
Application of AMF Alleviates Growth and Physiological Characteristics of Impatiens walleriana under Sub-Low Temperature


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Treatment Design
2.2.1. Hole Tray and Strain Preparation
2.2.2. Seed Pretreatment
2.2.3. Treatment Details
- (1)
- The control treatment (CK): normal temperature treatment without AMF inoculation;
- (2)
- AMF-50 treatment: 50 g AMF inoculated at normal temperature;
- (3)
- AMF-100 treatment: 100 g AMF inoculated at normal temperature;
- (4)
- CK+L treatment: sub-low temperature treatment without AMF inoculation (12 °C/8 °C (day/night));
- (5)
- AMF-50+L treatment: 50 g AMF inoculated with sub-low temperature treatment (12 °C/8 °C (day/night));
- (6)
- AMF-100+L treatment: 100 g AMF inoculated with sub-low temperature treatment (12 °C/8 °C (day/night)).
2.3. The Effect of Mycorrhizal Infection
2.4. Determination of Growth Indicators
2.5. Determination of Chlorophyll Fluorescence
2.6. Determination of Plasma Membrane Permeability of Leaves and Roots
2.7. Determination of O2− and H2O2 Content in Leaves
2.8. Determination of Damage Degree of Leaf Membrane Lipid Peroxidation
2.9. Data Statistics and Analysis
3. Results
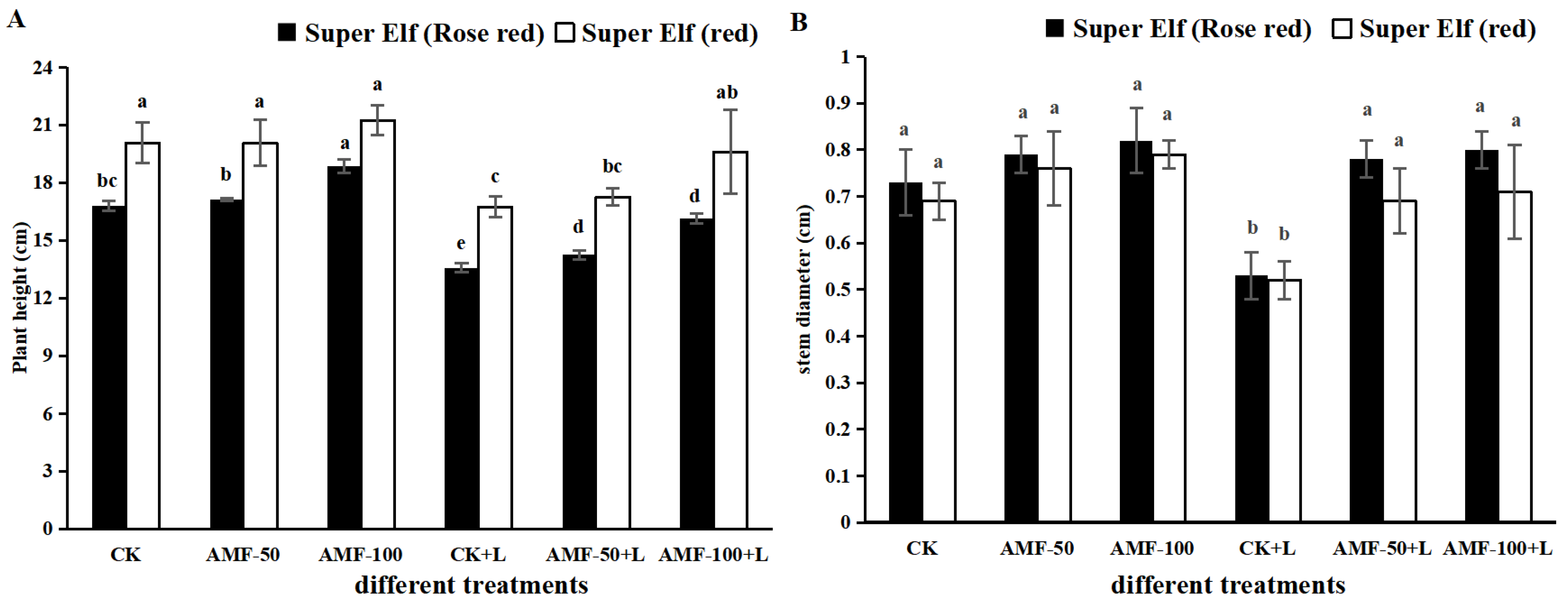
3.1. Effects of AMF on Growth Indices of Impatiens walleriana under Sub-Low Temperature Stress
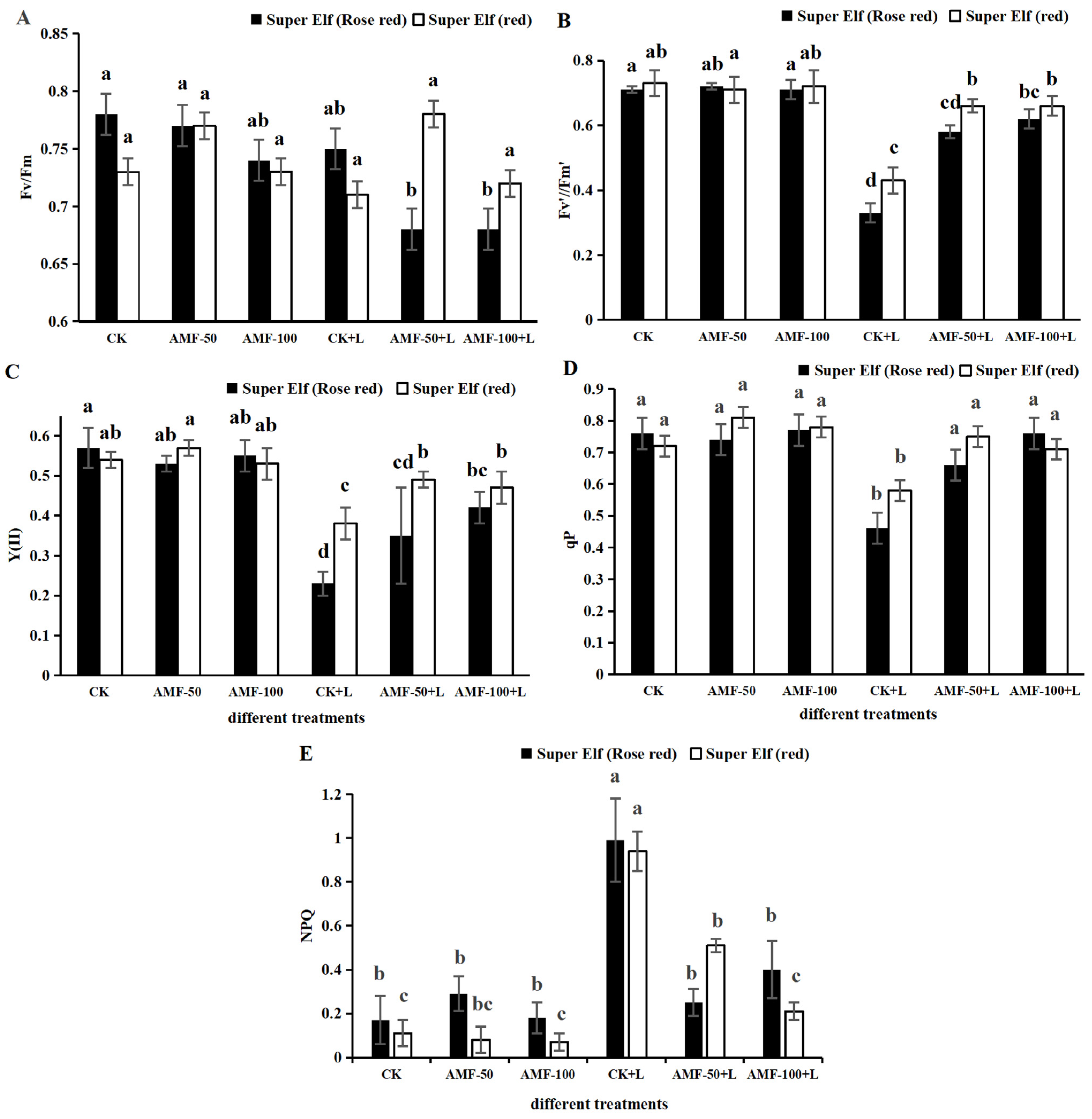
3.2. Effect of AMF on Chlorophyll Fluorescence of Impatiens walleriana Leaves under Sub-Low Temperature Stress
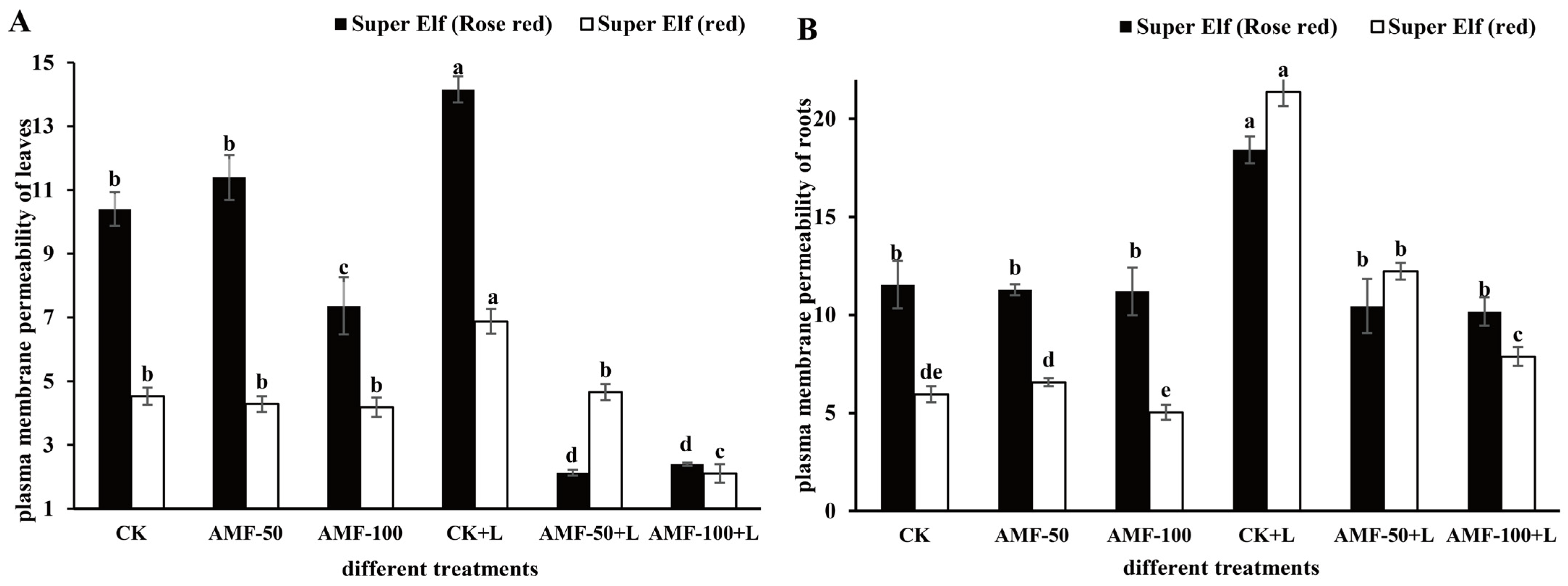
3.3. Effects of AMF on Plasma Membrane Permeability of Impatiens walleriana Leaves and Roots under Sub-Low Temperature Stress
3.4. Effects of AMF on O2− and H2O2 Content of Impatiens walleriana Leaves under Sub-Low Temperature Stress
