
Evolution of Biogenic Nitrogen from Digestates for Lettuce Fertilization and the Effect on the Bacterial Community

 ,
,  , , ,
, , ,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Liquid Fractions of Digestate
2.2. Agronomic Trial
2.3. Substrate Sampling
2.4. Agronomic and Physiological Measurements
2.5. Substrate Chemical Analysis
2.5.1. Nitrate, Ammonia, Total Nitrogen, and pH Determination
2.5.2. Substrates Respiration
2.6. Substrate Bacterial Analysis
2.6.1. Quantification of 16S Copy Number
2.6.2. Bacterial Community Characterization
2.7. Statistical Analysis
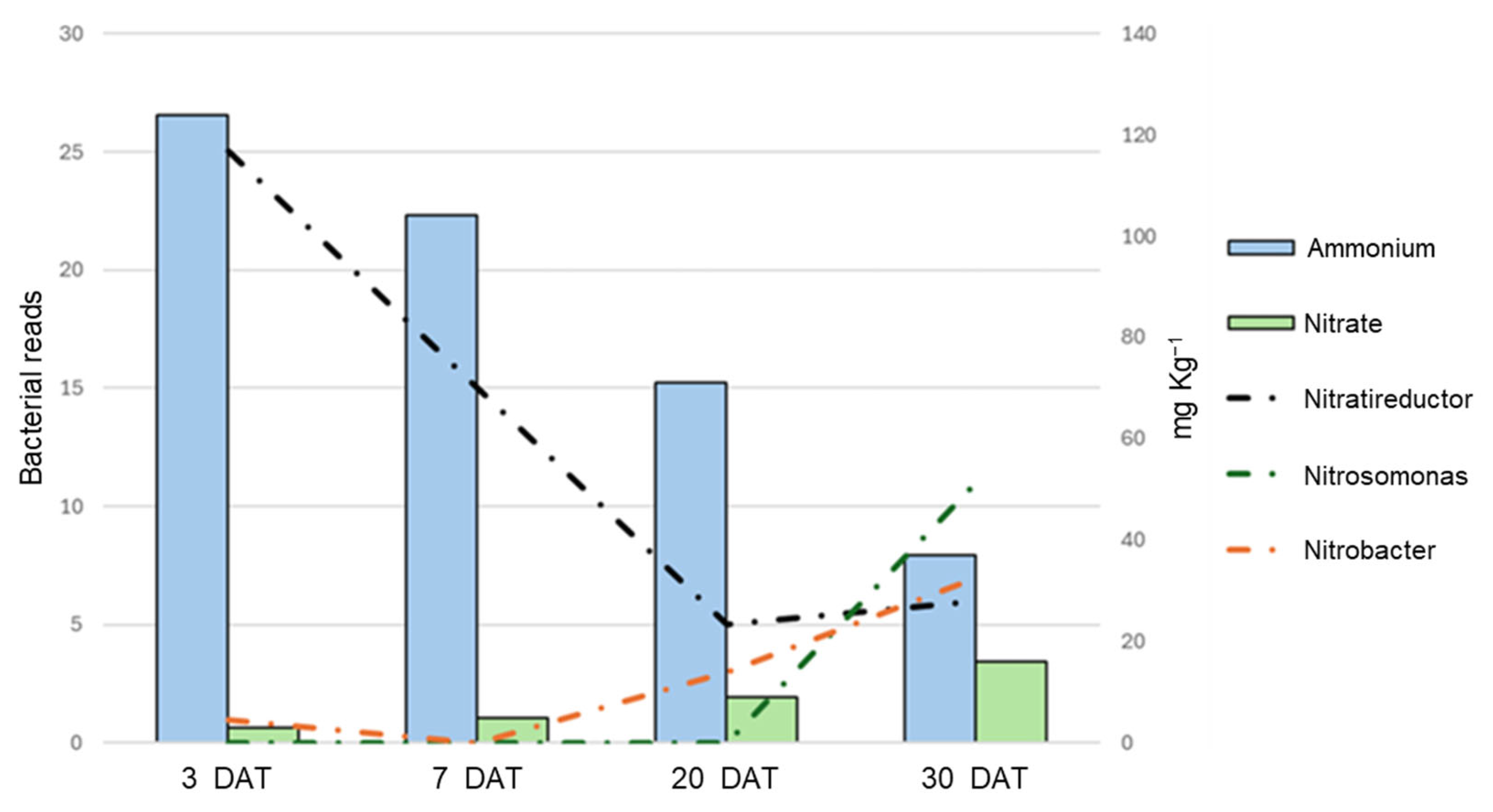
3. Results
