
Exploring the Mechanisms Underlying Petal Pigmentation Differences in Two Cultivars of Physalis philadelphica Based on HPLC and NGS

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Experimental Design
2.2. Determination of Pigment Content and Transcriptome Sequencing
2.2.1. Determination of Total Anthocyanin Content
2.2.2. Determination of Carotenoid Content
2.2.3. Determination of Anthocyanin and β-Carotene Contents
2.2.4. Transcriptome Sequencing
2.2.5. Single Nucleotide Polymorphism (SNP) and Indel Analyses
2.2.6. Validation of Next-Generation Sequencing (NGS) by Real-Time Quantitative PCR (qRT-PCR)
3. Results
3.1. Analyses of Color-Related Components
3.1.1. Total Anthocyanin Content
3.1.2. Carotenoid Content
3.1.3. Delphinidin, Cyanidin, Pelargonidin and Malvidin Contents
3.1.4. β-Carotene Content
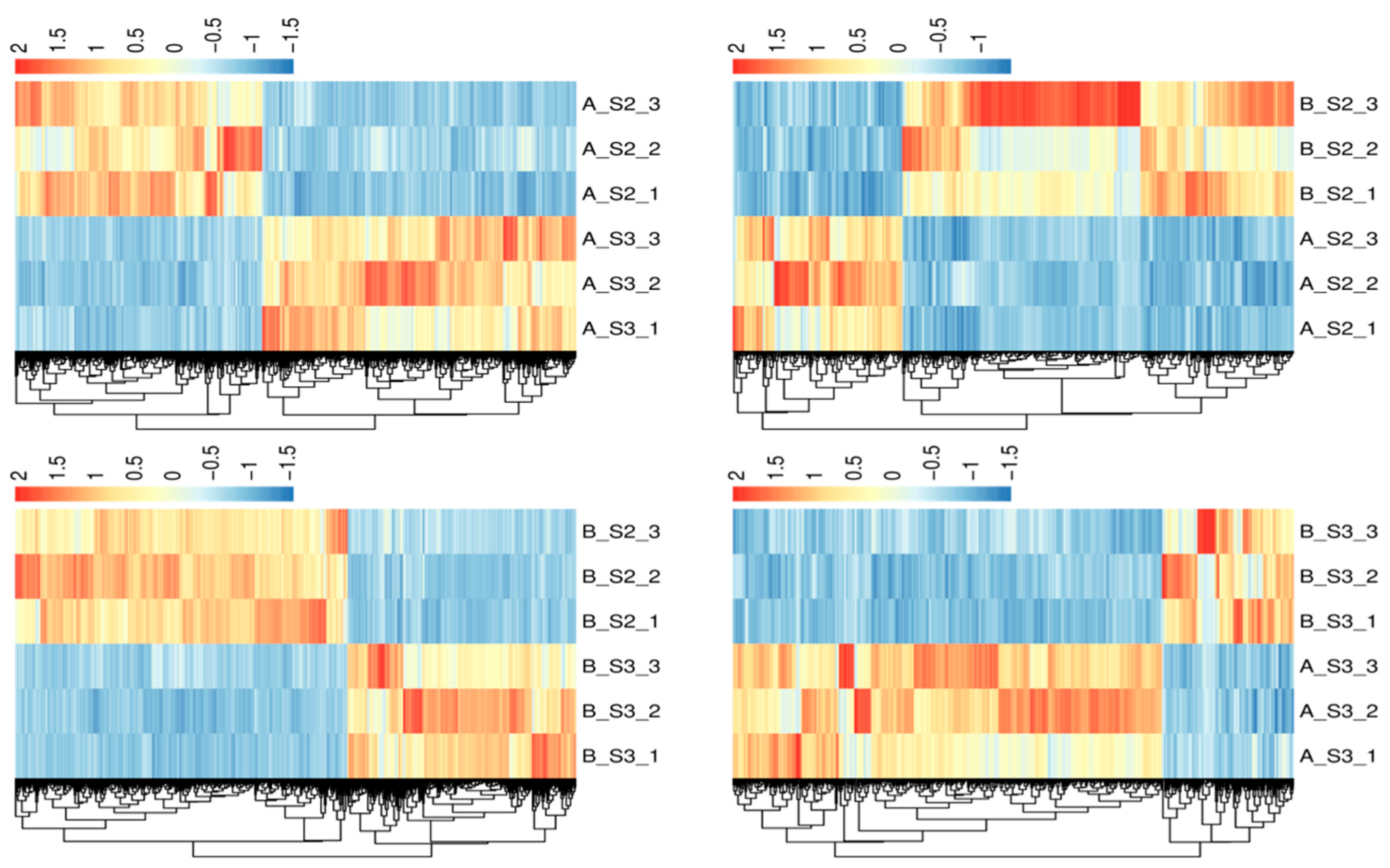
3.2. Transcriptomic Analyses
3.2.1. High-Throughput Sequencing Results
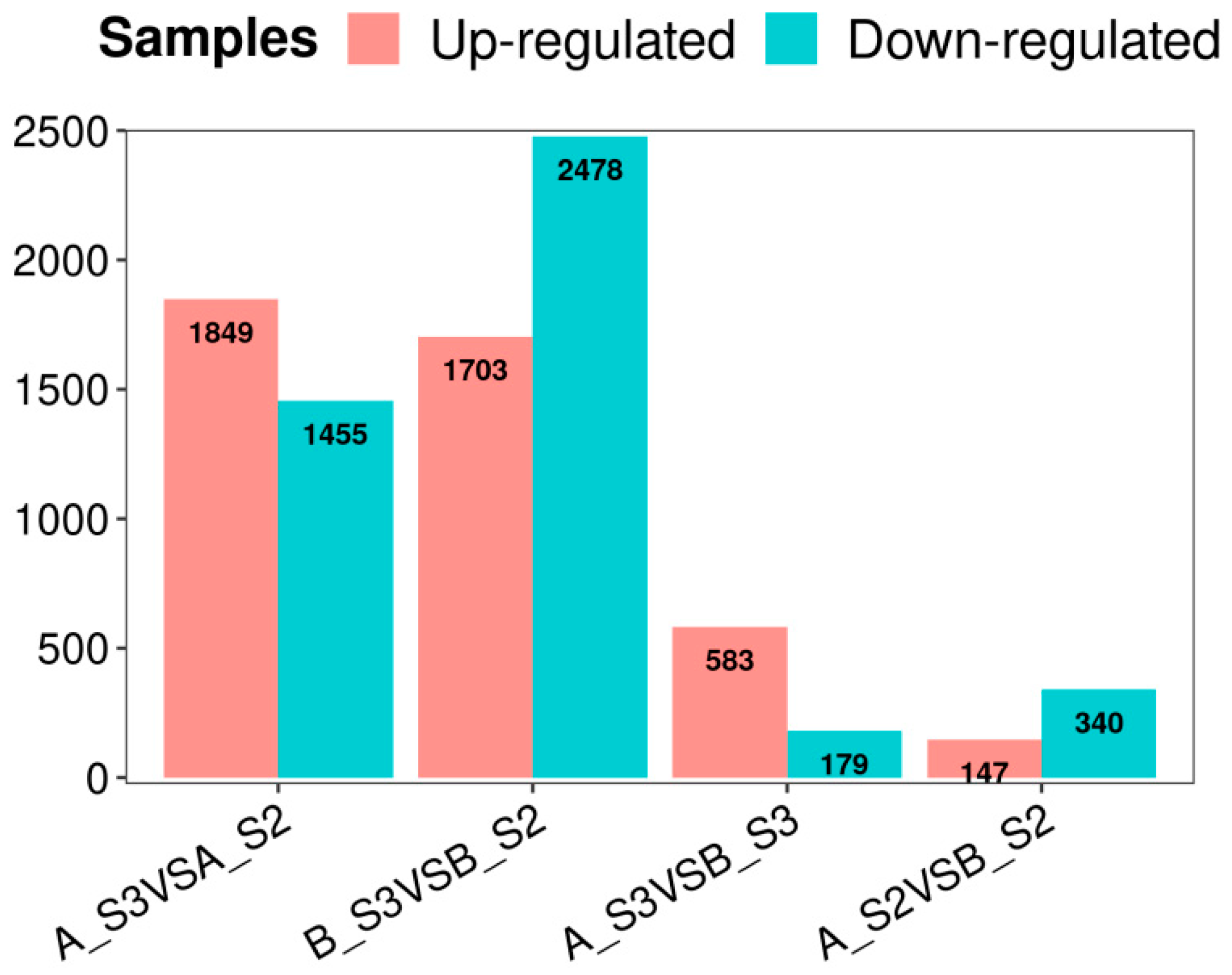
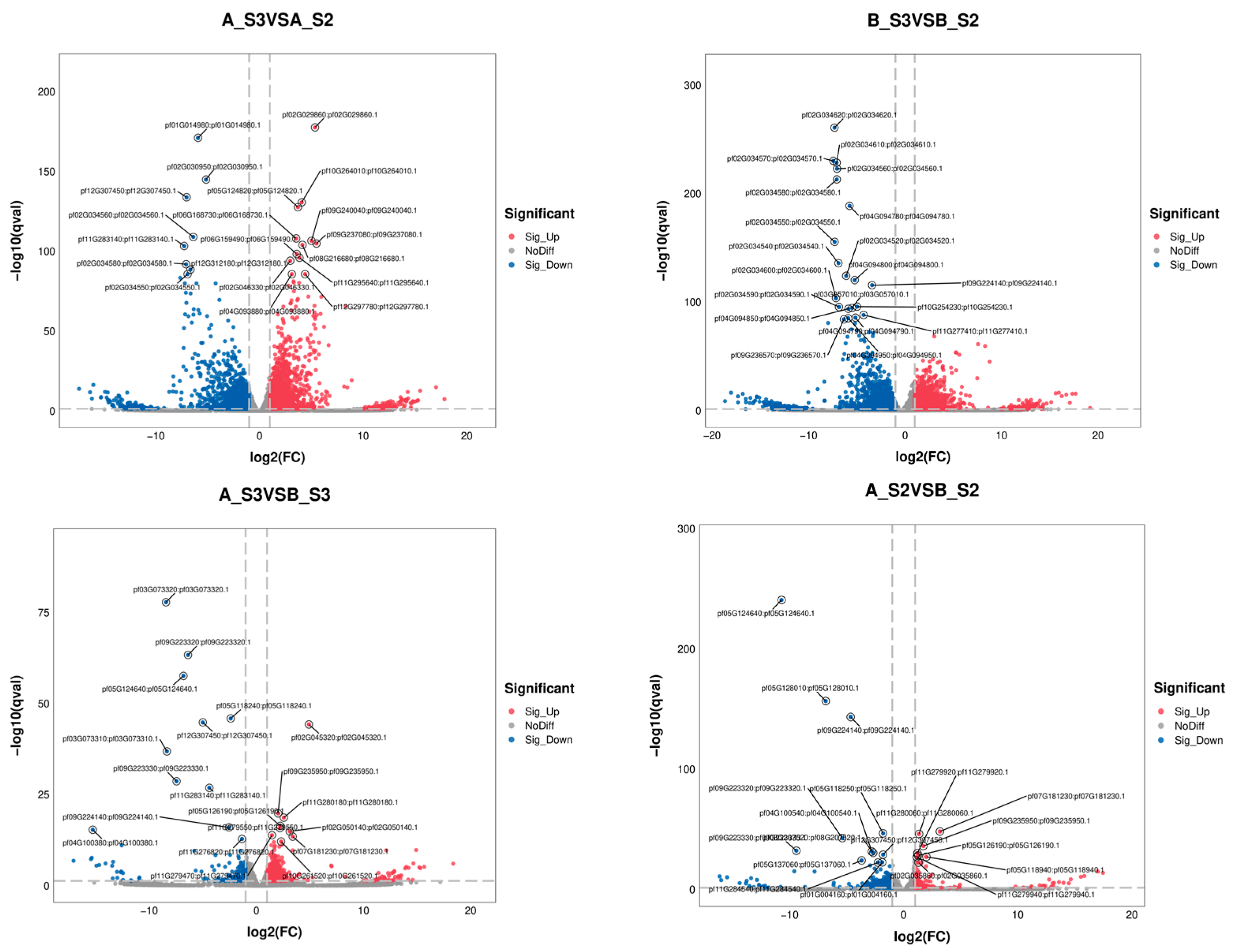
3.2.2. Identification of DEGs
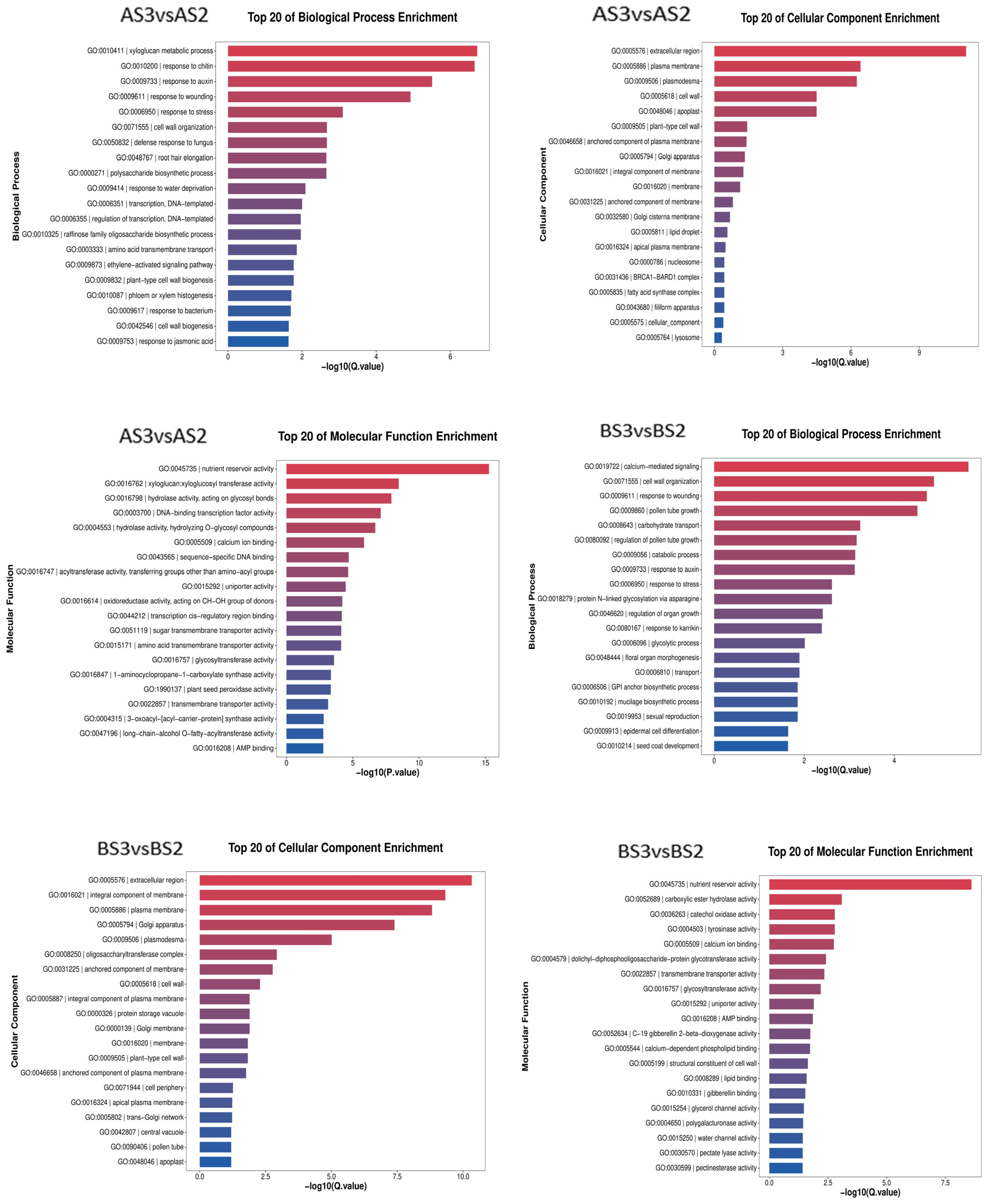
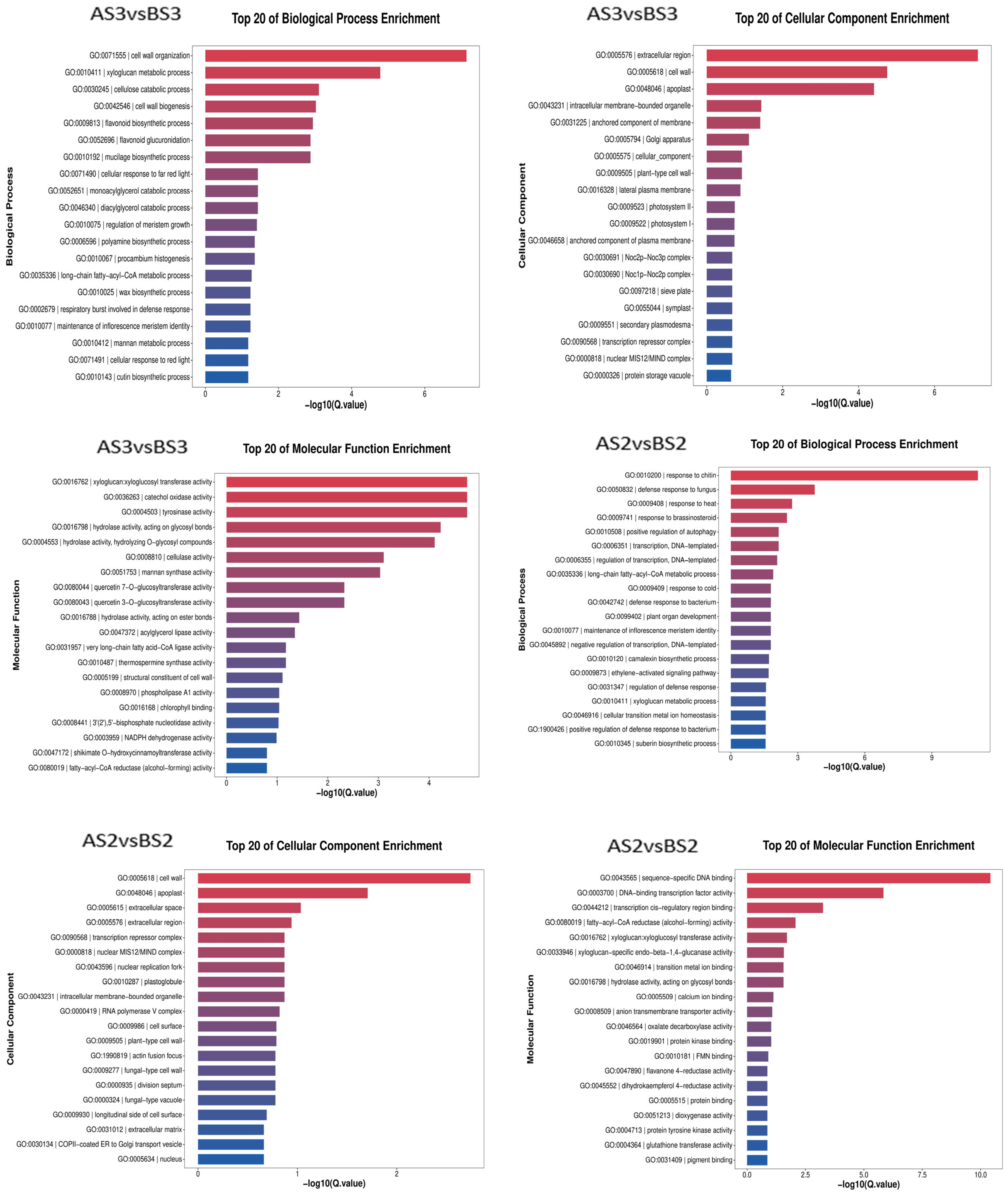
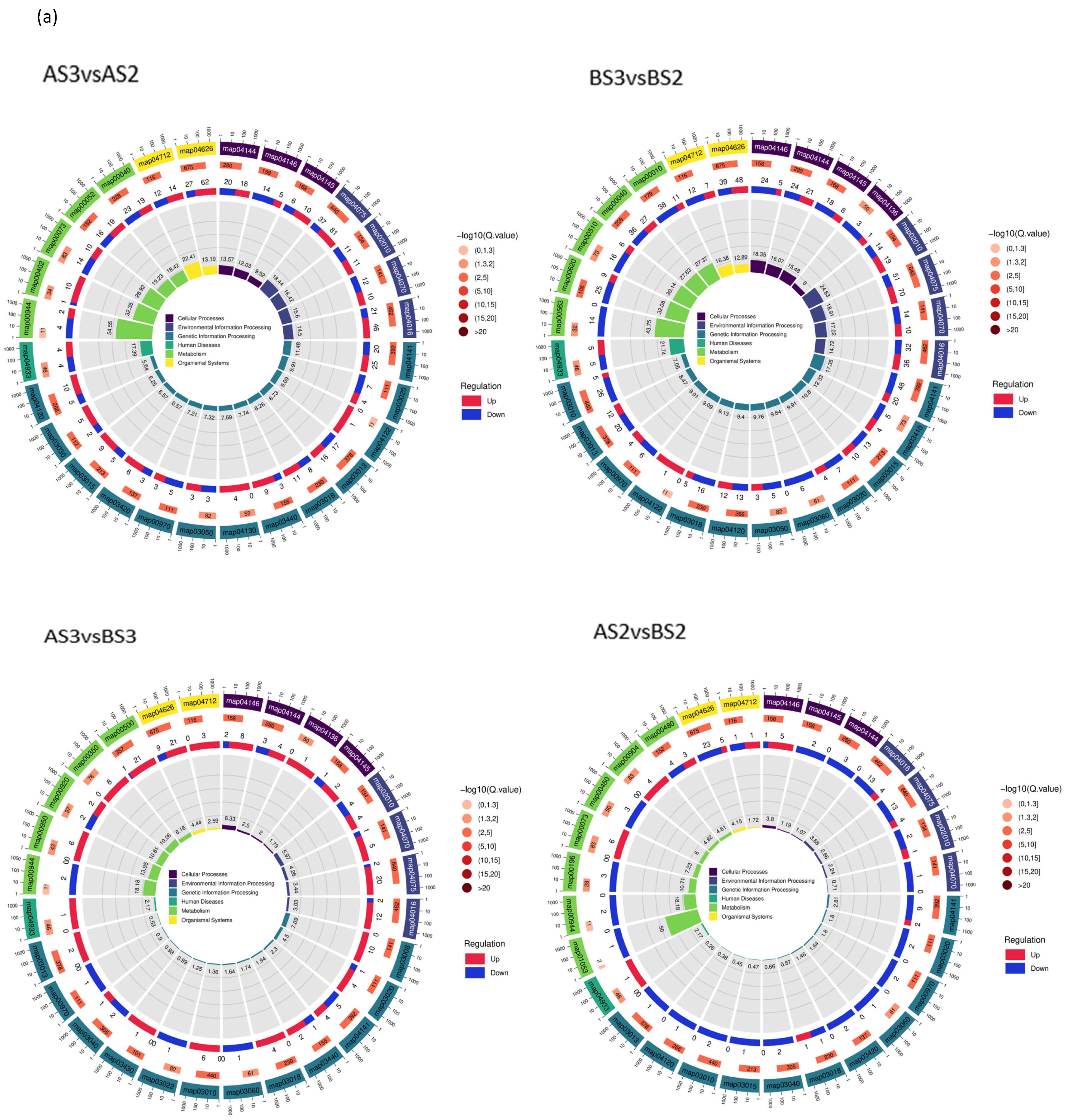
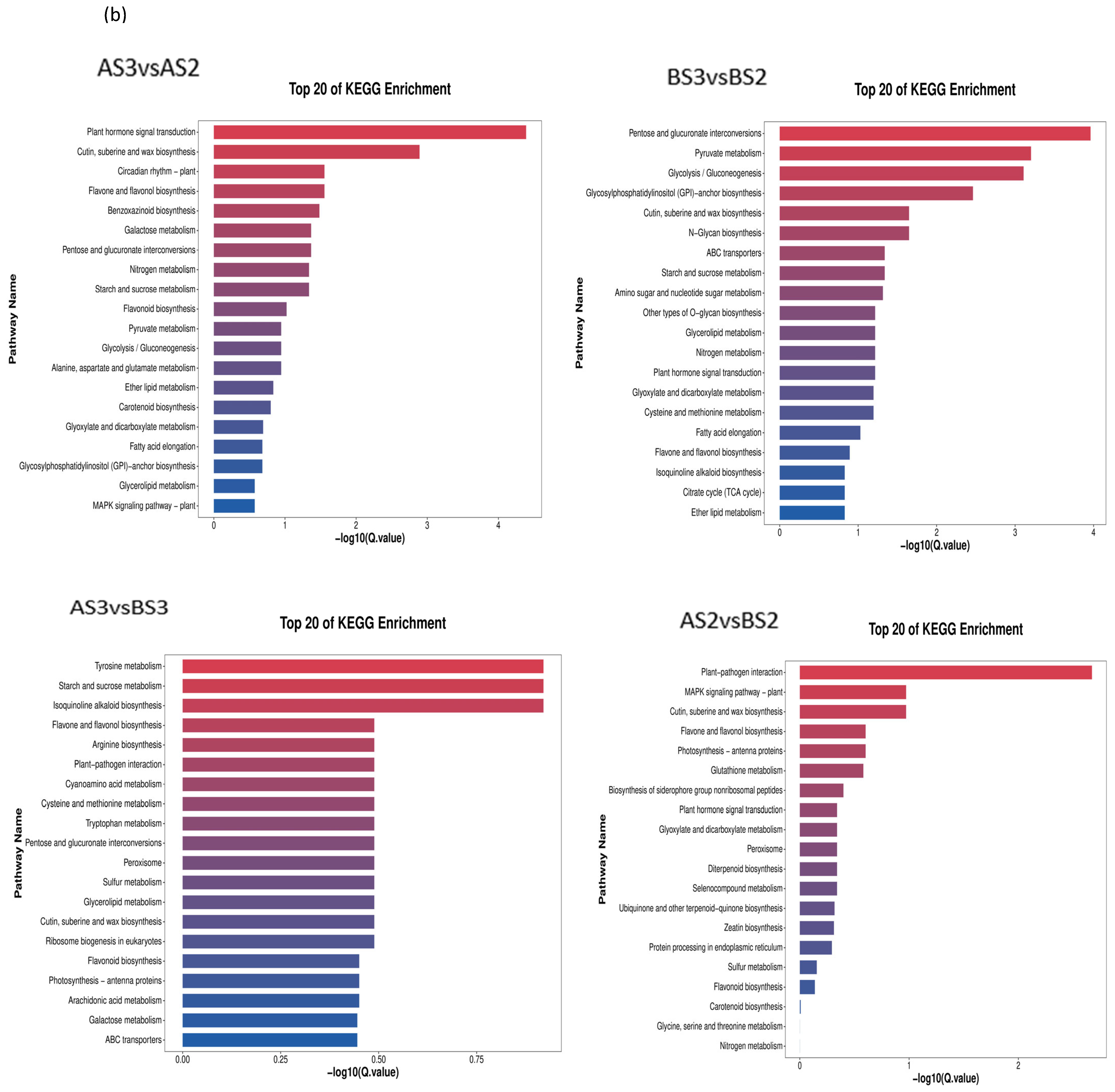
3.2.3. GO Enrichment and KEGG Analysis of Petal Coloring-Related DEGs in P. philadelphica
3.2.4. Mining of Genes Related to Flavonoids, Flavone and Flavonol, and Carotenoid Biosyntheses
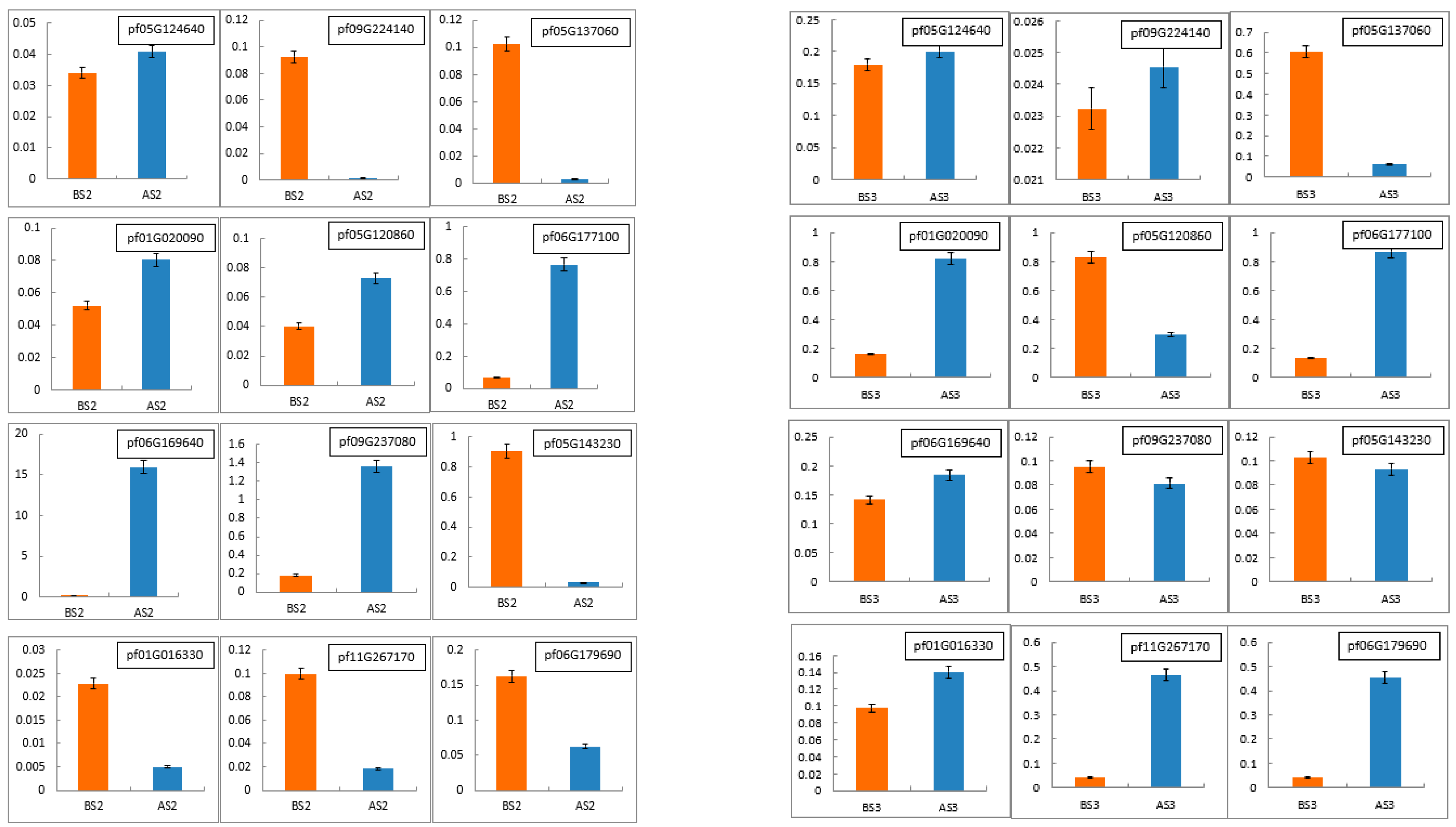
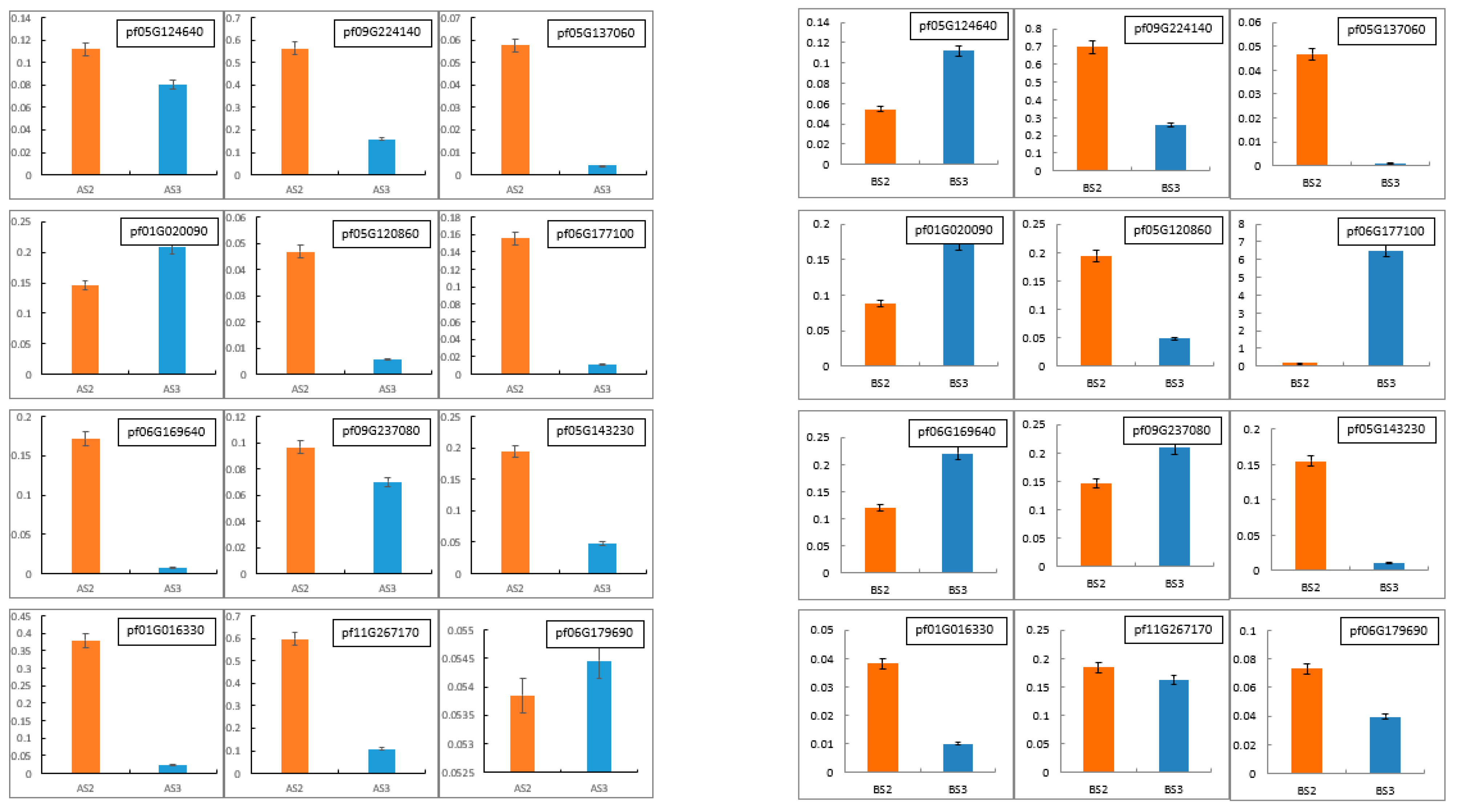
3.2.5. qRT-PCR Validation of NGS Results
4. Discussion
4.1. Influence of Key Enzyme Genes on Petal Coloration of P. philadelphica
4.2. TFs Associated with Petal Coloration in P. philadelphica
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- González-Pérez, J.E.; Guerrero-Beltrán, J.N. Tomatillo or husk tomato (Physalis philadelphica and Physalis ixocarpa): A review. Sci. Hortic. 2021, 288, 110306. [Google Scholar] [CrossRef]
- Yang, B.; Li, X.; Liu, Y.; Kuang, X. Research Progress on Physalis philadelphica. Chin. Tradit. Herb. Drugs 2017, 33, 104–111. [Google Scholar]
- Zhu, Y.; Jiao, K.; Feng, S.; Wang, H. Efficient cultivation techniques for dual-purpose medicinal and edible plant maosuanjiang. Zhejiang Agric. Sci. 2018, 59, 2. [Google Scholar]
- Mol, J.; Grotewold, E.; Koes, R. How genes paint flowers and seeds. Trends Plant Sci. 1998, 6, 212–217. [Google Scholar] [CrossRef]