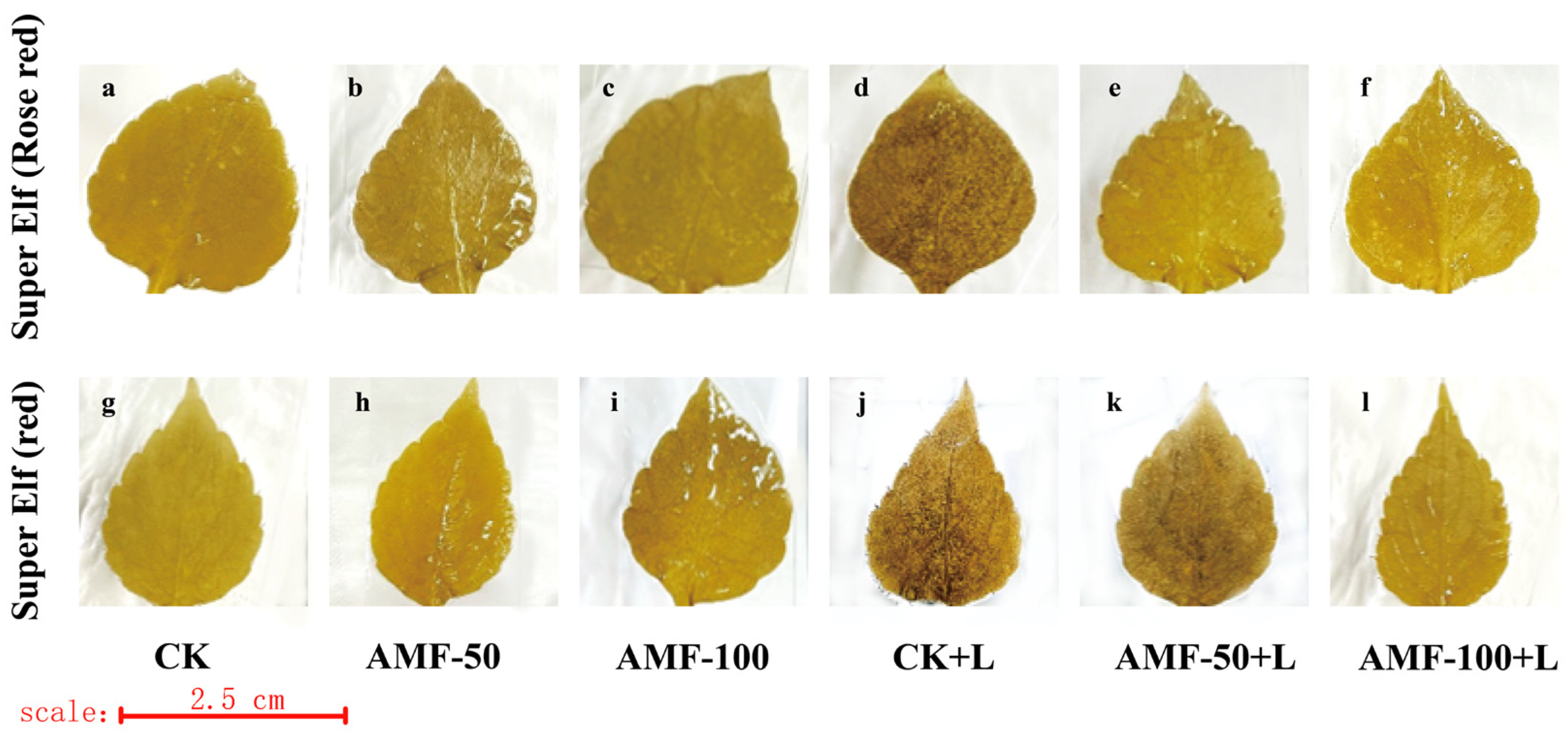
3.5. Effect of AMF on the Degree of Membrane Lipid Peroxidation of Impatiens walleriana Leaves under Sub-Low Temperature Stress
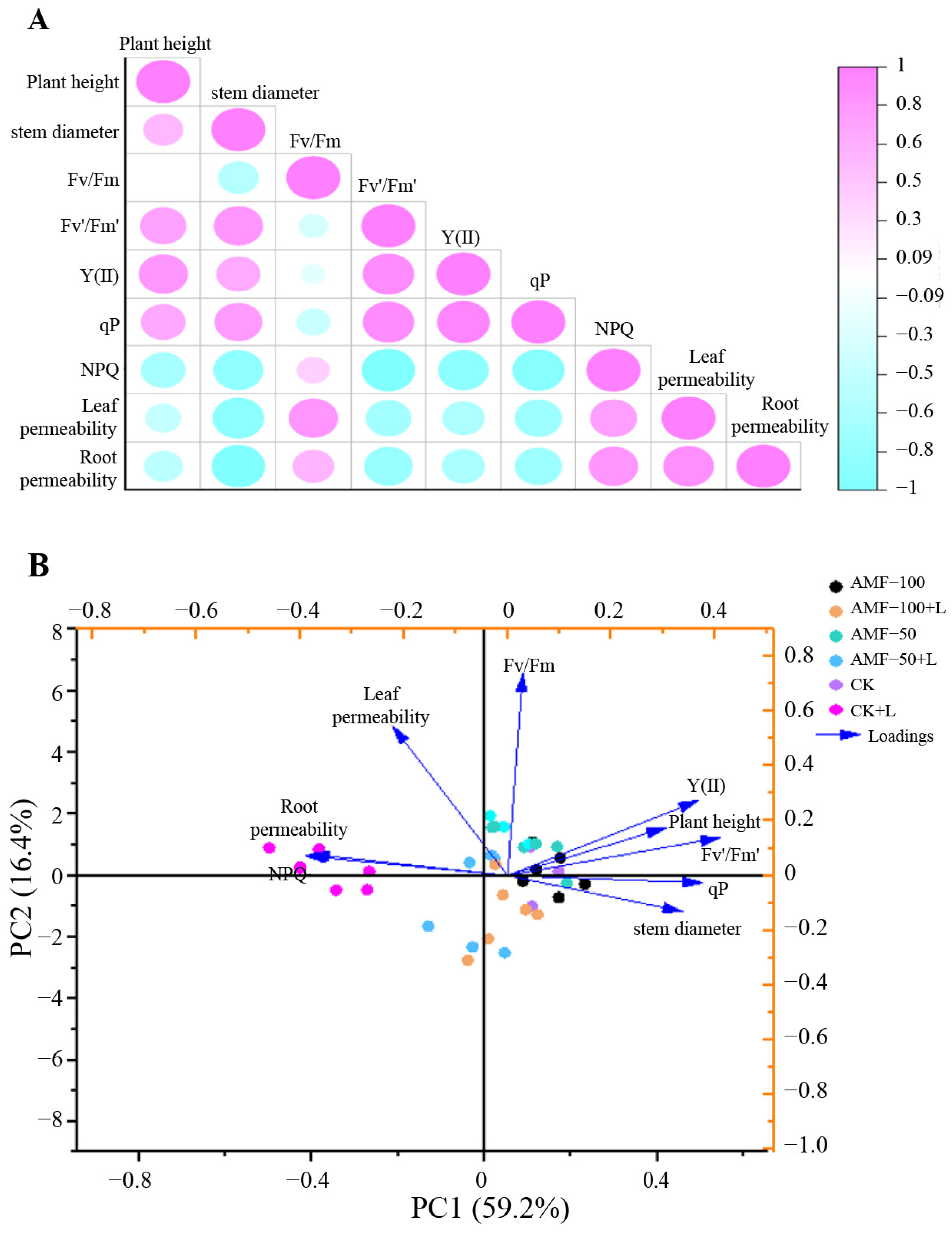
3.6. Principal Component and Correlation Analysis
4. Discussion
4.1. Effects of AMF on Plant Growth under Sub-Low Temperature Stress
4.2. Effects of AMF on Plant Photosynthesis under Sub-Low Temperature Stress
4.3. Effects of AMF on Plant Antioxidant System under Sub-Low Temperature Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhou, M.H.; Jiang, Y.; He, X.F.; Zhou, S.F. African Impatiens production and cultivation techniques. Agric. Sci. Technol. Inf. 2011, 4, 21–22. [Google Scholar] [CrossRef]
- Li, Y.; Guo, W.C.; Liu, J.H. The chemical constituents and pharmacological effects of Impatiens balsamina and its edible and ornamental. Spec. Econ. Anim. Plants 2015, 9, 33–36. [Google Scholar] [CrossRef]
- Hu, X.L.; Han, Z.X.; Liu, Y.F.; Zhao, H.; Li, Y.L.; Mao, N.W. Study on Antioxidative Activity of Extracts from Flowers of Impatiens balsamina L. Food Sci. 2007, 2, 48–50. [Google Scholar] [CrossRef]
- Shi, W.L. Research Status of Impatiens Germplasm Resources. Nanfang Agric. Mach. 2024, 55, 159–161. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Devireddy, A.R.; Tschaplinski, T.J.; Tuskan, G.A.; Muchero, W.; Chen, J.G. Role of Reactive Oxygen Species and Hormones in Plant Responses to Temperature Changes. Int. J. Mol. Sci. 2021, 22, 8843–8865. [Google Scholar] [CrossRef] [PubMed]
- Romero, F.R.; Gladon, R.J.; Taber, H.G. Effect of excessive calcium applications on growth and postharvest performance of bedding-plant impatiens. J. Plant Nutr. 2007, 30, 1639–1649. [Google Scholar] [CrossRef]
