


Plant Morphological and Leaf Anatomical Traits in Coffea arabica L. Cultivars Cropped in Gorongosa Mountain, Mozambique

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Plant Material
2.2. Morpho-Agronomic Traits of the Plants
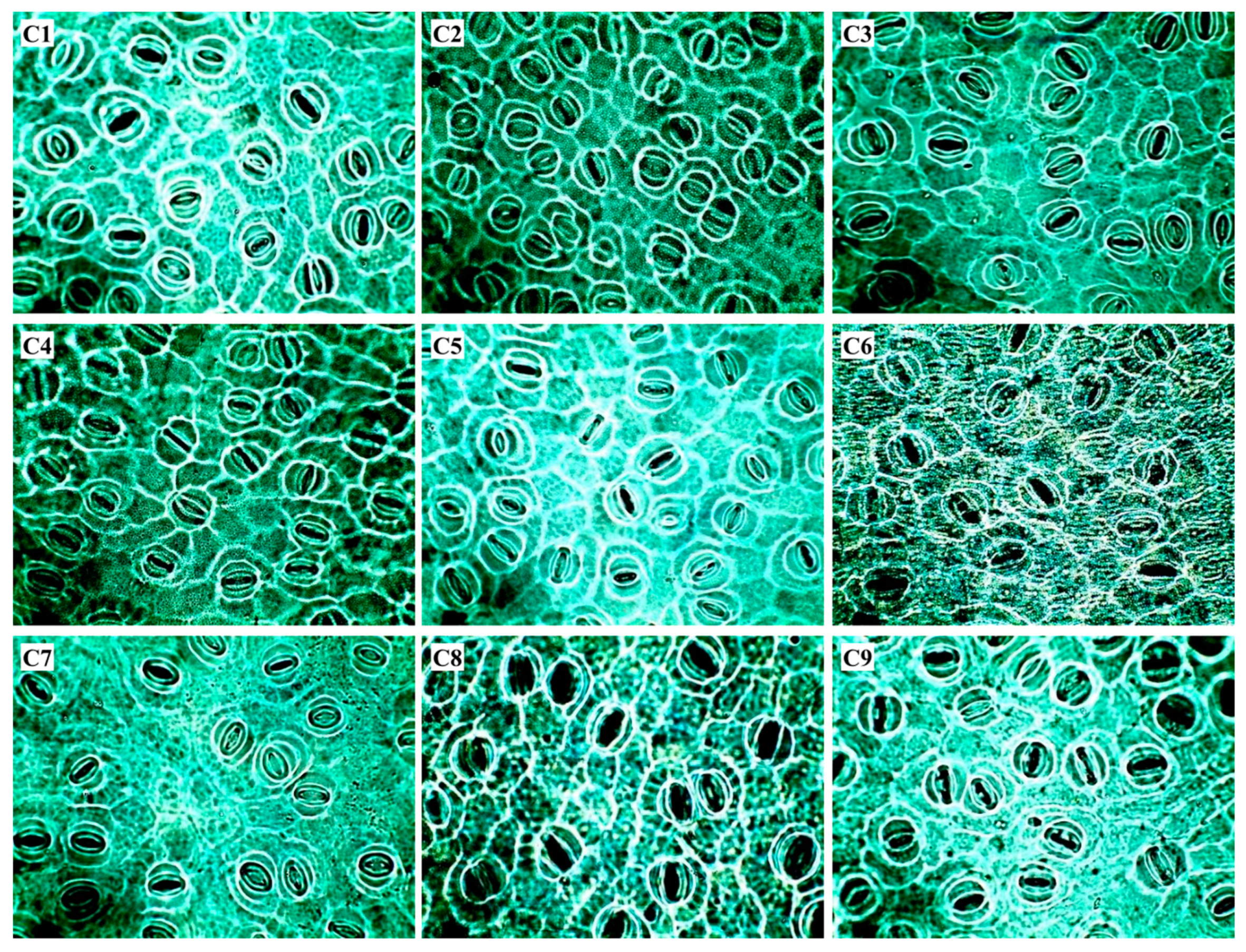
2.3. Leaf Anatomical Traits
2.4. Statistical Analysis
3. Results
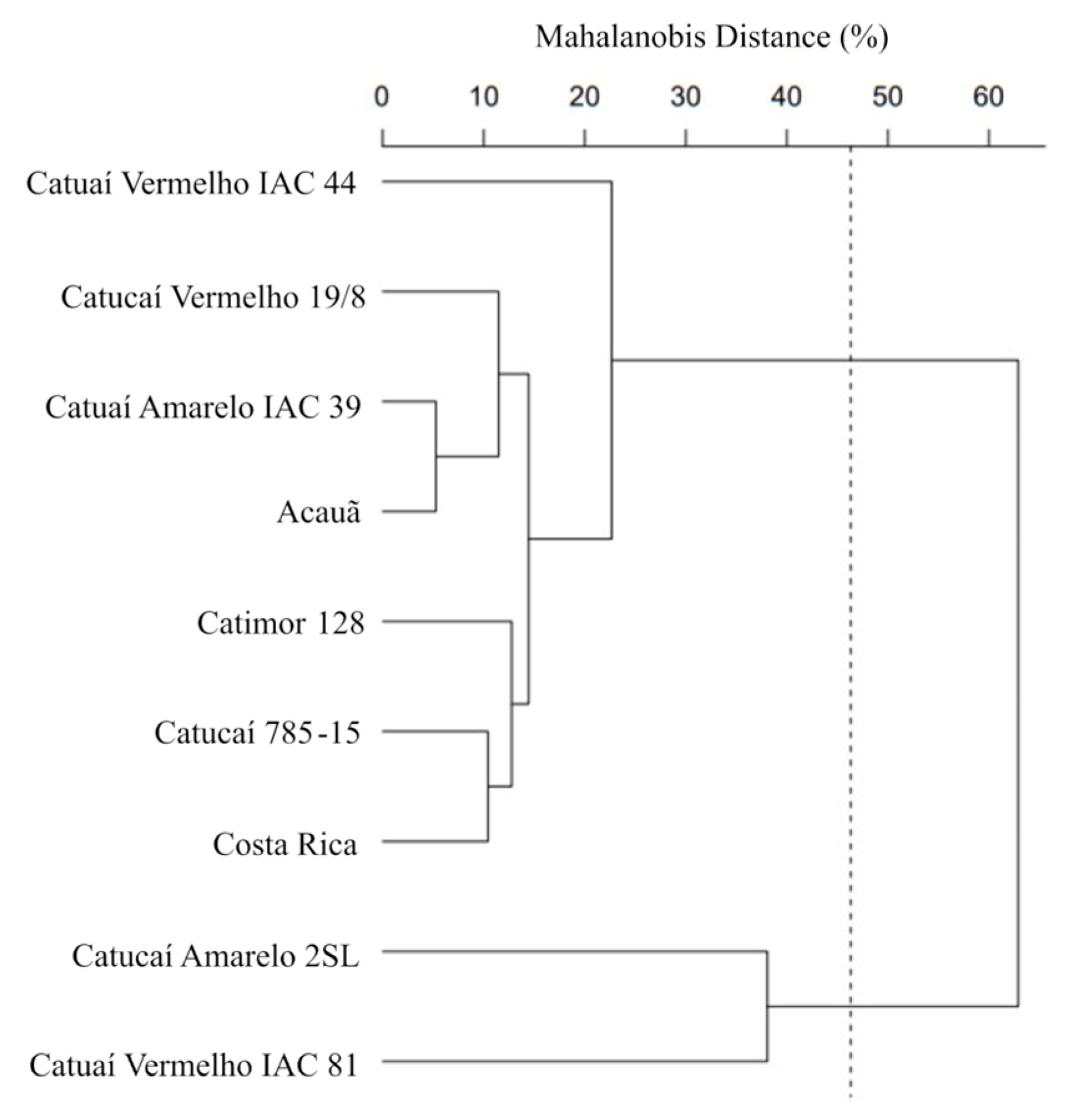
3.1. Morpho-Agronomic and Anatomical Dissimilarity between Genotypes and Genetic Contribution
3.2. Correlations among Morpho-Agronomic and Anatomical Leaf Traits in Nine Cultivars of C. arabica Cultivated in Gorongosa National Park, Mozambique
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, A.P.; Rakotonasolo, F. Six new species of coffee (Coffea) from northern Madagascar. Kew Bull. 2021, 76, 497–511. [Google Scholar] [CrossRef]
- Cassamo, C.T.; Mangueze, A.V.; Leitão, A.E.; Pais, I.P.; Moreira, R.; Campa, C.; Chiulele, R.; Reis, F.O.; Marques, I.; Scotti-Campos, P.; et al. Shade and altitude implications on the physical and chemical attributes of green coffee beans from Gorongosa Mountain, Mozambique. Agronomy 2022, 12, 2540. [Google Scholar] [CrossRef]
- Meyer, F.G. Notes on wild Coffea arabica from southwestern Ethiopia, with some historical considerations. Econ. Bot. 1965, 19, 136–151. [Google Scholar] [CrossRef]
- Mishra, M.K.; Tornincasa, P.; De-Nardi, B.; Asquini, E.; Dreos, R.; Terra, L.D.; Rathinavelu, R.; Rovelli, P.; Pallavicini, A.; Graziosi, G. Genome organization in coffee as revealed by EST PCR-RFLP, SNPs and SSR analysis. J. Crop Sci. Biotech. 2011, 14, 25–37. [Google Scholar] [CrossRef]
- Scalabrin, S.; Toniutti, L.; Di-Gaspero, G.; Scaglione, D.; Magris, G.; Vidotto, M.; Pinosio, S.; Cattonaro, F.; Magni, F.; Jurman, I.; et al. A single polyploidization event at the origin of the tetraploid genome of Coffea arabica is responsible for the extremely low genetic variation in wild and cultivated germplasm. Sci. Rep. 2020, 10, 4642. [Google Scholar] [CrossRef]