3.1. Liquid Fractions of Digestate Characteristics
3.2. Agronomic and Physiological Measurements
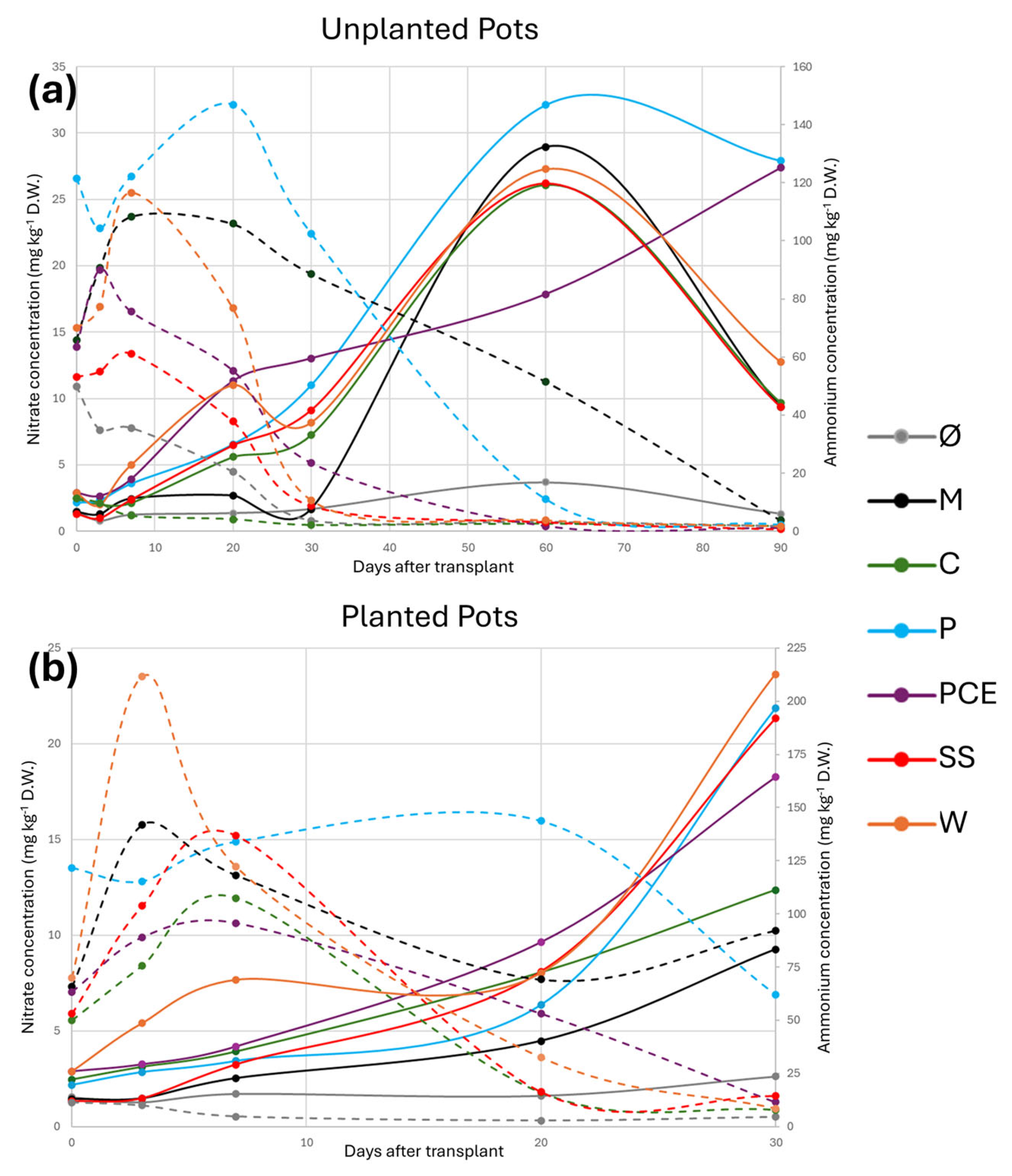
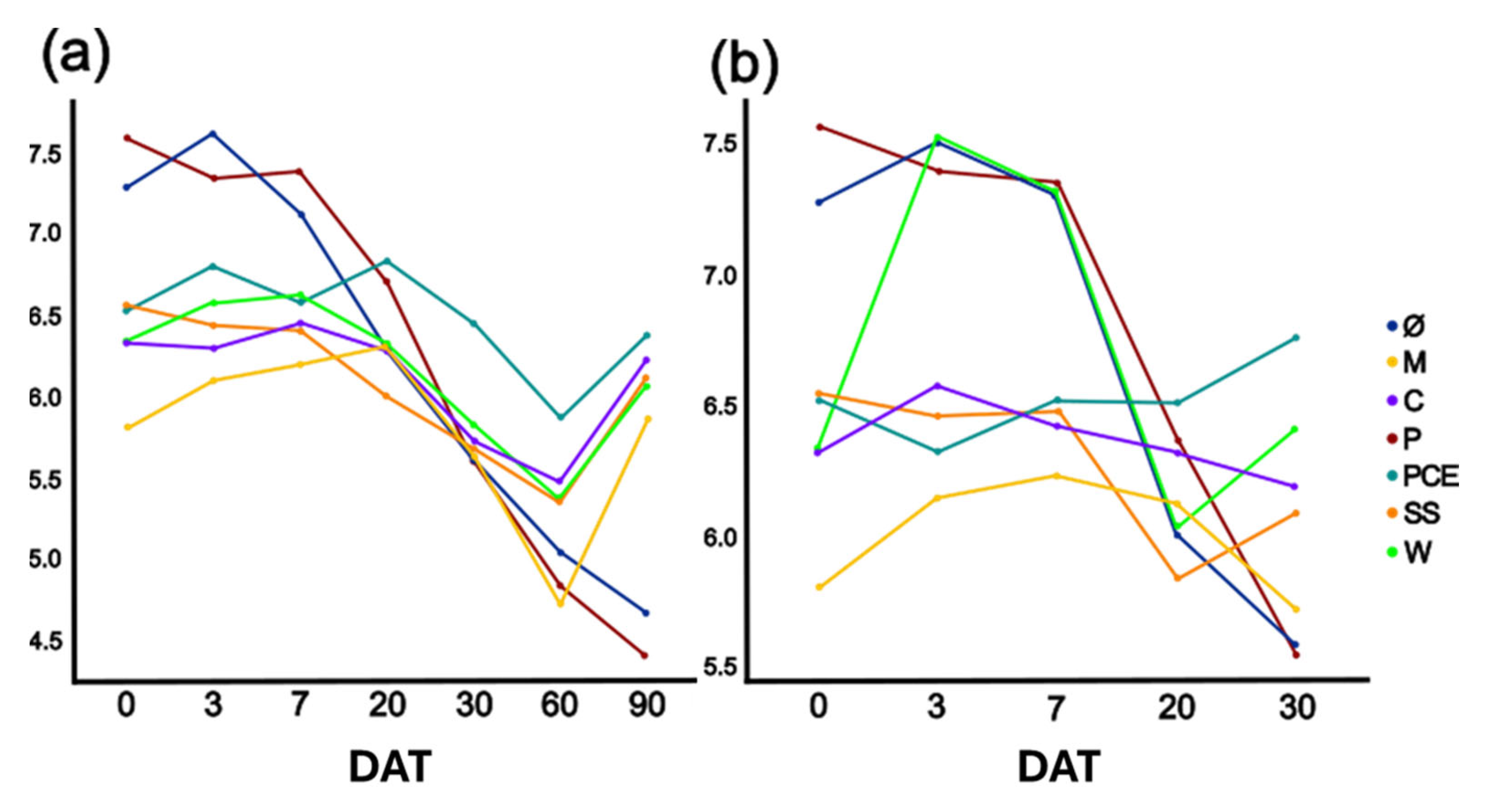
3.3. Substrate Chemical Analysis: Nitrate, Ammonia, Total Nitrogen Determination and pH
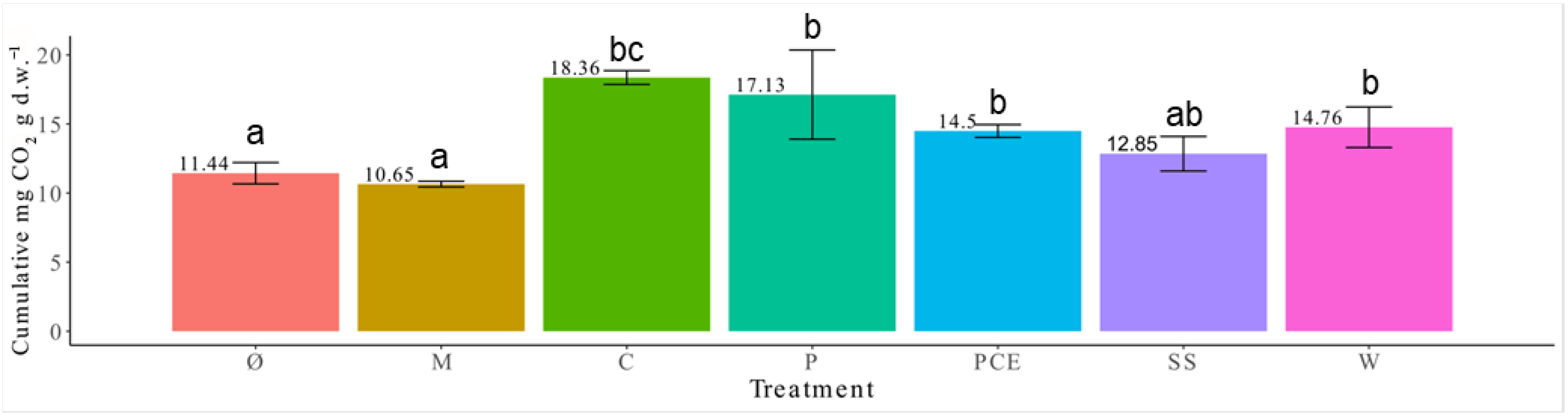
3.4. Substrates Respiration
3.5. Bacterial Quantification
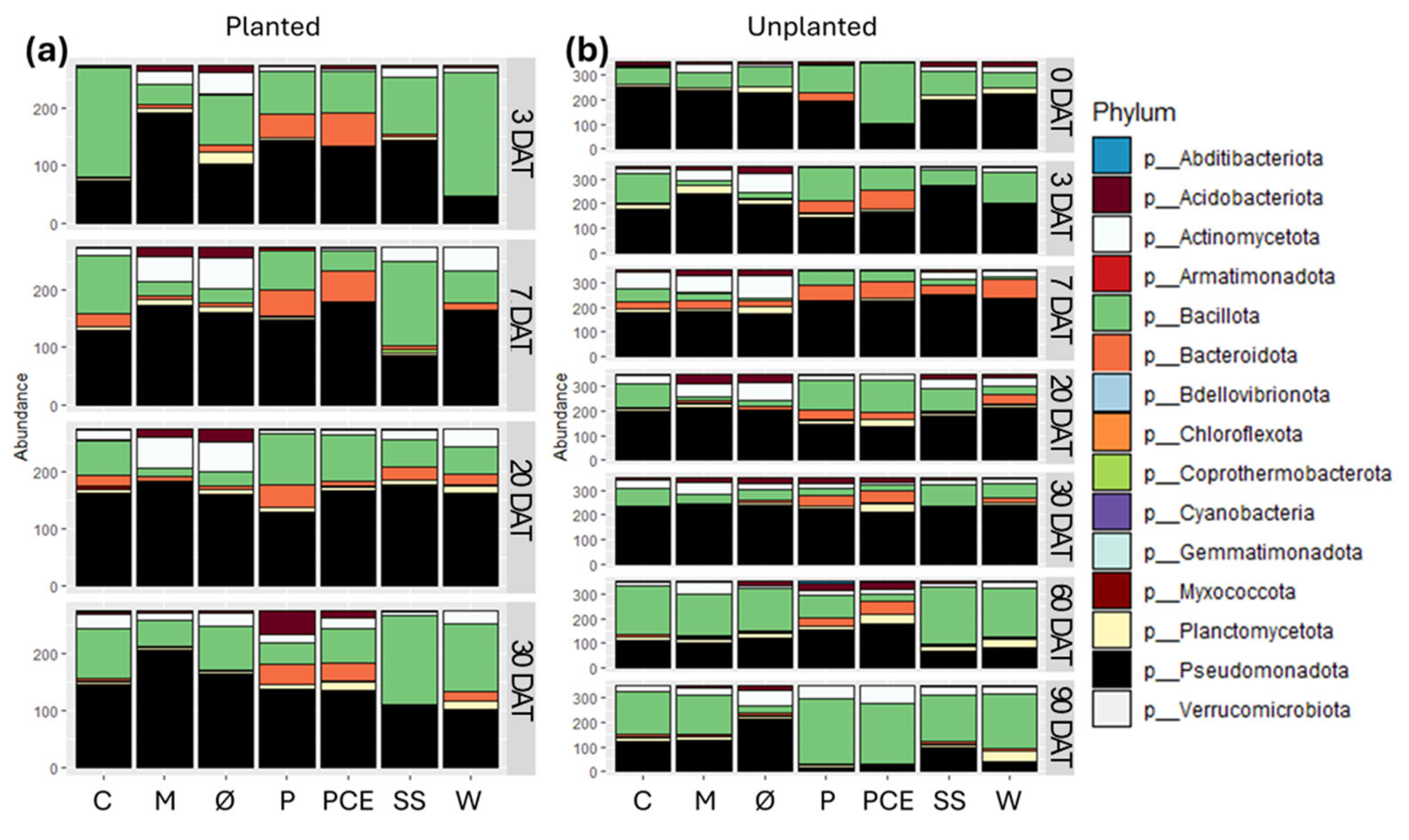
3.6. Bacterial Community Description
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Prospects 2022: Summary of Results; United Nations, Department of Economic and Social Affairs, Population Division: New York, NY, USA, 2022; UN DESA/POP/2022/TR/NO. 3. [Google Scholar]
