
Phenolic, Amino Acid, Mineral, and Vitamin Contents during Berry Development in ‘Italia’ and ‘Bronx Seedless’ Grape Cultivars





Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemical and Reagents
2.3. Determination of Phenolic Compounds from Grape Cultivars by HPLC
2.4. Determination of Amino Acid Profiling in Grape Cultivars by HPLC
2.5. Determination of Vitamin Profiling in Grape Cultivars by HPLC
2.6. Determination of Mineral Element Profiling in Grape Cultivars
2.7. Statistical Analysis
3. Results
3.1. Physico-Chemical Characteristics of Grape
3.2. Amino Acid Content
3.3. Phenolic Compounds
3.4. Minerals
3.5. Vitamins
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kaya, O.; Ates, F.; Kara, Z.; Turan, M.; Gutiérrez-Gamboa, G. Study of Primary and Secondary Metabolites of Stenospermocarpic, Parthenocarpic, and Seeded Raisin Varieties. Horticulturae 2022, 8, 1030. [Google Scholar] [CrossRef]
- Vilanova, M.; Genisheva, Z.; Bescansa, L.; Masa, A.; Oliveira, J.M. Changes in Free and Bound Fractions of Aroma Compounds of Four V. Vinifera Cultivars at the Last Ripening Stages. Phytochemistry 2012, 74, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.; Shrestha, P.M.; Smith, B.M. Nitrogen deficiency decreases the value of 3-isobutyl-2-methoxypyrazine in Cabernet Sauvignon grape berries. Food Chem. 2012, 134, 1506–1514. [Google Scholar]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Handbook of Enology, Volume 2: The Chemistry of Wine, Stabilization and Treatments, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Keskin, N.; Kaya, O.; Ates, F.; Turan, M.; Gutiérrez-Gamboa, G. Drying grapes after the application of different dipping solutions: Effects on hormones, minerals, vitamins, and antioxidant enzymes in Gök Üzüm (V. vinifera L.) raisins. Plants 2022, 11, 529. [Google Scholar] [CrossRef]
- Bordiga, M.; Travaglia, F.; Sacco, D.; Coïsson, J.D.; Arlorio, M. Characterisation of the Polyphenolic and Volatile Composition of European Elderberry (Sambucus nigra L.) Extracts. Plant Foods Hum. Nutr. 2011, 66, 29–36. [Google Scholar]
- Keller, M. Managing grapevines to optimize fruit development in a challenging environment: A climate change primer for viticulturists. Aust. J. Grape Wine Res. 2010, 16, 56–69. [Google Scholar] [CrossRef]
- Hannah, L.C.; Roane, M.K.; Hoyt, P. Characterization of two alcohol dehydrogenases from developing grape berries. Plant Physiol. 2006, 140, 979–988. [Google Scholar]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavor. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Kanellos, P.T.; Kaliora, A.C.; Gioxari, A.; Christopoulou, G.O.; Kalogeropoulos, N. Bioaccessibility of essential elements from mussels and grapes. Food Chem. 2018, 245, 490–497. [Google Scholar]
- Sarni-Manchado, P.; Cheynier, V.; Moutounet, M. Analysis and Role of Phenolic Compounds in Grapes, Musts, and Wines. In Fruit and Vegetable Flavour: Recent Advances and Future Prospects; Rouseff, R.L., da Fonseca, M.A.R.B., Eds.; American Chemical Society: Washington, DC, USA, 2005; pp. 309–340. [Google Scholar]
- Yilmaz, T.; Ates, F.; Turan, M.; Hatterman-Valenti, H.; Kaya, O. Dynamics of Sugars, Organic Acids, Hormones, and Antioxidants in Grape Varieties ‘Italia’ and ‘Bronx Seedless’ during Berry Development and Ripening. Horticulturae 2024, 10, 229. [Google Scholar] [CrossRef]
- He, F.; Liang, N.N.; Mu, L.; Pan, Q.H. Biosynthesis of Anthocyanins and Their Regulation in Colored Grapes. Molecules 2012, 17, 1190–1209. [Google Scholar] [CrossRef]
- Rolle, L.; Torchio, F.; Zeppa, G.; Gerbi, V. The role of some antioxidants in a pure Sangiovese must during fermentation and their evolution during different storage techniques. Food Chem. 2014, 147, 163–169. [Google Scholar]
- Hord, N.G.; Tang, Y.; Bryan, N.S. Food sources of nitrates and nitrites: The physiologic context for potential health benefits. Am. J. Clin. Nutr. 2009, 90, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth Stages of the Grapevine: Phenological growth stages of the grapevine (V. vinifera L. ssp. vinifera) Codes and descriptions according to the extended BBCH scale. Aust. J. Grape Wine Res. 1995, 1, 100–103. [Google Scholar] [CrossRef]
- Sagdic, O.; Ozturk, I.; Ozkan, G.; Yetim, H.; Ekici, L.; Yilmaz, M.T. RP-HPLC–DAD analysis of phenolic compounds in pomace extracts from five grape cultivars: Evaluation of their antioxidant, antiradical, and antifungal activities in orange and apple juices. Food Chem. 2011, 126, 1749–1758. [Google Scholar] [CrossRef] [PubMed]
- Barrado, E.; Rodriguez, J.A.; Castrillejo, Y. Determination of primary amino acids in wines by high performance liquid magneto-chromatography. Talanta 2009, 78, 672–675. [Google Scholar] [CrossRef]
- Kumar, S.S.; Samydurai, P.; Ramakrishnan, R.; Nagarajan, N. Polyphenols vitamin-E estimation and in vitro antioxidant activity of Adiantum capillus-veneris. Int. J. Innov. Pharm. Sci. 2013, 4, 258–262. [Google Scholar]
- Mozumder, N.R.; Akhter, M.J.; Khatun, A.A.; Rokibuzzaman, M.; Akhtaruzzaman, M. Estimation of water-soluble vitamin B-complex in selected leafy and non-leafy vegetables by HPLC method. Orient. J. Chem. 2019, 35, 1344. [Google Scholar] [CrossRef]
- AOAC. Official Method Analysis 975.03. Metals in Plants and Pets Food Atomic Absorption Spectrophotometric Method. 2005. Available online: https://scirp.org/reference/referencespapers.aspx?referenceid=2783175 (accessed on 15 July 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 12 August 2023).