- Jingade, P.; Kumar, C.H.A.; Kumar, M.M. First report on genome size and ploidy determination of five indigenous coffee species using flow cytometry and stomatal analysis genetics & evolutionary biology—Original article. Braz. J. Bot. 2021, 44, 381–389. [Google Scholar]
- Davis, A.P.; Govaerts, R.; Bridson, D.M.; Stoffelen, P. An annotated taxonomic conspectus of the genus Coffea (Rubiaceae). Bot. J. Linn. Soc. 2006, 152, 465–512. [Google Scholar] [CrossRef]
- Lashermes, P.; Combes, M.C.; Roberto, J.; Trouslot, P.; D’Hont, A.; Antônio, F.; Carregador, A. Molecular characterization and origin of the Coffea arabica L. Mol. Genet. Genom. 1999, 261, 259–266. [Google Scholar] [CrossRef]
- Wright, S.I.; Kalisz, S.; Slotte, T. Evolutionary consequences of self-fertilization in plants. Proc. Royal Soc. B Biol. Sci. 2013, 280, 20130133. [Google Scholar] [CrossRef]
- Anthony, F.; Quiros, O.; Ropart, P.; Bertrand, B.; Lashermes, P. Detection by simple sequence repeat markers of introgression from Coffea canephora in Coffea arabica varieties. Plant Breed. 2002, 121, 542–544. [Google Scholar] [CrossRef]
- Anthony, F.; Combes, M.C.; Astorga, C.; Bertrand, B.; Graziosi, G.; Lashermes, P. The origin of cultivated Coffea arabica L. varieties revealed by AFLP and SSR markers. Theor. Appl. Genet. 2002, 104, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Charrier, A.; Berthaud, J. Botanical classification of coffee. In Coffee: Botany, Biochemistry and Production of Beans and Beverage; Clifford, M.N., Willson, K.C., Eds.; Springer: Boston, MA, USA, 1985; pp. 13–47. [Google Scholar]
- Aguiar, A.T.E.; Guerreiro-Filho, O.; Maluf, M.P.; Gallo, P.B.; Fazuoli, L.C. Characterization of Coffea arabica cultivars by minimum descriptors. Bragantia 2004, 63, 179–192. [Google Scholar] [CrossRef]
- Labouisse, J.P.; Bellachew, B.; Bertrand, B.; Kotecha, S. Current status of coffee (Coffea arabica L.) genetic resources in Ethiopia: Implications for conservation. Genet. Res. Crop Evol. 2008, 55, 1079–1093. [Google Scholar] [CrossRef]