- Nelson, P.V.; Song, C.Y.; Huang, J.S.; Niedziela, C.E.; Swallow, W.H. Relative Effects of Fertilizer Nitrogen Form and Phosphate Level on Control of Bedding Plant Seedling Growth. Hortscience 2012, 47, 249–253. [Google Scholar] [CrossRef]
- Ye, L.F. Cultivation Techniques of African Impatiens. Agric. Inf. China 2013, 11, 81–82. [Google Scholar]
- Li, L.J.; Yang, B.L.; Zhao, X.P.; Wang, P.; Lyu, D.; Qin, S.J. Auxin Participates in the Regulation of the Antioxidant System in Malus baccata Borkh. Roots under Sub-Low Temperature by Exogenous Sucrose Application. Horticulturae 2023, 9, 297–315. [Google Scholar] [CrossRef]
- Liu, L.L.; Song, H.; Shi, K.J.; Liu, B.; Zhang, Y.; Tang, L.; Cao, W.X.; Zhu, Y. Response of wheat grain quality to low temperature during jointing and booting stages-On the importance of considering canopy temperature. Agric. For. Meteorol. 2019, 278, 107658. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Ma, K.F.; Li, Q.W. Effects of Low-Temperature Accumulation on Flowering of Prunus mume. Horticulturae 2023, 9, 628–637. [Google Scholar] [CrossRef]
- Gao, H.; Wu, F.Z. Physiological and transcriptomic analysis of tomato in response to sub-optimal temperature stress. Plant Signal. Behav. 2024, 19, e2332018. [Google Scholar] [CrossRef]
- Li, H.; Wang, F.; Chen, X.J.; Shi, K.; Xia, X.J.; Considine, M.J.; Yu, J.Q.; Zhou, Y.H. The sub/supra-optimal temperature-induced inhibition of photosynthesis and oxidative damage in cucumber leaves are alleviated by grafting onto figleaf gourd/luffa rootstocks. Physiol. Plant 2014, 152, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Bidabadi, S.S.; Mehralian, M. Arbuscular Mycorrhizal Fungi Inoculation to Enhance Chilling Stress Tolerance of Watermelon. Gesunde Pflanz. 