- Toyama-Kato, Y.; Yoshida, K.; Fujimori, E.; Haraguchi, H.; Shimizu, Y.; Kondo, T. Analysis of metal elements of hydrangea sepals at various growing stages by ICP-AES. Biochem. Eng. J. 2003, 14, 237–241. [Google Scholar] [CrossRef]
- Wu, E.Q.; Tai, L.H. Qualitative analysis of petal pigments. Inn. Mong. Norm. Univ. (Nat. Sci. Ed.) 2002, 31, 64–67. [Google Scholar]
- Xia, X.; Gong, R.; Zhang, C. Composition of anthocyanins and coloration mechanism in petals of Rhododendron with different colors. Agric. Sci. 2022, 38, 207–213. [Google Scholar]
- Zheng, Q.Z.; Zhao, B.; Zeng, H.M.; Shen, H.F.; Xu, J.J. Comparative analysis of pigment composition and content in petals of three different-colored Rhododendron beauties from Qinling. Northwest For. Univ. 2017, 32, 62–68. [Google Scholar]
- Zhu, Y.; Xu, Z.; Wang, Q.; Wang, Z.; Shi, Y. Comparison of leaf epidermal morphology of different ploidy levels in Dioscorea polystachya. Plant Res. 2010, 30, 267–272. [Google Scholar]
- Liu, S.H.; Zang, D.D.; Sun, Y.; Li, J.X.; Zhao, H.T. Research progress on the biosynthetic pathway and key enzymes of anthocyanin. Soil Crop 2022, 11, 336–346. [Google Scholar]
- Qiao, S.H.; Wang, Q.; Hu, L.D.; Ma, L.; Zhang, Y.H.; Wang, H.X. Advances in the application of anthocyanins in food quality indicators. Packag. Eng. 2022, 43, 49–58. [Google Scholar]
- Xu, Y.Y.; Zhang, Y.H.; Qian, Y.Y.; Huang, H.X.; Wu, Y.W.; Zhao, D.; Xu, T.H. Research progress on the pharmacological effects of anthocyanin components. Mod. Drugs Clin. Med. 2022, 37, 1886–1891. [Google Scholar]
- Zhao, D. Advances in research on genes related to the biosynthetic pathway of plant anthocyanins and their genetic engineering modifications. J. Trop. Biol. 2012, 3, 92–98. [Google Scholar]
- Xu, G.; Tan, G.; Zhu, C.; He, D. Genetic diversity analysis of F1 populations from different cauliflower varieties. North. Hortic. 2022, 17, 1–8. [Google Scholar]
- Kong, W.; Ling, J.; Yang, J. Identification of viruses and analysis of their small RNA features in deformed mulberry leaves. Plant Prot. 2022, 48, 251–255+268. [Google Scholar]
- Ge, W.; Wang, H.; Liu, M.; Zhang, L.; Liu, D.; Zhou, B.; Zuo, Z. Regulatory effect of curcumin on miRNA-mRNA expression profile in mice with autoimmune hepatitis. Chin. J. Tradit. Chin. Med. 2022, 37, 2547–2552. [Google Scholar]