- Akpertey, A.; Anim-Kwapong, E.; Ofori, A. Assessment of genetic diversity in Robusta coffee using morphological characters. Int. J. Fruit Sci. 2019, 19, 276–299. [Google Scholar] [CrossRef]
- Silva, L.O.E.; Schmidt, R.; Almeida, R.N.; Feitoza, R.B.B.; Cunha, M.; Partelli, F.L. Morpho-agronomic and leaf anatomical traits in Coffea canephora genotypes. Ciência Rural. 2023, 53, e20220005. [Google Scholar] [CrossRef]
- Oliveiro, G.F. Origin and Diverse Nature of Interspecific hybrid Coffee Cultivars; Documentos IAC, 119; Instituto Agronômico: Campinas, Brazil, 2021; p. 19. [Google Scholar]
- Hallé, F.; Oldeman, R.A.A.; Tomlinson, P.B. Tropical Trees and Forests-An Architectural Analysis; Springer: Berlin, Germany, 1978; 441p. [Google Scholar]
- Rakocevic, M. Coffee plant architecture. In Coffee—A Glimpse into the Future, Advances in Botanical Research; DaMatta, F.M., Ramalho, J.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2024. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Rodrigues, A.P.; Semedo, J.N.; Pais, I.P.; Martins, L.D.; Simoões-Costa, M.C.; Leitão, A.E.; Fortunato, A.S.; Batista-Santos, P.; Palos, I.M.; et al. Sustained photosynthetic performance of Coffea spp. under long-term enhanced [CO2]. PLoS ONE 2013, 8, e82712. [Google Scholar] [CrossRef]
- Lawson, T.; Blatt, M.R. Stomatal size, speed, and responsiveness impact on photosynthesis and water use efficiency. Plant Physiol. 2014, 164, 1556–1570. [Google Scholar] [CrossRef]
- Silva, P.C.; Junior, W.Q.R.; Ramos, M.L.G.; Rocha, O.C.; Veiga, A.D.; Silva, N.H.; Brasileiro, L.O.; Santana, C.C.; Soares, G.F.; Malaquias, J.V.; et al. Physiological changes of Arabica coffee under different intensities and durations of water stress in the Brazilian Cerrado. Plants 2022, 11, 2198. [Google Scholar] [CrossRef]
- Schlüter, U.; Muschak, M.; Berger, D.; Altmann, T. Photosynthetic performance of an Arabidopsis mutant with high stomatal density (sdd1-1) under different light regimes. J. Exp. Bot. 2003, 54, 867–874. [Google Scholar] [CrossRef]