2020, 72, 171–179. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, M.M.; Wang, Z.F.; Li, J.R.; Liu, K.; Huang, D. The role of arbuscular mycorrhizal symbiosis in plant abiotic stress. Front. Microbiol. 2024, 14, 1323881. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.S.; Zhang, Q.; Pan, J.B.; Jiang, S.J.; Liu, Y.J.; Bahadur, A.; Peng, Z.L.; Yang, Y.; Feng, H.Y. Effect of arbuscular mycorrhizal fungi on soil enzyme activity is coupled with increased plant biomass. Eur. J. Soil. Sci. 2020, 71, 84–92. [Google Scholar] [CrossRef]
- Dias, T.; Correia, P.; Carvalho, L.; Melo, J.; Varennes, A.; Cruz, C. Arbuscular mycorrhizal fungal species differ in their capacity to overrule the soil’s legacy from maize monocropping. Appl. Soil. Ecol. 2018, 125, 177–183. [Google Scholar] [CrossRef]
- Liu, A.R.; Chen, S.C.; Wang, M.M.; Liu, D.L.; Chang, R.; Wang, Z.H.; Lin, X.M.; Bai, B.; Ahammed, G.J. Arbuscular Mycorrhizal Fungus Alleviates Chilling Stress by Boosting Redox Poise and Antioxidant Potential of Tomato Seedlings. J. Plant Res. 2016, 35, 109–120. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S.; Kataria, S.; Alamri, S.A.; Siddiqui, M.H.; Rastogi, A. Inoculation with Arbuscular Mycorrhizal Fungi Alleviates the Adverse Effects of High Temperature in Soybean. Plants 2022, 11, 2210–2222. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.Q.; Xu, L.L.; Chen, X.Y.; Yang, P.; Han, S.M. Effects of arbuscular mycorrhizal fungi on physiological characteristics of Impatiens balsamina under cadmium stress. Jiangsu Agric. Sci. 2019, 47, 186–188. [Google Scholar] [CrossRef]