- Li, J.; Xu, Y.; Cai, S.; Wang, J. Transcriptome sequencing and bioinformatics analysis of Schisandra chinensis from China and Thailand. J. Guangzhou Univ. Chin. Med. 2022, 39, 2387–2393. [Google Scholar]
- Keppetipola, N.M.; Yeom, K.-H.; Hernandez, A.L.; Bui, T.; Sharma, S.; Black, D.L. Multiple determinants of splicing repression activity in the polypyrimidine tract binding proteins, PTBP1 and PTBP2. RNA 2016, 22, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakeeld, M.J.; Smyth, G.K.; Oshlack, A. goseq: Gene Ontology testing for RNA-seq datasets. R Bioconductor 2012, 8, 1–25. [Google Scholar]
- Fang, Z.; Jiang, C.; Zhou, D.; Pan, S.; Ye, X. Identification and analysis of the Aux/IAA gene family in the March plum and its red-fleshed mutant. Nucl. Agric. Sci. 2020, 34, 247–255. [Google Scholar]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Li, H. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Fan, S.G.; Li, H.P.; Hu, C.Q. Genetic diversity study on four geographical populations of sea cucumber Apostichopus japonicus in the South China Sea. Fish. Sci. 2018, 37, 404–408. [Google Scholar]
- Wang, G.; Guo, Z.; Wang, X.; Guan, S.L.; Gao, H.; Qi, K.; Gu, C.; Zhang, S. Identification and testing of reference genes for qRT-PCR analysis during pear fruit development. Biologia 2022, 77, 2763–2777. [Google Scholar] [CrossRef]
- Zheng, Y.; Ma, Y.; Luo, J.; Li, J.; Zheng, X.; Gong, H.; Deng, L.; Zhao, G.; Luo, C.; Liu, X. Identification and Analysis of Reference and Tissue-Specific Genes in Bitter Gourd Based on Transcriptome Data. Horticulturae 2023, 9, 1262. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Q.; Gao, R.; Zhen, B.; Wang, Y.; Gong, X.; Zhang, F. Research progress on Physalis philadelphica. Agric. Prod. Process. 2020, 8, 72–76. [Google Scholar]
- Guo, P.; Zhang, B.; Hu, Z.; Zhou, S.; Wang, Y.; Xie, Q.; Chen, G. Anthocyanin accumulation and transcriptional regulation in purple flowering stalk (Brassica campestris L. var. purpurea Bailey). Plant Mol. Biol. 2023, 111, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Park, H.W.; Yeo, H.J.; Jung, D.H.; Jeon, K.S.; Kim, T.J.; Kim, J.K.; Park, S.U. Combined transcriptome and metabolome analysis and evaluation of antioxidant and antibacterial activities in white, pink, and violet flowers of Angelica gigas. Ind. Crops Prod. 2022, 188, 115605. [Google Scholar] [CrossRef]