- Smil, V. Population growth and nitrogen: An exploration of a critical existential link. Environ. Sci. Phil. Popul. Devel. Rev. 1991, 17, 569–601. [Google Scholar] [CrossRef]
- Fertilizers Europe. Forecast of Food, Farming and Fertilizer Use in the European Union 2022–2023. 2023. Available online: https://www.fertiliserseurope.com/wp-content/uploads/2023/01/Forecast-2022-32.pdf (accessed on 16 May 2024).
- Köninger, J.; Lugato, E.; Panagos, P.; Kochupillai, M.; Orgiazzi, A.; Briones, M. Manure management and soil biodiversity: Towards more sustainable food systems in the EU. Agricult. Syst. 2021, 194, 103251. [Google Scholar] [CrossRef]
- European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions. A new Circular Economy Action Plan For a Cleaner and More Competitive Europe. COM(2020) 98 Final. 2020. Available online: https://edz.bib.uni-mannheim.de/edz/doku/wsa/2020/ces-2020-1189-en.pdf (accessed on 16 May 2024).
- Ubando, A.T.; Rivera, D.R.T.; Chen, W.H.; Culaba, A.B. A comprehensive review of life cycle assessment (LCA) of microalgal and lignocellulosic bioenergy products from thermochemical processes. Bioresour. Technol. 2019, 291, 121837. [Google Scholar] [CrossRef]
- Chojnacka, K.; Moustakas, K.; Witek-Krowiak, A. Bio-based fertilizers: A practical approach towards circular economy. Bioresour. Technol. 2020, 295, 122223. [Google Scholar] [CrossRef]
- Tambone, F.; Scaglia, B.; D’Imporzano, G.; Schievano, A.; Orzi, V.; Salati, S.; Adani, F. Assessing amendment and fertilizing properties of digestates from anaerobic digestion through a comparative study with digested sludge and compost. Chemosphere 2010, 81, 577–583. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. Basic Information about Anaerobic Digestion. 2022. Available online: https://www.epa.gov/anaerobic-digestion/basic-information-about-anaerobic-digestion (accessed on 16 May 2024).