- Liu, A.R.; Chen, S.C.; Chang, R.; Liu, D.L.; Chen, H.R.; Ahammed, G.J.; Lin, X.M.; He, C.X. Arbuscular mycorrhizae improve low temperature tolerance in cucumber via alterations in H2O2 accumulation and ATPase activity. J. Plant Res. 2014, 127, 775–785. [Google Scholar] [CrossRef]
- Hajiboland, R.; Joudmand, A.; Aliasgharzad, N.; Tolrá, R.; Poschenrieder, C. Arbuscular mycorrhizal fungi alleviate low-temperature stress and increase freezing resistance as a substitute for acclimation treatment in barley. Crop Pasture Sci. 2019, 70, 218–233. [Google Scholar] [CrossRef]
- Li, W.; Zhai, Y.L.; Xing, H.S.; Xing, L.J.; Guo, S.X. Arbuscular mycorrhizal fungi promote photosynthesis in Antirrhinum majus L. under low-temperature and weak-light conditions. Not. Bot. Horti Agrobot. Cluj. Napoca 2023, 51, 13012. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Shao, X.F.; Gong, Y.F.; Zhu, Y.; Wang, H.F.; Zhang, X.L.; Yu, D.D.; Yu, F.; Qiu, Z.Y.; Lu, H. The metabolism of soluble carbohydrates related to chilling injury in peach fruit exposed to cold stress. Postharvest Biol. Technol. 2013, 86, 53–61. [Google Scholar] [CrossRef]
- Xu, J.; Zhu, Y.Y.; Ge, Q.; Li, Y.L.; Sun, J.H.; Zhang, Y.; Liu, X.J. Comparative physiological responses of Solanum nigrum and Solanum torvum to cadmium stress. New Phytol. 2012, 196, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Pompella, A.; Maellaro, E.; Casini, A.F.; Comporti, M. Histochemical detection of lipid peroxidation in the liver of bromobenzene-poisoned mice. Am. J. Pathol. 1987, 129, 295–301. [Google Scholar] [CrossRef]
- Mathur, S.; Sharma, M.P.; Jajoo, A. Improved photosynthetic efficacy of maize (Zea mays) plants with arbuscular mycorrhizal fungi (AMF) under high temperature stress. J. Photochem. Photobiol. B 2018, 180, 149–154. [Google Scholar] [CrossRef]