- De Rosso, V.V.; Mercadante, A.Z. Identification and quantification of carotenoids, by HPLC-PDA-MS/MS, from Amazonian fruits. J. Agric. Food Chem. 2007, 55, 5062. [Google Scholar] [CrossRef] [PubMed]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Lei, J.; Liu, Y. Research progress on breeding of red-fleshed strawberries. J. Northeast Agric. Univ. 2012, 43, 5. [Google Scholar]
- Liang, Y.; Chen, X.; Chen, D.; Wang, S.; Li, Y.; Huang, C. Research progress on MYB transcription factors regulating plant anthocyanin biosynthesis. Jiangsu Agric. Sci. 2022, 50, 10. [Google Scholar]
- Griesser, M.; Hoffmann, T.; Bellido, M.L.; Rosati, C.; Fink, B.; Kurtzer, R.; Aharoni, A.; Munoz-Blanco, J.; Schwab, W. Redirection of flavonoid biosynthesis through the down-regulation of an anthocyanidin glucosyltransferase in ripening strawberry fruit. Plant Physiol. 2008, 146, 1528–1539. [Google Scholar] [CrossRef]
- Zhang, N.; Hu, Z.; Chen, X.; Hou, X.; Li, Y.; Chen, G. Analysis of the anthocyanin metabolic pathway in plants and establishment of a regulatory model. Chin. J. Biotechnol. 2008, 28, 9. [Google Scholar]
- Zhou, H.; Lin-Wang, K.; Wang, H.; Gu, C.; Dare, A.P.; Espley, R.V.; He, H.; Allan, A.C.; Han, Y. Molecular genetics of blood-fleshed peach reveals activation of anthocyanin biosynthesis by NAC transcription factors. Plant J. 2015, 82, 105–121. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene IDs | Forward Primers (5′-3′) | Reverse Primers (5′-3′) |
|---|---|---|
| pf05G124640 | GCTTCTTTGTCTTGTCCGTTGTCTG | TGCCATTGAGACTTGCCGACAG |
| pf09G224140 | GCTTGGCTAGGAGTGGCATAAAGG | TCGTGGGTACTTGAGGTCCTTCATC |
| pf05G137060 | GTGTGGTGGTGGCGGATATGC | CTGTCATCAGTTCCACGACCATGTC |
| pf01G020090 | GAGTTGGATAGGCTTTCGGCTGAG | AATGGCTCCCTCAAATGGCTGATG |
| pf05G120860 | CTTGGAACCTTATGACCACCTCTGC | TTGCCATCATCAGGAAGAGCCATG |
| pf06G177100 | GCCAGTTTGACCACCCTCATTCTC | CAGGGAGGTTCAGCAGGAATAGAAG |
| pf06G169640 | ACATGGTACGGCAAATCATCCTCAG | CTCGTGGTCTCATTGGTCTGGTTG |
| pf09G237080 | ATTGGTGTCATCTTTGCAGCTAGGG | GTGACGGCGAGTAGGACACTAGG |
| pf05G143230 | TGGAGCCAGTGGTCGAAGGTC | ATCCAAGTGCGATGTCCAAGTATCC |
| pf01G016330 | GCTGTTGTGTCCGAGAACGAAGAG | GTCCCCATCAAGTAGTGCAAACCC |
| pf11G267170 | AAGCCAGAATGAGCATGAGCAGAG | AGCCAGTGTCACCATCAGCAATG |
| pf06G179690 | AACGGATTGCCCTCGACTGAAAC | AGCCTCAATACCCTTTGCCAACG |
| Pp.GAPDH | TGTGGGTGTCAACGAGAAGGAATAC | ATAAGACCCTCCACAATGCCAAACC |
| Flowering Stages | Anthocyanin Content (nmol/g) |
|---|---|
| Tieba initial flowering stage (AS2) | 5.59 b ± 0.48 |
