
Longevity and Potential Mechanisms of Fenpropathrin Resistance in Asian Citrus Psyllid, Diaphorina citri Kuwayama

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects, Plants, and Chemicals
2.2. Stability of Fenpropathrin Resistance
2.3. Nucleic Acid Extraction and cDNA Synthesis
2.4. Real-Time Quantitative PCR for Expression
2.5. Statistical Analysis
3. Results
3.1. Stability of Fenpropathrin Resistance among D. citri Populations
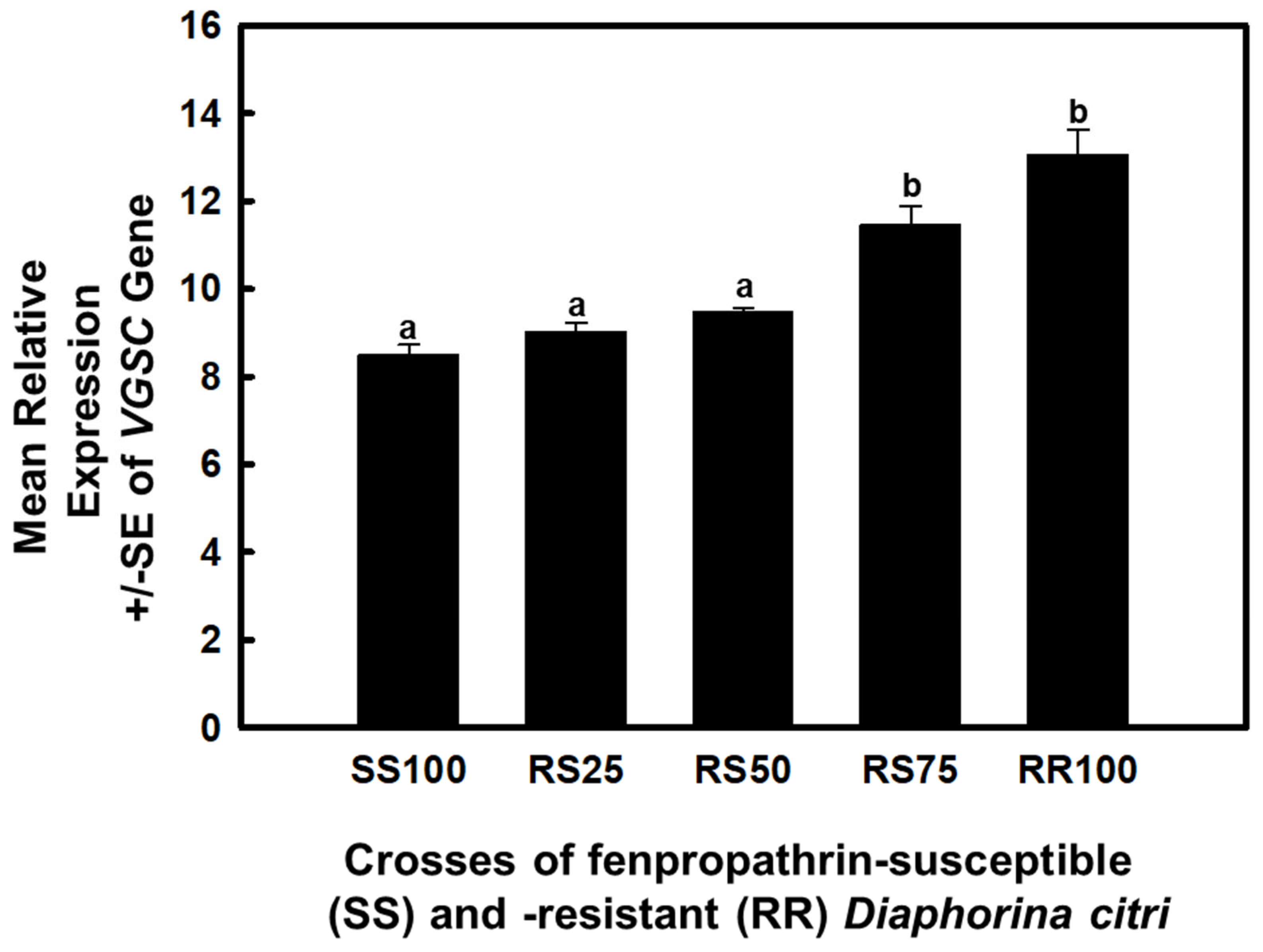
3.2. Expression of VGSC in Insecticide-Resistant, Susceptible and Crossed Populations
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bove, J.M. Huanglongbing: A Destructive, Newly-Emerging, Century-Old Disease of Citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and Management of Asian Citrus Psyllid, Vector of the Huanglongbing Pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed]
- Kanga, L.H.B.; Eason, J.; Haseeb, M.; Qureshi, J.; Stansly, P. Monitoring for Insecticide Resistance in Asian Citrus Psyllid (Hemiptera: Psyllidae) Populations in Florida. J. Econ. Entomol. 2016, 109, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Mann, R.S.; Rogers, M.E.; Stelinski, L.L. Insecticide Resistance in Field Populations of Asian Citrus Psyllid in Florida. Pest Manag. Sci. 2011, 67, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Mo, X.; Rizvi, S.A.H.; Li, C.; Zeng, X. Detection and Biochemical Characterization of Insecticide Resistance in Field Populations of Asian Citrus Psyllid in Guangdong of China. Sci. Rep. 2018, 8, 12587. [Google Scholar] [CrossRef] [PubMed]
- Coy, M.R.; Bin, L.; Stelinski, L.L. Reversal of Insecticide Resistance in Florida Populations of Diaphorina citri (Hemiptera: Liviidae). Fla. Entomol. 2016, 99, 26–32. [Google Scholar] [CrossRef]
- Vázquez-García, M.; Velázquez-Monreal, J.; Medina-Urrutia, V.M.; Cruz-Vargas, C.D.J.; Sandoval-Salazar, M.; Virgen-Calleros, G.; Torres-Morán, J.P. Insecticide Resistance in Adult Diaphorina citri Kuwayama1 from Lime Orchards in Central West Mexico. Southwest. Entomol. 2013, 38, 579–596. [Google Scholar] [CrossRef]
- Pardo, S.; Martínez, A.M.; Figueroa, J.I.; Chavarrieta, J.M.; Viñuela, E.; Rebollar-Alviter, Á.; Miranda, M.A.; Valle, J.; Pineda, S. Insecticide Resistance of Adults and Nymphs of Asian Citrus Psyllid Populations from Apatzingán Valley, Mexico. Pest Manag. Sci. 2018, 74, 135–140. [Google Scholar] [CrossRef]