- Lee, Y.; Krishnamoorthy, R.; Selvakumar, G.; Kim, K.; Sa, T. Alleviation of salt stress in maize plant by co-inoculation of arbuscular mycorrhizal fungi and Methylobacterium oryzae CBMB20. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 533–540. [Google Scholar] [CrossRef]
- Chen, X.Y.; Song, F.B.; Liu, F.L.; Tian, C.J.; Liu, S.Q.; Xu, H.W.; Zhu, X. Effect of Different Arbuscular Mycorrhizal Fungi on Growth and Physiology of Maize at Ambient and Low Temperature Regimes. Sci. World J. 2014, 1, 956141. [Google Scholar] [CrossRef]
- Yan, P.; Li, G.; Sun, H.Q.; Zhang, Z.H.; Yang, R.Y.; Sun, J.N. Can arbuscular mycorrhizal fungi and biochar enhance plant resistance to low-temperature stress? Agron. J. 2021, 113, 1457–1466. [Google Scholar] [CrossRef]
- Chen, S.C.; Jin, W.J.; Liu, A.R.; Zhang, S.J.; Liu, D.L.; Wang, F.H.; Lin, X.M.; He, C.X. Arbuscular mycorrhizal fungi (AMF) increase growth and secondary metabolism in cucumber subjected to low temperature stress. Sci. Hortic. 2013, 160, 222–229. [Google Scholar] [CrossRef]
- Liu, Y.F.; Han, X.R.; Zhan, X.M.; Yang, J.F.; Wang, Y.Z.; Song, Q.B.; Chen, X. Regulation of Calcium on Peanut Photosynthesis Under Low Night Temperature Stress. J. Integr. Agric. 2013, 12, 2172–2178. [Google Scholar] [CrossRef]
- Machino, S.; Nagano, S.; Hikosaka, K. The latitudinal and altitudinal variations in the biochemical mechanisms of temperature dependence of photosynthesis within Fallopia japonica. Environ. Exp. Bot. 2021, 181, 104248. [Google Scholar] [CrossRef]
- He, B.; Chen, Y.; Zhang, H.; Xia, C.Y.; Zhang, Q.; Li, W. The effect of colored plastic films on the photosynthetic characteristics and content of active ingredients of Dysosma versipellis. Hortic. Environ. Biotechnol. 2018, 59, 519–528. [Google Scholar] [CrossRef]