| Qingjin initial flowering stage (BS2) | 57.90 a ± 0.32 |
| Tieba full flowering stage (AS3) | 11.72 b ± 1.27 |
| Qingjin full flowering stage (BS3) | 50.69 a ± 0.09 |
| Flowering Stages | Carotenoid Content (mg/g) |
|---|---|
| Tieba initial flowering stage (AS2) | 1.45 c ± 0.03 |
| Qingjin initial flowering stage (BS2) | 2.84 a ± 0.06 |
| Tieba full flowering stage (AS3) | 1.40 c ± 0.10 |
| Qingjin full flowering stage (BS3) | 2.01 b ± 0.09 |
| Flowering Stages | Delphinidin (μg/g) | Cyanidin (μg/g) | Pelargonidin (μg/g) | Malvidin (μg/g) |
|---|---|---|---|---|
| Tieba initial flowering stage (AS2) | Below detection limit | Below detection limit | Below detection limit | Below detection limit |
| Qingjin initial flowering stage (BS2) | 14.04 a ± 0.01 | 1.48 a ± 0.04 | Below detection limit | 1.20 a ± 0.02 |
| Tieba full flowering stage (AS3) | Below detection limit | Below detection limit | Below detection limit | Below detection limit |
| Qingjin full flowering stage (BS3) | 13.57 a ± 0.04 | 1.42 a ± 0.03 | Below detection limit | 0.88 b ± 0.11 |
| Flowering Stages | β-Carotene Content (μg/g) |
|---|---|
| Tieba initial flowering stage (AS2) | 83.5198 b ± 0.02 |
| Tieba full flowering stage (AS3) | 108.0468 a ± 0.03 |
| Qingjin initial flowering stage (BS2) | 45.6094 c ± 0.03 |
| Qingjin full flowering stage (BS3) | 110.8920 a ± 0.02 |
| Group | Gene Name | Description | Pathway |
|---|---|---|---|
| P. philadelphica Qingjin | pf09G224140 | Anthocyanin synthase | Flavonoid |
| pf05G124640 | Dihydroflavonol 4-reductase | ||
| pf05G125540 | Flavanone 3-hydroxylase | ||
| pf08G220750 | Flavonol synthase/Flavanone 3-hydroxylase | ||
| pf07G186580 | Chalcone-flavonoid isomerase 3 subtype X1 | ||
| pf09G234590 | Bright blue pigment dioxygenase | ||
| pf08G220770 | Flavonol synthase/Flavanone 3-hydroxylase | ||
| pf01G020090 | Anthocyanin 3-O-glucosyltransferase | Flavonol | |
| pf02G043370 | β-carotene hydroxylase | Carotenoid | |
| pf03G054950 | Decatriol dehydrogenase | ||
| pf09G235870 | Abscisic acid 8′-hydroxylase | ||
| pf09G237080 | Abscisic acid 8′-hydroxylase | ||
| pf12G300180 | Decatriol dehydrogenase | ||
| pf10G255070 | Isoprenol dehydrogenase | ||
| pf09G244090 | Isoprenol/Carverol dehydrogenase | ||
| pf04G094750 | Aflatoxin dehydrogenase-like subtype | ||
| pf09G244170 | Carverol dehydrogenase | ||