- Naeem, A.; Freed, S.; Jin, F.L.; Akmal, M.; Mehmood, M. Monitoring of Insecticide Resistance in Diaphorina citri Kuwayama (Hemiptera: Psyllidae) from Citrus Groves of Punjab, Pakistan. Crop Prot. 2016, 86, 62–68. [Google Scholar] [CrossRef]
- Chen, X.D.; Sanada-Morimura, S.; Yanagi, S.I.; Nakasuji, F. Rapid Recovery of Susceptibility under Harsh Environmental Conditions in Fenvalerate-Resistant Strains of the Diamondback Moth, Plutella xylostella (Lepidoptera: Yponomeutidae). Appl. Entomol. Zool. 2006, 41, 641–650. [Google Scholar] [CrossRef]
- Afzal, M.B.S.; Shad, S.A.; Abbas, N.; Ayyaz, M.; Walker, W.B. Cross-Resistance, the Stability of Acetamiprid Resistance and Its Effect on the Biological Parameters of Cotton Mealybug, Phenacoccus solenopsis (Homoptera: Pseudococcidae), in Pakistan. Pest Manag. Sci. 2015, 71, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Saddiq, B.; Shahzad Afzal, M.B.; Shad, S.A. Studies on Genetics, Stability and Possible Mechanism of Deltamethrin Resistance in Phenacoccus solenopsis Tinsley (Homoptera: Pseudococcidae) from Pakistan. J. Genet. 2016, 95, 1009–1016. [Google Scholar] [CrossRef]
- Bielza, P.; Quinto, V.; Grávalos, C.; Fernández, E.; Abellán, J.; Contreras, J. Stability of Spinosad Resistance in Frankliniella occidentalis (Pergande) under Laboratory Conditions. Bull. Entomol. Res. 2008, 98, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.C.; Smith, L.B.; Silva, J.J.; Fan, Y.; Sun, H.; Scott, J.G. Fitness Studies of Insecticide Resistant Strains: Lessons Learned and Future Directions. Pest Manag. Sci. 2021, 77, 3847–3856. [Google Scholar] [CrossRef] [PubMed]
- Basit, M.; Sayyed, A.H.; Saleem, M.A.; Saeed, S. Cross-Resistance, Inheritance and Stability of Resistance to Acetamiprid in Cotton Whitefly, Bemisia tabaci Genn (Hemiptera: Aleyrodidae). Crop Prot. 2011, 30, 705–712. [Google Scholar] [CrossRef]
- Inoue, K. The Change of Susceptibility of Mite Population to Dicofol and Genetic Analysis of Dicofol-Resistance in the Citrus Red Mite, Panonychus citri (McG). J. Pestic. Sci. 1979, 4, 337–344. [Google Scholar] [CrossRef]
- Needham, P.H.; Sawicki, R.M. Diagnosis of Resistance to Organophosphorus Insecticides in Myzus persicae (Sulz). Nature 1971, 230, 125–126. [Google Scholar] [CrossRef]