- Ghosh, D.; Mohapatra, S.; Dogra, V. Improving photosynthetic efficiency by modulating non-photochemical quenching. Trends Plant Sci. 2023, 28, 264–266. [Google Scholar] [CrossRef]
- Feng, Z.; Liang, F.; Zheng, C.S.; Shu, H.R.; Sun, X.Z.; Yong-kweon, Y. Effects of Acetylsalicylic Acid and Calcium Chloride on Photosynthetic Apparatus and Reactive Oxygen-Scavenging Enzymes in Chrysanthemum Under Low Temperature Stress with Low Light. Agric. Sci. China 2010, 9, 1777–1786. [Google Scholar] [CrossRef]
- Li, S.X.; Yang, W.Y.; Guo, J.H.; Li, X.N.; Lin, J.X.; Zhu, X.C. Changes in photosynthesis and respiratory metabolism of maize seedlings growing under low temperature stress may be regulated by arbuscular mycorrhizal fungi. Plant Physiol. Biochem. 2020, 154, 1–10. [Google Scholar] [CrossRef]
- Sagi, M.; Fluhr, R. Production of reactive oxygen species by plant NADPH oxidases. Plant Physiol. 2006, 141, 336–340. [Google Scholar] [CrossRef]
- Cerqueira, J.A.; Andrade, M.T.; Rafael, D.D.; Zhu, F.; Martins, S.V.C.; Nunes-Nesi, A.; Benedito, V.; Fernie, A.R.; Zsögön, A. Anthocyanins and reactive oxygen species: A team of rivals regulating plant development? Plant Mol. Biol. 2023, 112, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Zhang, P.; Zhang, L.L.; Wang, H.; Ho, C.T.; Li, S.M.; Shahidi, F.; Zhao, H. Detection of cellular redox reactions and antioxidant activity assays. J. Funct. Foods 2017, 37, 467–479. [Google Scholar] [CrossRef]