| pf10G253440 | Carotene ε-monooxygenase | ||
| P. philadelphica Tieba | pf07G186580 | Chalcone flavonoid isomerase 3 subtype | Flavonoid |
| pf05G125540 | Flavanone 3-hydroxylase | ||
| pf08G220770 | Flavonol synthase/Flavanone 3-hydroxylase | ||
| pf08G220750 | Flavonol synthase/Flavanone 3-hydroxylase | ||
| pf09G224140 | Anthocyanin synthase | ||
| pf01G020090 | Anthocyanin 3-O-glucosyltransferase | Flavonol | |
| pf11G295430 | Anthocyanin 3-O-glucosyltransferase 7 | ||
| pf09G237080 | Abscisic acid 8′-hydroxylase | Carotenoid | |
| pf06G177100 | Zeaxanthin epoxide enzyme | ||
| pf06G169640 | (9-cis-epoxycarotenoid dioxygenase | ||
| pf02G043370 | β-carotene hydroxylase | ||
| pf09G235870 | Abscisic acid 8′-hydroxylase | ||
| pf12G298210 | Monooxygenase | ||
| pf12G301650 | Decaisotriol dehydrogenase-like | ||
| pf04G091760 | Abscisic acid 8′-hydroxylase | ||
| pf11G280920 | Short-chain dehydrogenase reductase | ||
| pf03G054950 | Decaisotriol dehydrogenase-like | ||
| pf08G205980 | Shedding aldehyde oxidase-like subtype | ||
| pf10G253440 | Carotene ε monooxygenase | ||
| pf08G205960 | Indole-3-acetaldehyde oxidase | ||
| pf11G283010 | β-carotene hydroxylase | ||
| pf12G300180 | Decaisotriol dehydrogenase-like | ||
| pf06G161300 | Short-chain dehydrogenase reductase 4 |
| Comparison Groups | Gene Name | Description | Regulation | Transcription Factors |
|---|---|---|---|---|
| AS3 vs. AS2 | pf05G125540 | Hesperetin 3-hydroxylase | Down | Dof |
| pf07G186580 | Chalcone-flavonoid isomerase | Down | NAC | |
| pf08G220750 | Flavonol synthase/flavanone 3-hydroxylase | Down | STAT | |
| pf08G220770 | Flavonol synthase/flavanone 3-hydroxylase | Down | STAT | |
| pf09G224140 | Anthocyanin synthase | Down | STAT | |
| BS3 vs. BS2 | pf05G124640 | Dihydroflavonol 4-reductase | Down | GeBP |
| pf05G125540 | Flavanone 3-hydroxylase | Down | Dof | |
| pf07G186580 | Chalcone-flavonoid isomerase | Down | NAC | |
| pf08G220750 | Flavonol synthase/flavanone 3-hydroxylase | Down | STAT | |
| pf08G220770 | Flavonol synthase/flavanone 3-hydroxylase | Down | STAT | |
| pf09G224140 | Anthocyanin synthase | Down | STAT | |
| pf09G234590 | Biliverdin dioxygenase | Down | STAT | |
| AS3 vs. BS3 | pf05G124640 | Dihydroflavonol 4-reductase | Down | GeBP |
| pf09G224140 | Anthocyanin synthase | Down | STAT | |
| AS2 vs. BS2 | pf05G124640 | Dihydroflavonol 4-reductase | Down | GeBP |