- Blatt, M.R. Cell signaling and volume control in stomatal movements in plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 221–241. [Google Scholar] [CrossRef]
- Pautov, A.; Bauer, S.; Ivanova, O.; Ivanova, A.; Krylova, E. The mechanical advantage of subsidiary cells depends on the structure of the stomatal complex. Flora 2024, 311, 152457. [Google Scholar] [CrossRef]
- Lawson, T.; Oxborough, K.; Morison, J.I.L.; Baker, N.R. The photosynthesis response of guard cells to CO2, O2, light and drought stress in a variety of species is similar. J. Exp. Bot. 2003, 54, 1734–1752. [Google Scholar] [CrossRef] [PubMed]
- Lawson, T. Guard cell photosynthesis and stomatal function. New Phytol. 2009, 181, 13–34. [Google Scholar] [CrossRef]
- Bernado, W.P.; Rakocevic, M.; Santos, A.R.; Ruas, K.F.; Baroni, D.F.; Abraham, A.C.; Pireda, S.; Oliveira, D.S.; Cunha, M.; Ramalho, J.C.; et al. Biomass and leaf acclimations to ultraviolet solar radiation in juvenile plants of Coffea arabica and C. canephora. Plants 2021, 10, 640. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F.M.; Martins, S.C.V.; Ramalho, J.D.C. Ecophysiology of coffee growth and production in a context of climate changes. In Coffee—A Glimpse into the Future, Advances in Botanical Research; DaMatta, F.M., Ramalho, J.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2024. [Google Scholar]
- Bridson, D.M.; Verdcourt, B. Coffea in Flora of Tropical East Africa. In Flora of Tropical East Africa; A. A. Balkema: Rotterdam, The Netherlands, 1988; pp. 703–723. [Google Scholar]
- Alberto, N.J.; Ramalho, J.C.; Barros, A.I.R.; Viana, A.P.; Krohling, C.A.; Moiane, S.S.; Alberto, Z.; Rodrigues, W.P.; Partelli, F.L. Diversity in Coffea arabica cultivars in the mountains of Gorongosa National Park, Mozambique, regarding bean and leaf nutrient accumulation and physical fruit traits. Agronomy 2023, 13, 1162. [Google Scholar] [CrossRef]