- Zhang, Z.D.; Yuan, L.Q.; Ma, Y.B.; Zhou, F.; Gao, Y.; Yang, S.C.; Li, T.L.; Hu, X.H. Exogenous 5-aminolevulinic acid alleviates low-temperature damage by modulating the xanthophyll cycle and nutrient uptake in tomato seedlings. Plant Physiol. Biochem. 2022, 189, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Long, H.; Li, Z.; Suo, H.; Ou, L.; Miao, W.; Deng, W. Study on the Mechanism of Grafting to Improve the Tolerance of Pepper to Low Temperature. Agronomy 2023, 13, 1347–1362. [Google Scholar] [CrossRef]
- Pasbani, B.; Salimi, A.; Aliasgharzad, N.; Hajiboland, R. Colonization with arbuscular mycorrhizal fungi mitigates cold stress through improvement of antioxidant defense and accumulation of protecting molecules in eggplants. Sci. Hortic. 2020, 272, 109575. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, D.; Zhou, X.; Liu, X.; Wang, W.; Bian, J.; He, Z. Application of AMF Alleviates Growth and Physiological Characteristics of Impatiens walleriana under Sub-Low Temperature. Horticulturae 2024, 10, 856. https://doi.org/10.3390/horticulturae10080856
Ye D, Zhou X, Liu X, Wang W, Bian J, He Z. Application of AMF Alleviates Growth and Physiological Characteristics of Impatiens walleriana under Sub-Low Temperature. Horticulturae. 2024; 10(8):856. https://doi.org/10.3390/horticulturae10080856
Chicago/Turabian StyleYe, Deyang, Xiaoting Zhou, Xiaoru Liu, Wenjie Wang, Jinlin Bian, and Zhongqun He. 2024. "Application of AMF Alleviates Growth and Physiological Characteristics of Impatiens walleriana under Sub-Low Temperature" Horticulturae 10, no. 8: 856. https://doi.org/10.3390/horticulturae10080856
APA StyleYe, D., Zhou, X., Liu, X., Wang, W., Bian, J., & He, Z. (2024). Application of AMF Alleviates Growth and Physiological Characteristics of Impatiens walleriana under Sub-Low Temperature. Horticulturae, 10(8), 856. https://doi.org/10.3390/horticulturae10080856