| pf09G224140 | Anthocyanin synthase | Down | STAT |
| Comparison Groups | Gene Name | Description | Regulation | Transcription Factors |
|---|---|---|---|---|
| AS3 vs. AS2 | pf01G020090 | Anthocyanin 3-O-glucosyltransferase | Down | HSF |
| pf11G295430 | Anthocyanin 3-O-glucosyltransferase | Down | GRAS | |
| BS3 vs. BS2 | pf01G020090 | Anthocyanin 3-O-glucosyltransferase | Down | HSF |
| AS3 vs. BS3 | pf01G020090 | Anthocyanin 3-O-glucosyltransferase | Down | HSF |
| AS2 vs. BS2 | pf01G020090 | Anthocyanin 3-O-glucosyltransferase | Down | HSF |
| Comparison Groups | Gene Name | Description | Regulation | Transcription Factors |
|---|---|---|---|---|
| AS3 vs. AS2 | pf04G091760 | Abscisic acid 8′-hydroxylase | Up | G2-like |
| pf06G161300 | Short-chain dehydrogenase reductase | Down | NAC | |
| pf08G205960 | Indole-3-acetaldehyde oxidase | Down | LBD | |
| pf09G235870 | Abscisic acid 8′-hydroxylase | Up | G2-like | |
| pf09G237080 | Abscisic acid 8′-hydroxylase | Up | G2-like | |
| pf10G253440 | Carotene ε-monooxygenase | Down | bHLH | |
| pf11G280920 | Short-chain dehydrogenase reductase | Up | NAC | |
| BS3 vs. BS2 | pf04G094750 | Aflatoxin dehydrogenase | Down | NAC |
| pf09G235870 | Abscisic acid 8′-hydroxylase | Up | G2-like | |
| pf09G237080 | Abscisic acid 8′-hydroxylase | Up | G2-like | |
| pf09G244090 | Isopentenol dehydrogenase | Down | bHLH | |
| pf09G244170 | Isopentenol dehydrogenase | Down | NAC | |
| pf10G253440 | Carotene ε-monooxygenase | Down | bHLH | |
| pf10G255070 | Divinyl ether synthase | Down | NAC | |
| AS3 vs. BS3 | pf10G255070 | Divinyl ether synthase | Up | NAC |
| AS2 vs. BS2 | pf09G237080 | Abscisic acid 8′-hydroxylase | Down | G2-like |
| Gene_Name | FPKM. A_S2_1 | FPKM. A_S2_2 | FPKM. A_S2_3 | FPKM. A_S3_1 | FPKM. A_S3_2 | FPKM. A_S3_3 |
|---|---|---|---|---|---|---|
| pf01G016330 | 137.83 | 95.08 | 137.20 | 15.39 | 12.22 | 22.80 |
| pf01G020090 | 21.22 | 15.23 | 15.81 | 0.52 | 0.24 | 0.10 |
| pf05G120860 | 5.24 | 6.28 | 4.51 | 25.08 | 17.11 | 25.89 |
| pf05G124640 | 0.12 | 0.16 | 0.21 | 0.34 | 0.10 | 0.26 |
| pf05G137060 | 17.29 | 7.12 | 12.47 | 0.32 | 0.27 | 0.06 |
| pf05G143230 | 97.28 | 58.98 | 89.15 | 2.93 | 1.03 | 1.52 |
| pf06G169640 | 15.47 | 7.32 | 19.59 | 56.88 | 72.64 | 69.55 |
| pf06G177100 | 4.90 | 4.75 | 4.09 | 48.37 | 46.04 | 41.24 |
| pf06G179690 | 24.93 | 20.92 | 21.57 | 6.69 | 7.97 | 8.74 |