- Tinley, K.L. Framework of the Gorongosa Ecosystem. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 1977. [Google Scholar]
- Bouley, P.; Poulos, M.; Branco, R.; Carter, N.H. Post-war recovery of the African lions in response to large-scale ecosystem restoration. Biol. Conserv. 2018, 227, 233–242. [Google Scholar] [CrossRef]
- Cassamo, C.T.; Draper, D.; Romeiras, M.M.; Marques, I.; Chiulele, R.; Rodrigues, M.; Stalmans, M.; Partelli, F.L.; Ribeiro-Barros, A.; Ramalho, J.C. Impact of climate changes in the suitable areas for Coffea arabica L. production in Mozambique: Agroforestry as an alternative management system to strengthen crop sustainability. Agric. Ecosyst. Environ. 2023, 346, 108341. [Google Scholar] [CrossRef]
- Ryan, C.M.; Williams, M. How does fire intensity and frequency affect Miombo woodland tree populations and biomass? Ecol. Appl. 2011, 21, 48–60. [Google Scholar] [CrossRef]
- Daskin, J.H.; Stalmans, M.; Pringle, R.M. Ecological legacies of civil war: 35-year increase in savanna tree cover following wholesale large-mammal declines. J. Ecol. 2016, 104, 79–89. [Google Scholar] [CrossRef]
- Köppen, W.P. Das Geographische System der Klimate; Gebrüdcr Borntraeger: Berlin, Germany, 1936. [Google Scholar]
- Aguiar, T.V.; Sant’Anna-Santos, B.F.; Azevedo, A.A.; Ferreira, R.S. Anati Quanti: Quantitative analysis software for plant anatomy studies. Planta Daninha 2007, 25, 649–659. [Google Scholar] [CrossRef]
- Castro, E.M.; Pereira, F.J.; Paiva, R. Vegetable Histology: Structure and Function of Vegetative Organs; Ufla: Lavras, Brazil, 2009; 234p. [Google Scholar]
- Sack, L.; Buckley, T.N. The developmental basis of stomatal density and flux. Plant Physiol. 2016, 171, 2358–2363. [Google Scholar] [CrossRef] [PubMed]
- Mojena, R. Hierarchical grouping methods and stopping rules: An evaluation. Comput. J. 1977, 20, 359–363. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing (R Version 4.3.0). 2022. Available online: https://www.r-project.org/ (accessed on 5 June 2023).