- Nkoa, R. Agricultural benefits and environmental risks of soil fertilization with anaerobic digestates: A review. Agron. Sustain. Dev. 2014, 34, 473–492. [Google Scholar] [CrossRef]
- Tambone, F.; Genevini, P.; D’Imporzano, G.; Adani, F. Assessing amendment properties of digestate by studying the organic matter composition and the degree of biological stability during the anaerobic digestion of the organic fraction of MSW. Bioresour. Technol. 2009, 100, 2140–3142. [Google Scholar] [CrossRef]
- Möller, K.; Müller, T. Effects of anaerobic digestion on digestate nutrient availability and crop growth: A review. Eng. Life Sci. 2012, 12, 242–257. [Google Scholar] [CrossRef]
- Tambone, F.; Terruzzi, L.; Scaglia, B.; Adani, F. Composting of the solid fraction of digestate derived from pig slurry: Biological processes and compost properties. Waste Manag. 2015, 35, 55–61. [Google Scholar] [CrossRef]
- Garg, R.N.; Pathak, H.; Das, D.K.; Tomar, R.K. Use of fly ash and biogas slurry for improving wheat yield and physical properties of the soil. Environ. Monit. Assess. 2005, 107, 1–9. [Google Scholar] [CrossRef]
- Riva, C.; Orzi, V.; Carozzi, M.; Acutis, M.; Boccasile, G.; Lonati, S.; Tambone, F.; D’Imporzano, G.; Adani, F. Short-term experiments in using digestate products as substitutes for mineral (N) fertilizer: Agronomic performance, odours, and ammonia emission impacts. Sci. Total Environ. 2016, 547, 206–214. [Google Scholar] [CrossRef]
- Tambone, F.; Adani, F. Nitrogen mineralisation from digestate in comparison to sewage sludge, compost and urea in a laboratory incubated soil experiment. J. Plant Nutr. Soil Sci. 2017, 180, 355–365. [Google Scholar] [CrossRef]
- Sharifi, M.; Baker, S.; Hojabri, L.; Hajiaghaei-Kamrani, M. Short-term nitrogen dynamics in a soil amended with anaerobic digestate. Can. J. Soil Sci. 2019, 99, 242–257. [Google Scholar] [CrossRef]
- Zilio, M.; Pigoli, A.; Rizzi, B.; Geromel, G.; Meers, E.; Schoumans, O.; Giordano, A.; Adani, F. Measuring ammonia and odours emissions during full field digestate use in agriculture. Sci. Total Environ. 2021, 782, 146882. [Google Scholar] [CrossRef]
- Alburquerque, J.A.; de la Fuente, C.; Ferrer-Costa, A.; Carrasco, L.; Cegarra, J.; Abad, M.; Bernal, M.P. Assessment of fertilizer potential of digestates from farm and agroindustrial residues. Biomass Bioenerg. 2012, 40, 181–189. [Google Scholar] [CrossRef]
- Luo, Y.; Chavez-Rico, V.N.; Sechi, V.; Bezemer, T.M.; Buisman, C.J.N.; ter Heijne, A. Effect of organic amendments obtained from different pretreatment technologies on soil microbial community. Environ. Res. 2023, 232, 116346. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater, 20th ed.; APHA: Washington, DC, USA, 1998. [Google Scholar]
- Lenzi, A.; Orlandini, A.; Bulgari, R.; Ferrante, A.; Bruschi, P. Antioxidant and mineral composition of three wild leafy species: A comparison between microgreens and baby greens. Foods 2019, 8, 487. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chorophylls and carotenoids: Pigments of photosynthetic biomembranes. Meth. Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissues by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Branca, F.; Romano, D. Enhancing the quality of two species of baby leaves sprayed with moringa leaf extract as biostimulant. Agronomy 2021, 11, 1399. [Google Scholar] [CrossRef]
- ISO 16072; Soil Quality—Laboratory Methods for Determination of Microbial Soil Respiration. International Organization for Standardization: Geneva, Switzerland, 2002.
- Sims, A.; Horton, J.; Gajaraj, S.; McIntoch, S.; Miles, R.J.; Mueller, R.; Reed, R.; Hu, Z. Temporal and spatial distribution of ammonia-oxidising archaea and bacteria and their ratio as indicator of oligotrophic conditions in natural wetlands. Water Res. 2012, 46, 4121–4129. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Kumar, K.; Debnath, P.; Singh, S.; Kumar, N. An Overview of Plant Phenolics and Their Involvement in Abiotic Stress Tolerance. Stresses 2023, 3, 570–585. [Google Scholar] [CrossRef]