- Keiding, J. Resistance in the Housefly in Denmark and Elsewhere Kriding Pesticide Management and Insecticide Resistance. In Pesticide Management and Insecticide Resistance; Watson, D.L., Brown, A.W.A., Eds.; Academic Press: New York, NY, USA, 1977; pp. 261–302. [Google Scholar]
- Scott, J.G.; Georghiou, G.P. Mechanisms Responsible for High Levels of Permethrin Resistance in the House Fly. Pestic. Sci. 1986, 17, 195–206. [Google Scholar] [CrossRef]
- Gajendiran, A.; Abraham, J. An Overview of Pyrethroid Insecticides. Front. Biol. 2018, 13, 79–90. [Google Scholar] [CrossRef]
- Catterall, W.A. From Ionic Currents to Molecular Mechanisms: The Structure and Function of Voltage-Gated Sodium Channels. Neuron 2000, 26, 13–25. [Google Scholar] [CrossRef]
- Dong, K.; Du, Y.; Rinkevich, F.; Nomura, Y.; Xu, P.; Wang, L.; Silver, K.; Zhorov, B.S. Molecular Biology of Insect Sodium Channels and Pyrethroid Resistance. Insect Biochem. Mol. Biol. 2014, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Coy, M.R.; Wang, J.J.; Stelinski, L.L. Characterization of the Voltage-Gated Sodium Channel of the Asian Citrus Psyllid, Diaphorina citri. Insect Sci. 2017, 24, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.S.; Martinez-Torres, D.; Hick, C.A.; Devonshire, A.L. Identification of Mutations in the Housefly Para-Type Sodium Channel Gene Associated with Knockdown Resistance (Kdr) to Pyrethroid Insecticides. Mol. Gen. Genet. 1996, 252, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Soderlund, D.M.; Knipple, D.C. The Molecular Biology of Knockdown Resistance to Pyrethroid Insecticides. Insect Biochem. Mol. Biol. 2003, 33, 563–577. [Google Scholar] [CrossRef] [PubMed]
- Narahashi, T. Neuroreceptors and Ion Channels as the Basis for Drug Action: Past, Present, and Future. J. Pharmacol. Exp. Ther. 2000, 294, 1–26. [Google Scholar] [PubMed]
- Chen, X.D.; Sandoval-Mojica, A.F.; Bonilla, S.I.; Ebert, T.A.; Gossett, H.; Pelz-Stelinski, K.S.; Stelinski, L.L. Fenpropathrin Resistance in Asian Citrus Psyllid, Diaphorina citri Kuwayama: Risk Assessment and Changes in Expression of CYP and GST Genes Associated with Resistance. Int. J. Pest Manag. 2023, 69, 54–63. [Google Scholar] [CrossRef]
- Chen, X.D.; Stelinski, L.L. Resistance Management for Asian Citrus Psyllid, Diaphorina citri Kuwayama, in Florida. Insects 2017, 8, 103. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- SAS Institute (2002–2012) User’s Manual, Version 9.4; SAS Institute: Cary, NC, USA, 2016.