- Cruz, C.D. Programa Genes—Ampliado e integrado aos aplicativos R. Matlab e Selegen. Acta Sci.Agron. 2016, 38, 547–552. [Google Scholar] [CrossRef]
- Gomes, F.P. Curso de Estatística Experimental; Nobel: São Paulo, Brazil, 1985; 467p. [Google Scholar]
- Ferrão, R.G.; Cruz, C.D.; Ferreira, A.; Cecon, P.R.; Ferrão, M.A.G.; Fonseca, A.F.F.; Carneiro, P.C.S.; Silva, M.F. Genetic parameters in Conilon coffee. Pesqui. Agropecu. Bras. 2008, 43, 61–69. [Google Scholar] [CrossRef]
- Fritsche-Neto, R.; Vieira, R.A.; Scapim, C.A.; Miranda, G.V.; Rezende, L.M. Updating the ranking of the coefficients of variation from maize experiments. Acta Sci. Agron. 2012, 34, 99–101. [Google Scholar] [CrossRef]
- Couto, M.F.; Peternelli, L.A.; Barbosa, M.H.P. Classification of the coefficients of variation for sugarcane crops. Ciênc Rural. 2013, 43, 957–961. [Google Scholar] [CrossRef]
- Witkowski, E.T.F.; Lamont, B.B. Leaf specific mass confounds leaf density and thickness. Ecology 1991, 88, 486–493. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Oren, R.; Schulze, E.D.; Matyssek, R.; Zimmermann, R. Estimation of photosynthetic rate and annual carbon gain in conifers based on specific leaf weight and leaf biomass. Ecologia 1986, 70, 178–193. [Google Scholar]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From the tropics to the tundra: Global convergence in plant functioning. Proc. Nat. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef]
- Quero, J.L.; Villar, R.; Marañon, T.; Zamora, R. Interactions of the effects of drought and shade on seedlings of four Quercus species: Physiological and structural responses of the leaves. New Phytol. 2006, 170, 819–834. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F. The global spectrum of the leaf economy. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Pompelli, M.F.; Martins, S.C.V.; Celin, E.F.; Ventrella, M.C.; DaMatta, F.M. What is the influence of ordinary epidermal cells and stomata on the leaf plasticity of coffee plants grown under full-sun and shady conditions? Braz. J. Biol. 2010, 70, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Medrano, H. Drought-inhibition of Photosynthesis in C3 Plants: Stomatal and Non-stomatal Limitations Revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef]
- Resco, V.; Ewers, B.E.; Sun, W.; Huxman, T.E.; Weltzin, J.F.; Williams, D.G. Drought-induced hydraulic limitations constrain leaf gas exchange recovery after precipitation pulses in the C3 woody legume, Prosopis velutina. New Phytol. 2009, 181, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Radin, J.W.; Turcotte, E.L.; Percy, R.; Zeiger, E. High yields in advanced lines of Pima cotton are associated with higher stomatal conductance, reduced leaf area and lower leaf temperature. Physiol. Plant 1994, 92, 266–272. [Google Scholar] [CrossRef]
- Radin, J.W.; Lu, Z.; Percy, R.G.; Zeiger, E. Genetic variability for stomatal conductance in Pima cotton and its relation to improvements of heat adaptation. Proc. Nat. Acad. Sci. USA 1994, 91, 7217–7221. [Google Scholar] [CrossRef]
- Aliniaeifard, S.; van Meeteren, U. Natural variation in stomatal response to closing stimuli among Arabidopsis thaliana accessions after exposure to low VPD as a tool to recognize the mechanism of disturbed stomatal functioning. J. Exp. Bot. 2014, 65, 201412. [Google Scholar] [CrossRef]