- Fedeli, R.; Celletti, S.; Loppi, S.; Vannini, A. Comparison of the Effect of Solid and Liquid Digestate on the Growth of Lettuce (Lactuca sativa L.) Plants. Agronomy 2023, 13, 782. [Google Scholar] [CrossRef]
- Weimers, K.; Bergstrand, K.J.; Hultberg, M.; Asp, H. Liquid Anaerobic Digestate as Sole Nutrient Source in Soilless Horticulture—Or Spiked with Mineral Nutrients for Improved Plant Growth. Front. Plant Sci. 2022, 13, 770179. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Singh, P.; Ibrahim, M.H.; Hashim, R. Land Application of Sewage Sludge: Physicochemical and Microbial Response. Rev. Environ. Contam. Toxicol. 2012, 214, 41–61. [Google Scholar]
- Tan, F.; Zhu, Q.; Guo, X.; He, L. Effects of Digestate on Biomass of a Selected Energy Crop and Soil Properties. J. Sci. Food Agric. 2021, 101, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Zhang, S.; Wang, Y.; Wang, R. Advantages of the Integrated Pig-Biogas-Vegetable Greenhouse System in North China. Ecol. Eng. 2005, 24, 175–183. [Google Scholar] [CrossRef]
- Šamec, D.; Karalija, E.; Šola, I.; Vujcic Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Rashid, M.T.P.V.; Parkin, G. Predicting nitrogen fertilizer requirements for corn by chlorophyll meter under different N availability conditions. Can. J. Soil Sci. 2005, 85, 149–159. [Google Scholar] [CrossRef]
- European Union. Commission Regulation (EU) 2023/915 on maximum levels for certain contaminants in food and repealing Regulation (EC) No. 1881/2006. Off. J. Eur. Union 2023, 119, 103–157. [Google Scholar]
- M’hamdi, M.; Boughttas, I.; Chick-Rouhou, E.; Bettaieb, T. Effect of different levels of nitrogen fertilizer on morphological and physiological parameters and nitrates accumulation of lettuce cultivars (Lactuca sativa L.). Res. Plant Biol. 2014, 4, 27–38. [Google Scholar]
- Goberna, M.; Podmirseg, S.M.; Waldhuber, S.; Knapp, B.A.; García, C.; Insam, H. Pathogenic bacteria and mineral N in soil following the land spreading of biogas digestates and fresh manure. Appl. Soil Ecol. 2011, 49, 18–25. [Google Scholar] [CrossRef]
- Song, J.; Wang, Y.; Zhang, S.; Song, Y.; Xue, S.; Liu, L.; Lvy, X.; Wang, X.; Yang, G. Coupling biochar with anaerobic digestion in a circular economy perspective: A promising way to promote sustainable energy, environment and agriculture development in China. Renew. Sustain. Energ. Rev. 2021, 144, 110973. [Google Scholar] [CrossRef]
- Sublett, W.L.; Barickman, C.; Sams, C.E. The Effect of Environment and Nutrients on Hydroponic Lettuce Yield, Quality, and Phytonutrients. Horticulturae 2018, 4, 48. [Google Scholar] [CrossRef]
- Flavel, T.C.; Murphy, D.V. Carbon and nitrogen mineralization rates after application of organic amendments to soil. J. Environ. Qual. 2006, 35, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Brandon, M.; Juarez, M.F.D.; Zangerle, M.; Insam, H. Effects of digestate on soil chemical and microbiological properties: A comparative study with compost and vermicompost. J. Hazard Mater. 2016, 302, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Odlare, M.; Arthurson, V.; Pell, M.; Svensson, K.; Nehrenheim, E.; Abubaker, J. Land application of organic waste—Effects on the soil ecosystem. Appl. Energy 2011, 88, 2210–2218. [Google Scholar] [CrossRef]
- Chen, J.S.; Toth, J.; Kasap, M. Nitrogen-fixation genes and nitrogenase activity in Clostridium acetobutylicum and Clostridium beijerinckii. J. Ind. Microbiol. Biotech. 2001, 27, 281–286. [Google Scholar] [CrossRef]
