google Scholar] [CrossRef]
- An, J.; Li, Q.X.; Yang, J.J.; Zhang, G.Q.; Zhao, Z.X.; Wu, Y.Z.; Wang, Y.; Wang, W. Wheat F-box protein TaFBA1 positively regulates plant drought tolerance but negatively regulates stomatal closure. Front. Plant Sci. 2019, 10, 1242. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.H.; Zhao, Y.; Zhao, Y.; Feng, C.; Zhang, Y.H.; Wang, F.W.; Li, X.W.; Gao, H.T.; Liu, W.C.; Jing, Y. Soybean F-box-like protein GmFBL144 interacts with small heat shock protein and negatively regulates plant drought stress tolerance. Front. Plant Sci. 2022, 13, 823529. [Google Scholar] [CrossRef]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
- Thakur, K.; Garg, N. Oxidative Stress and Antioxidant Enzymes in Cereals Under Abiotic Stress. In Sustainable Remedies for Abiotic Stress in Cereals; Abdel Latef, A.A.H., Ed.; Springer Nature: Singapore, 2022; pp. 51–82. [Google Scholar]
- Türkan, İ.; Bor, M.; Özdemir, F.; Koca, H. Differential responses of lipid peroxidation and antioxidants in the leaves of drought-tolerant P. acutifolius Gray and drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress. Plant Sci. 2005, 168, 223–231. [Google Scholar] [CrossRef]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef]
- Navarro-De la Sancha, E.; Coello-Coutiño, M.P.; Valencia-Turcotte, L.G.; Hernández-Domínguez, E.E.; Trejo-Yepes, G.; Rodríguez-Sotres, R. Characterization of two soluble inorganic pyrophosphatases from Arabidopsis thaliana. Plant Sci. 2007, 172, 796–807. [Google Scholar] [CrossRef]
- Schulze, S.; Mant, A.; Kossmann, J.; Lloyd, J.R. Identification of an Arabidopsis inorganic pyrophosphatase capable of being imported into chloroplasts. FEBS Lett. 2004, 565, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Luna, F.M.; Navarro de la Sancha, E.; Valencia-Turcotte, L.G.; Vázquez-Santana, S.; Rodríguez-Sotres, R. Evidence for a non-overlapping subcellular localization of the family I isoforms of soluble inorganic pyrophosphatase in Arabidopsis thaliana. Plant Sci. 2016, 253, 229–242. [Google Scholar] [CrossRef]
- Tang, C.; Qiao, X.; Zhu, X.X.; Khan, W.; Wu, J.; Zhang, S.L. Expression and evolutionary analysis of soluble inorganic pyrophosphatase gene family in pear and four other Rosaceae species. Plant Syst. Evol. 2020, 306, 46. [Google Scholar] [CrossRef]
- Wang, J.; Kang, L.Y.; Liu, Z.B.; Lv, J.H.; Liu, Y.H.; Zou, X.X. Research Progress on the Impact of Drought on Plant. Hunan Agric. Sci. 2017, 7, 123–126+130. [Google Scholar] [CrossRef]
- Li, B.; Zeng, Q.; Zhao, D.; Zhao, D.G. Cloning and Function Analysis of EuERD16 Gene in Eucommia ulmoides. Genom. Appl. Biol. 2023, 42, 373–383. [Google Scholar] [CrossRef]
- Chen, S.Y. Membrane lipid peroxidation and plant adversity stresses. Bull. Bot. 1989, 212–215. [Google Scholar]
- Li, M.Q. Physiological and Molecular Regulatory Mechanisms of Soybean in Response to Drought. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2023. [Google Scholar]
- Wang, Q.M. Effects of Drought Stress on Protective Enzymes Activities and Membrane Lipid Peroxidation in Leaves of Soybean Seedlings. J. Agro-Environ. Sci. 2006, 918–921. [Google Scholar]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Zheng, A.Z.; Liu, C.P.; Shen, Z.G. Effect of Cadmium on MDA Content, POD and SOD Activities of Brassica Pekinensis and Brassica Chinensis. Hubei Agric. Sci. 2005, 1, 67–69. [Google Scholar]
- Song, X.G.; She, X.P. The Generation and the Role of Hydrogen Peroxide in Plant. J. Lianyungang Teach. Coll. 2010, 27, 99–103. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.; Chen, X.; Zhao, D.; Li, C. Overexpression of the EuSIP5 Gene to Improve Drought Resistance in Tobacco. Horticulturae 2024, 10, 1010. https://doi.org/10.3390/horticulturae10091010
Lin Y, Chen X, Zhao D, Li C. Overexpression of the EuSIP5 Gene to Improve Drought Resistance in Tobacco. Horticulturae. 2024; 10(9):1010. https://doi.org/10.3390/horticulturae10091010
Chicago/Turabian StyleLin, Yueling, Xi Chen, Degang Zhao, and Chao Li. 2024. "Overexpression of the EuSIP5 Gene to Improve Drought Resistance in Tobacco" Horticulturae 10, no. 9: 1010. https://doi.org/10.3390/horticulturae10091010
APA StyleLin, Y., Chen, X., Zhao, D., & Li, C. (2024). Overexpression of the EuSIP5 Gene to Improve Drought Resistance in Tobacco. Horticulturae, 10(9), 1010. https://doi.org/10.3390/horticulturae10091010