- Giday, H.; Fanourakis, D.; Kjaer, K.H.; Fomsgaard, I.S.; Ottosen, C.O. Threshold response of stomatal closing ability to leaf abscisic acid concentration during growth. J. Exp. Bot. 2014, 65, 4361–4370. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Berry, J.A.; Beerling, D.J.; Franks, P.J. Stomata: Key players in the earth system, past and present. Curr. Opin. Plant Biol. 2010, 13, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Pieruschka, R.; Huber, G.; Berry, J.A. Control of transpiration by radiation. Proc. Nat. Acad. Sci. USA 2010, 107, 13372–13377. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, F. Any trait or trait-related allele can confer drought tolerance: Just design the right drought scenario. J. Exp. Bot. 2011, 63, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Fanourakis, D.; Giday, H.; Milla, R.; Pieruschka, R.; Kjaer, K.; Bolger, M.; Vasilevski, A.; Nunes-Nesi, A.N.; Fiorani, F.; Ottosen, C.O. Pore size regulates operating stomatal conductance, while stomatal densities drive the partitioning of conductance between leaf sides. Ann. Bot. 2015, 115, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Torii, K.U. Hormonal and environmental signals guiding stomatal development. BMC Biol. 2018, 16, 21. [Google Scholar] [CrossRef]
- Woodward, F.I.; Kelly, C.K. The influence of CO2 concentration on stomatal density. New Phytol. 1995, 131, 311–327. [Google Scholar] [CrossRef]
- Khan, P.S.S.V.; Kozai, T.; Nguyen, Q.T.; Kubota, C.; Dhawan, V. Growth and net photosynthetic rates of Eucalyptus tereticornis Smith under photomixotrophic and various photoautotrophic micropropagation conditions. Plant Cell Tissue Organ Cult. 2002, 71, 141–146. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Code | Cultivars | Genetic Origin | Maturation |
|---|---|---|---|
| 1 | Catuaí Vermelho IAC 44 | Bourbon 75% | Medium/Late |
| 2 | Catucaí Amarelo 2SL | Icatú Vermelho × Catuaí | Medium |
| 3 | Catuaí Amarelo IAC 39 | Bourbon 75% | Medium/Late |
| 4 | Catucaí 785-15 | Icatu Vermelho × Catuaí Vermelho | Early |
| 5 | Catucaí Vermelho 19/8 | Bourbon 75% | Early |
| 6 | Catuaí Vermelho IAC 81 | Bourbon 75% | Medium/Late |
| 7 | Acauã | Mundo Novo IAC 388-17 × Sarchimor IAC 1668 | Late |
| 8 | Catimor 128 | Undefined | Undefined |
| 9 | Costa Rica | Undefined | Undefined |
| Chemical Properties | |
|---|---|
| Organic matter—dag dm−3 | 5.50 |
| pH (water—Proportion 1:2.5) | 5.10 |
| Phosphorus (P)—mg dm−3 | 8.40 |
| Potassium (K)—mg dm−3 | 80.0 |
| Calcium (Ca)—cmolc·dm−3 | 1.20 |
| Magnesium (Mg)—cmolc·dm−3 | 0.40 |
| Aluminum (Al)—cmolc·dm−3 | 0.60 |
| H + Al—cmolc·dm−3 | 11.0 |
| Sum of bases (SB)—cmolc·dm−3 | 1.81 |
| Cation exchange capacity pH = 7—cmolc·dm−3 | 12.8 |
| Base saturation—% | 14.0 |
| Cation exchange capacity (K)—% | 2.00 |
| Cation exchange capacity (Ca)—% | 9.00 |
| Cation exchange capacity (Mg)—% | 3.00 |
| Cation exchange capacity (Al)—% | 4.70 |
| Cation exchange capacity (H + Al)—% | 86.0 |
| Natrium (Na)—mg dm−3 | 3.00 |
| Boron (B)—mg dm−3 | 0.20 |
| Zinc (Zn)—mg dm−3 | 1.40 |
| Manganese (Mn)—mg dm−3 | 30.6 |
| Cuprum (Cu)—mg dm−3 | 4.90 |
| Iron (Fe)—mg dm−3 | 28.0 |