- Wainaina, S.; Lukitawesa, K.; Awasthi, M.; Taherzadeh, M.J. Bioengineering of anaerobic digestion for volatile fatty acids, hydrogen or methane production: A critical review. Bioengineered 2019, 10, 437–458. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, L.; Hassan, M.; Xie, B. Succession of the functional microbial communities and the metabolic functions in maize straw composting process. Bioresour. Technol. 2018, 256, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.Z.; Ma, S.C.; Wang, S.P.; Wang, T.T.; Sun, Z.Y.; Tang, Y.Q.; Deng, Y.; Kida, K. A comparative study of composting the solid fraction of dairy manure with or without bulking material: Performance and microbial community dynamics. Bioresour. Technol. 2018, 247, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Hamdali, H.; Hafidi, M.; Virolle, M.J.; Ouhdouch, Y. Growth promotion and protection against damping-off of wheat by two rock phosphate solubilizing actinomycetes in a P-deficient soil under greenhouse conditions. Appl. Soil Ecol. 2008, 40, 510–517. [Google Scholar] [CrossRef]
- Jog, R.; Pandya, M.; Nareshkumar, G.; Rajkumar, S. Mechanism of phosphate solubilization and antifungal activity of Streptomyces spp. isolated from wheat roots and rhizosphere and their application in improving plant growth. Microbiology 2014, 160, 778–788. [Google Scholar] [CrossRef]
- Bastabak, B.; Kocar, G. A review of the biogas digestate in agricultural framework. J. Mater. Cycles Waste Manag. 2020, 22, 1318–1327. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Solids | pH | N-NH4 | TKN | N-NH4 | Ashes | |
|---|---|---|---|---|---|---|
| (%FM) | (g kg−1 DM) | (%TKN) | (%DM) | |||
| C | 3.28 | 8.21 | 0.98 | 2.56 | 38.3 | 33.06 |
| P | 5.24 | 8.31 | 2.35 | 3.36 | 69.9 | 46.31 |
| PCE | 3.62 | 8.63 | 1.44 | 2.59 | 55.6 | 33.85 |
| SS | 10.98 | 8.15 | 4.30 | 8.66 | 49.7 | 45.60 |
| W | 5.08 | 8.92 | 4.23 | 6.42 | 65.9 | 36.58 |
| Calcium (Ca) | Iron (Fe) | Magnesium (Mg) | Sodium (Na) | Phosphorus (P) | Potassium (K) | |
|---|---|---|---|---|---|---|
| Ø | 19.61 ± 1.22 d | 322.60 ± 8.04 d | 3.97 ± 0.36 a | 4.36 ± 0.09 b | 1.55 ± 0.05 a | 50.68 ± 2.24 b |
| M | 10.82 ± 0.58 a | 174.88 ± 4.56 bc | 3.39 ± 0.11 a | 1.01 ± 0.12 a | 8.56 ± 0.22 e | 80.72 ± 7.63 cd |
| C | 16.17 ±1.49 bcd | 173.77 ± 15.13 bc | 4.53 ± 0.54 a | 11.40 ± 0.62 d | 3.89 ± 0.75 b | 73.27 ± 15.41 bc |
| P | 15.69 ± 0.63 bc | 156.51 ± 7.25 ab | 4.30 ± 0.09 a | 2.98 ± 0.23 ab | 8.27 ± 0.53 de | 79.74 ± 2.19 cd |
| PCE | 13.43 ± 0.47 ab | 130.40 ± 1.79 a | 3.53 ± 0.43 a | 4.33 ± 1.25 b | 7.84 ± 0.02 de | 102.13 ± 2.09 d |
| SS | 17.59 ± 1.78 cd | 203.06 ± 22.24 c | 7.26 ± 0.52 b | 11.32 ± 0.34 d | 6.80 ± 0.76 cd | 16.80 ± 2.61 a |
| W | 14.21 ± 0.06 abc | 164.49 ± 3.48 ab | 4.26 ± 0.21 a | 7.70 ± 1.12 c | 5.48 ± 0.22 c | 54.18 ± 5.25 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caterino, D.; Follador, A.; Galbusera, J.; Guffanti, D.; Cocetta, G.; Geuna, F.; Dell’Orto, M.; Scaglia, B.; Passera, A.; Riva, E.; et al. Evolution of Biogenic Nitrogen from Digestates for Lettuce Fertilization and the Effect on the Bacterial Community. Horticulturae 2024, 10, 685. https://doi.org/10.3390/horticulturae10070685
Caterino D, Follador A, Galbusera J, Guffanti D, Cocetta G, Geuna F, Dell’Orto M, Scaglia B, Passera A, Riva E, et al. Evolution of Biogenic Nitrogen from Digestates for Lettuce Fertilization and the Effect on the Bacterial Community. Horticulturae. 2024; 10(7):685. https://doi.org/10.3390/horticulturae10070685
Chicago/Turabian StyleCaterino, Domenico, Alessia Follador, Jonathan Galbusera, Davide Guffanti, Giacomo Cocetta, Filippo Geuna, Marta Dell’Orto, Barbara Scaglia, Alessandro Passera, Elisabetta Riva, and et al. 2024. "Evolution of Biogenic Nitrogen from Digestates for Lettuce Fertilization and the Effect on the Bacterial Community" Horticulturae 10, no. 7: 685. https://doi.org/10.3390/horticulturae10070685
APA StyleCaterino, D., Follador, A., Galbusera, J., Guffanti, D., Cocetta, G., Geuna, F., Dell’Orto, M., Scaglia, B., Passera, A., Riva, E., Casati, P., Ferrante, A., & Tambone, F. (2024). Evolution of Biogenic Nitrogen from Digestates for Lettuce Fertilization and the Effect on the Bacterial Community. Horticulturae, 10(7), 685. https://doi.org/10.3390/horticulturae10070685