- Robertson, J.L.; Preisler, H.K. Pesticide Bioassays with Arthropods; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar] [CrossRef]
- Liu, Y.H.; Tsai, J.H. Effects of temperature and life table parameters of the Asian citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae). Ann. Appl. Biol. 2000, 137, 201–206. [Google Scholar] [CrossRef]
- Robb, K.L. Analysis of Frankliniella occidentalis (Pergande) as a Pest of Floricultural Crops in California 405 Greenhouses. Ph.D. Thesis, University of California, Riverside, CA, USA, 1989. [Google Scholar]
- Kontsedalov, S.; Weintraub, P.G.; Horowitz, A.R.; Ishaaya, I. Effects of Insecticides on Immature and Adult Western Flower Thrips (Thysanoptera: Thripidae) in Israel. J. Econ. Entomol. 1998, 91, 1067–1071. [Google Scholar] [CrossRef]
- Chen, X.D.; Ebert, T.A.; Pelz-Stelinski, K.S.; Stelinski, L.L. Fitness Costs Associated with Thiamethoxam and Imidacloprid Resistance in Three Field Populations of Diaphorina citri (Hemiptera: Liviidae) from Florida. Bull. Entomol. Res. 2020, 110, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Roush, R.T.; Croft, B.A. Experimental population genetics and ecological studies of pesticide resistance in insects and mites. In Pesticide Resistance Strategies and Tactics for Management; National Academy Press: Washington, DC, USA, 1986; pp. 257–270. [Google Scholar]
- Denholm, I.; Rowland, M.W. Tactics for Managing Pesticide Resistance in Arthropods: Theory and Practice. Annu. Rev. Entomol. 1992, 37, 91–112. [Google Scholar] [CrossRef] [PubMed]
- Nkya, T.E.; Akhouayri, I.; Kisinza, W.; David, J.P. Impact of Environment on Mosquito Response to Pyrethroid Insecticides: Facts, Evidences and Prospects. Insect Biochem. Mol. Biol. 2013, 43, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Amichot, M.; Castella, C.; Cuany, A.; Berge, J.B.; Pauron, D. Target Modification as a Molecular Mechanism of Pyrethroid Resistance in Drosophila melanogaster. Pestic. Biochem. Physiol. 1992, 44, 183–190. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, H.; Zhang, L.; Liu, N. Sodium Channel Gene Expression Associated with Pyrethroid Resistant House Flies and German Cockroaches. Gene 2006, 379, 62–67. [Google Scholar] [CrossRef]
- Sonoda, S.; Igaki, C.; Ashfaq, M.; Tsumuki, H. Pyrethroid-Resistant Diamondback Moth Expresses Alternatively Spliced Sodium Channel Transcripts with and without T929I Mutation. Insect Biochem. Mol. Biol. 2006, 36, 904–910. [Google Scholar] [CrossRef]

{kind=link}
| Gene | Forward/Reverse | Sequence (Forward/Reverse) | Reference |
|---|---|---|---|
| F | AGCGGAAAATTACACGTGGG | ||
| VGSC | R | CGGATACCTTTGGCCCCTTT | [23] |
| F | CCCTGGACTTTGAACAGGAA | ||
| Actin | R | CTCGTGGATACCGCAAGATT | [27] |
| Line | Month | Slope ± SE | LC50 (ng/µL) (95 % Confidence Interval) | LC90 (ng/µL) (95 % Confidence Interval) |
|---|---|---|---|---|
| 100 RR + 00 SS | 0 | 0.78 ± 0.17 | 18.57 (3.31–161.99) | 912.09 (107.592–3966) |
| 2 | 0.69 ± 0.15 | 10.70 (1.52–134.51) | 781.53 (76.23–547546) | |
| 4 | 0.82 ± 0.12 | 11.90 (5.01–30.69) | 436.53 (131.87–3111) | |
| 6 | 0.82 ± 0.13 | 14.75 (5.324–3.67) | 532.98 (142.90–5757.00) | |
| 8 | 0.69 ± 0.12 | 5.02 (1.67–15.54) | 249.68 (61.72–3256.98) | |
| 75 RR + 25 SS | 2 | 0.88 ± 0.31 | 4.84 (2.21–11.13) | 570.42 (174.23–3169) |
| 4 | 0.81 ± 0.14 | 6.54 (1.50–35.52) | 329.94 (54.15–18627) | |