| Cultivars | PH | CD | OD | NP | PD | PL | IL | LDM | LMA |
|---|---|---|---|---|---|---|---|---|---|
| cm | cm | cm | - | cm | cm | cm | mg | mg cm−2 | |
| Catuaí Vermelho IAC 44 | 89.4 a | 118.8 a | 3.45 a | 42.4 a | 0.95 b | 64.9 a | 2.80 a | 583.1 c | 19.8 b |
| Catucaí Amarelo 2SL | 103.7 a | 134.7 a | 3.50 a | 44.9 a | 0.91 b | 69.3 a | 3.11 a | 574.7 c | 20.5 b |
| Catuaí Amarelo IAC 39 | 91.8 a | 132.7 a | 3.59 a | 43.7 a | 0.92 b | 66.7 a | 2.81 a | 614.2 c | 22.4 b |
| Catucaí 785-15 | 92.0 a | 128.2 a | 3.65 a | 42.8 a | 1.00 a | 66.5 a | 2.81 a | 667.5 c | 23.7 a |
| Catucaí Vermelho 19/8 | 92.8 a | 129.0 a | 3.42 a | 42.4 a | 0.93 b | 67.8 a | 2.89 a | 609.3c | 21.0 b |
| Catuaí Vermelho IAC 81 | 97.8 a | 133.3 a | 3.78 a | 44.6 a | 0.90 b | 63.1 a | 2.68 a | 631.3 c | 23.2 a |
| Acauã | 94.2 a | 136.6 a | 3.56 a | 42.3 a | 1.00 a | 71.6 a | 2.90 a | 658.2 c | 23.5 a |
| Catimor 128 | 95.3 a | 128.8 a | 3.70 a | 38.0 a | 0.91 b | 65.7 a | 3.00 a | 777.6 a | 27.0 a |
| Costa Rica | 93.5 a | 120.6 a | 3.84 a | 44.2 a | 0.92 b | 60.2 a | 2.66 a | 695.8 b | 24.7 a |
| CV (%) | 8.01 | 7.40 | 7.09 | 8.82 | 5.78 | 8.17 | 8.11 | 8.23 | 9.95 |
| Cultivars | EC | SI | SD | SF |
|---|---|---|---|---|
| Cells mm−2 | % | Stomata mm−2 | - | |
| Catuaí Vermelho IAC 44 | 661.5 b | 33.5 a | 197.6 a | 1.76 a |
| Catucaí Amarelo 2SL | 822.0 a | 36.2 a | 234.0 a | 1.72 b |
| Catuaí Amarelo IAC 39 | 776.7 a | 36.0 a | 215.9 a | 1.68 b |
| Catucaí 785-15 | 731.4 a | 34.6 a | 212.3 a | 1.80 a |
| Catucaí Vermelho 19/8 | 761.3 a | 33.5 a | 228.5 a | 1.76 a |
| Catuaí Vermelho IAC 81 | 742.7 a | 38.7 a | 192.4 a | 1.70 b |
| Acauã | 766.2 a | 36.1 a | 212.7 a | 1.68 b |
| Catimor 128 | 765.5 a | 35.4 a | 216.7 a | 1.72 b |
| Costa Rica | 769.4 a | 37.1 a | 207.7 a | 1.80 a |
| CV (%) | 6.05 | 8.25 | 11.7 | 3.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alberto, N.J.; Ferreira, A.; Ribeiro-Barros, A.I.; Aoyama, E.M.; Silva, L.O.E.; Rakocevic, M.; Ramalho, J.C.; Partelli, F.L. Plant Morphological and Leaf Anatomical Traits in Coffea arabica L. Cultivars Cropped in Gorongosa Mountain, Mozambique. Horticulturae 2024, 10, 1002. https://doi.org/10.3390/horticulturae10091002
Alberto NJ, Ferreira A, Ribeiro-Barros AI, Aoyama EM, Silva LOE, Rakocevic M, Ramalho JC, Partelli FL. Plant Morphological and Leaf Anatomical Traits in Coffea arabica L. Cultivars Cropped in Gorongosa Mountain, Mozambique. Horticulturae. 2024; 10(9):1002. https://doi.org/10.3390/horticulturae10091002
Chicago/Turabian StyleAlberto, Niquisse José, Adésio Ferreira, Ana I. Ribeiro-Barros, Elisa Mitsuko Aoyama, Larícia Olária Emerick Silva, Miroslava Rakocevic, José Cochicho Ramalho, and Fábio Luiz Partelli. 2024. "Plant Morphological and Leaf Anatomical Traits in Coffea arabica L. Cultivars Cropped in Gorongosa Mountain, Mozambique" Horticulturae 10, no. 9: 1002. https://doi.org/10.3390/horticulturae10091002
APA StyleAlberto, N. J., Ferreira, A., Ribeiro-Barros, A. I., Aoyama, E. M., Silva, L. O. E., Rakocevic, M., Ramalho, J. C., & Partelli, F. L. (2024). Plant Morphological and Leaf Anatomical Traits in Coffea arabica L. Cultivars Cropped in Gorongosa Mountain, Mozambique. Horticulturae, 10(9), 1002. https://doi.org/10.3390/horticulturae10091002