| pf09G224140 | 4.25 | 3.18 | 4.21 | 1.58 | 2.03 | 1.02 |
| pf09G237080 | 0.57 | 1.03 | 0.79 | 42.06 | 33.00 | 33.05 |
| pf11G267170 | 57.51 | 58.49 | 41.90 | 9.35 | 18.56 | 31.15 |
| Gene_Name | FPKM. B_S2_1 | FPKM. B_S2_2 | FPKM. B_S2_3 | FPKM. B_S3_1 | FPKM. B_S3_2 | FPKM. B_S3_3 |
|---|---|---|---|---|---|---|
| pf01G016330 | 289.09 | 264.30 | 220.95 | 23.24 | 22.72 | 30.06 |
| pf01G020090 | 57.01 | 58.46 | 38.68 | 3.03 | 1.39 | 5.39 |
| pf05G120860 | 15.91 | 15.51 | 28.10 | 17.91 | 17.05 | 35.35 |
| pf05G124640 | 293.83 | 295.91 | 229.43 | 26.21 | 15.19 | 36.54 |
| pf05G137060 | 175.92 | 190.60 | 107.30 | 15.54 | 4.24 | 16.09 |
| pf05G143230 | 105.13 | 86.95 | 72.13 | 2.95 | 2.59 | 4.28 |
| pf06G169640 | 53.47 | 36.49 | 56.95 | 46.96 | 43.04 | 76.05 |
| pf06G177100 | 10.70 | 21.88 | 29.88 | 23.83 | 34.22 | 32.77 |
| pf06G179690 | 25.27 | 28.91 | 22.19 | 3.62 | 4.62 | 6.03 |
| pf09G224140 | 100.57 | 102.84 | 87.72 | 9.12 | 7.99 | 9.99 |
| pf09G237080 | 2.54 | 2.13 | 11.36 | 14.99 | 10.24 | 27.84 |
| pf11G267170 | 66.09 | 54.08 | 47.89 | 6.90 | 9.85 | 6.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, H.; Zhao, W.; Tian, S.; Wang, D.; Wu, H.; Wang, C.; Zhu, J.; Li, N.; Zhu, X.; Mu, S.; et al. Exploring the Mechanisms Underlying Petal Pigmentation Differences in Two Cultivars of Physalis philadelphica Based on HPLC and NGS. Horticulturae 2024, 10, 507. https://doi.org/10.3390/horticulturae10050507
Qiao H, Zhao W, Tian S, Wang D, Wu H, Wang C, Zhu J, Li N, Zhu X, Mu S, et al. Exploring the Mechanisms Underlying Petal Pigmentation Differences in Two Cultivars of Physalis philadelphica Based on HPLC and NGS. Horticulturae. 2024; 10(5):507. https://doi.org/10.3390/horticulturae10050507
Chicago/Turabian StyleQiao, Hongyu, Wennan Zhao, Song Tian, Da Wang, Haiyan Wu, Chenyu Wang, Jiaming Zhu, Nan Li, Xu Zhu, Shujing Mu, and et al. 2024. "Exploring the Mechanisms Underlying Petal Pigmentation Differences in Two Cultivars of Physalis philadelphica Based on HPLC and NGS" Horticulturae 10, no. 5: 507. https://doi.org/10.3390/horticulturae10050507
APA StyleQiao, H., Zhao, W., Tian, S., Wang, D., Wu, H., Wang, C., Zhu, J., Li, N., Zhu, X., Mu, S., Zhang, J., & He, H. (2024). Exploring the Mechanisms Underlying Petal Pigmentation Differences in Two Cultivars of Physalis philadelphica Based on HPLC and NGS. Horticulturae, 10(5), 507. https://doi.org/10.3390/horticulturae10050507