| 6 | 0.61 ± 0.09 | 7.50 (3.06–19.77) | 389.60 (59.00–1082) | |
| 8 | 0.89 ± 0.09 | 3.31 (1.42–7.54) | 91.38 (33.59–418) | |
| 50 RR + 50 SS | 2 | – | – | – |
| 4 | 0.60 ± 0.20 | 5.57 (1.18–35.54) | 303.57 (46.09–22548) | |
| 6 | 0.51 ± 0.07 | 2.51 (0.81–8.43) | 794.56 (144.00–13207) | |
| 8 | 0.88 ± 0.13 | 1.99 (0.80–5.12) | 102.19 (30.70–671.50) | |
| 25 RR + 75 SS | 2 | 0.42 ± 0.06 | 0.91 (0.30–2.63) | 314.73 (70.41–3281) |
| 4 | 0.56 ± 0.09 | 0.85 (0.15–5.34) | 164.11 (19.18–16330) | |
| 6 | 0.67 ± 0.09 | 0.81 (0.33–2.00) | 33.76 (13.13–235.75) | |
| 8 | 0.89 ± 0.13 | 0.77 (0.42–1.42) | 64.23 (27.35–196.74) | |
| 00 RR + 100 SS | 0 | 0.89 ± 0.12 | 0.35 (0.16–0.78) | 9.73 (3.803–8.46) |
| 2 | 1.11 ± 0.14 | 0.40 (0.22–0.77) | 5.71 (2.75–16.59) | |
| 4 | 0.76 ± 0.16 | 0.40 (0.06–3.62) | 19.36 (2.71–475.16) | |
| 6 | 0.70 ± 0.10 | 0.74 (0.29–1.90) | 48.70 (14.41–317.51) | |
| 8 | 0.79 ± 0.11 | 0.43 (0.27–0.67) | 16.67 (9.84–31.78) |
| Line | Month | RR50 | RR90 |
|---|---|---|---|
| 100 RR + 00 SS | 0 | 60.14 (16.99–212.83) | 951.00 (82.00–10941.00) |
| 2 | 34.00 (11.73–100.09) | 80.32 (13.00–472.00) | |
| 4 | 32.85 (13.63–79.14) | 31.07 (7.3–7131.01) | |
| 6 | 19.96 (5.64–70.63) | 10.95 (1.37–87.25) | |
| 8 | 10.83 (2.91–40.34) | 20.14 (3.16–128.36) | |
| 75 RR + 25 SS | 2 | 13.92 (4.56–42.48) | 58.60 (9.61–357.47) |
| 4 | 18.06 (7.38–44.17) | 23.48 (5.37–102.64) | |
| 6 | 10.15 (2.78–37.09) | 35.20 (0.93–69.70) | |
| 8 | 7.61 (2.20–26.32) | 5.48 (1.04–28.81) | |
| 50 RR + 50 SS | 2 | – | – |
| 4 | 15.85 (6.46–38.95) | 22.57 (5.14–99.17) | |
| 6 | 3.39 (0.78–14.75) | 16.32 (1.20–221.31) | |
| 8 | 2.69 (0.88–8.27) | 2.10 (0.34–12.84) | |
| 25 RR + 75 SS | 2 | 3.89 (0.84–18.09) | 30.45 (3.69–251.60) |
| 4 | 2.35 (0.77–7.20) | 11.68 (1.75–77.59) | |
| 6 | 1.15 (0.2–83.94) | 1.32 (0.16–11.15) | |
| 8 | 1.97 (0.51–7.57) | 1.41 (0.22–9.16) | |
| 00 RR + 100 SS | 0 | 1 (0.34–2.98) | 1 (0.19–5.47) |
| 2 | 1 (0.35–2.83) | 1 (0.18–5.47) | |
| 4 | 1 (0.40–2.47) | 1 (0.23–4.26) | |
| 6 | 1 (0.27–3.75) | 1 (0.11–9.23) | |
| 8 | 1 (0.22–4.55) | 1 (0.14–7.03) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Stockton, D.G.; Gill, T.A.; Gossett, H.; Qureshi, J.A.; Pelz-Stelinski, K.S.; Stelinski, L.L. Longevity and Potential Mechanisms of Fenpropathrin Resistance in Asian Citrus Psyllid, Diaphorina citri Kuwayama. Horticulturae 2024, 10, 448. https://doi.org/10.3390/horticulturae10050448
Chen X, Stockton DG, Gill TA, Gossett H, Qureshi JA, Pelz-Stelinski KS, Stelinski LL. Longevity and Potential Mechanisms of Fenpropathrin Resistance in Asian Citrus Psyllid, Diaphorina citri Kuwayama. Horticulturae. 2024; 10(5):448. https://doi.org/10.3390/horticulturae10050448
Chicago/Turabian StyleChen, Xuedong, Dara G. Stockton, Torrence A. Gill, Hunter Gossett, Jawwad A. Qureshi, Kirsten S. Pelz-Stelinski, and Lukasz L. Stelinski. 2024. "Longevity and Potential Mechanisms of Fenpropathrin Resistance in Asian Citrus Psyllid, Diaphorina citri Kuwayama" Horticulturae 10, no. 5: 448. https://doi.org/10.3390/horticulturae10050448
APA StyleChen, X., Stockton, D. G., Gill, T. A., Gossett, H., Qureshi, J. A., Pelz-Stelinski, K. S., & Stelinski, L. L. (2024). Longevity and Potential Mechanisms of Fenpropathrin Resistance in Asian Citrus Psyllid, Diaphorina citri Kuwayama. Horticulturae, 10(5), 448. https://doi.org/10.3390/horticulturae10050448