- Murillo-Peña, R.; Garde-Cerdán, T.; Martínez-Vidaurre, J.M. Evaluation of foliar applications of urea at three concentrations on grape amino acids composition. J. Sci. Food Agric. 2023, 103, 4826–4837. [Google Scholar] [CrossRef] [PubMed]
- Moukarzel, R.; Parker, A.K.; Schelezki, O.J.; Gregan, S.M.; Jordan, B. Bunch microclimate influences amino acids and phenolic profiles of Pinot noir grape berries. Front. Plant Sci. 2023, 14, 1162062. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, C.; Gao, Z.; Yao, Y.; Kang, H.; Du, Y. VvPL15 Is the Core Member of the Pectate Lyase Gene Family Involved in Grape Berries Ripening and Softening. Int. J. Mol. Sci. 2023, 24, 9318. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, M.; Fujishima, H.; Chijiwa, H. Evaluation of table grape genetic resources for sugar, organic acid, and amino acid composition of berries. Euphytica 2010, 174, 1–13. [Google Scholar] [CrossRef]
- Esparza, I.; Moler, J.A.; Arteta, M.; Jiménez-Moreno, N.; Ancín-Azpilicueta, C. Phenolic composition of grape stems from different Spanish varieties and vintages. Biomolecules 2021, 11, 1221. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuw, R.; Kevers, C.; Pincemail, J.; Defraigne, J.O.; Dommes, J. Antioxidant capacity and phenolic composition of red wines from various grape varieties: Specificity of Pinot Noir. J. Food Compos. Anal. 2014, 36, 40–50. [Google Scholar] [CrossRef]
- Gómez-Plaza, E.; Bautista-Ortín, A.B.; Ruiz-García, Y.; Fernández-Fernández, J.I.; Gil-Muñoz, R. Effect of elicitors on the evolution of grape phenolic compounds during the ripening period. J. Sci. Food Agric. 2017, 97, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Magariño, S.; González-San José, M.L. Effect of ripening stage of grapes on the low molecular weight phenolic compounds of red wines. Eur. Food Res. Technol. 2005, 220, 597–606. [Google Scholar] [CrossRef]
- Askari-Khorasgani, O.; Pessarakli, M. Grapevine selection for improving nutrient content and composition and the associated quality indices—A review. J. Plant Nutr. 2019, 42, 2176–2187. [Google Scholar] [CrossRef]
- Sweetman, C.; Deluc, L.G.; Cramer, G.R.; Ford, C.M.; Soole, K.L. Regulation of malate metabolism in grape berry and other developing fruits. Phytochemistry 2009, 70, 1329–1344. [Google Scholar] [CrossRef]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Gerós, H. Biochemical Changes throughout Grape Berry Development and Fruit and Wine Quality; Global Science Books: London, UK, 2007. [Google Scholar]
- Rolle, L.; Río Segade, S.; Torchio, F.; Giacosa, S.; Cagnasso, E.; Marengo, F.; Gerbi, V. Influence of grape density and harvest date on changes in phenolic composition, phenol extractability indices, and instrumental texture properties during ripening. J. Agric. Food Chem. 2011, 59, 8796–8805. [Google Scholar] [CrossRef]
- Jediyi, H.; Naamani, K.; Elkoch, A.A.; Dihazi, A.; El Fels, A.E.A.; Arkize, W. First study on technological maturity and phenols composition during the ripeness of five V. vinifera L grape varieties in Morocco. Sci. Hortic. 2019, 246, 390–397. [Google Scholar] [CrossRef]
- Zhang, X.; Kontoudakis, N.; Suklje, K.; Antalick, G.; Blackman, J.W.; Rutledge, D.N.; Clark, A.C. Changes in red wine composition during bottle aging: Impacts of grape variety, vineyard location, maturity, and oxygen availability during aging. J. Agric. Food Chem. 2020, 68, 13331–13343. [Google Scholar] [CrossRef]
- Roberto, S.R.; de Assis, A.M.; Yamamoto, L.Y.; Miotto, L.C.V.; Sato, A.J.; Koyama, R.; Genta, W. Application timing and concentration of abscisic acid improve color of ‘Benitaka’ table grape. Sci. Hortic. 2012, 142, 44–48. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Gutiérrez-Gamboa, G.; Fernández-Novales, J.; Pérez-Álvarez, E.P.; Diago, M.P. Towards the definition of optimal grape harvest time in Grenache grapevines: Nitrogenous maturity. Sci. Hortic. 2018, 239, 9–16. [Google Scholar] [CrossRef]
- Magwaza, L.S.; Mditshwa, A.; Tesfay, S.Z.; Opara, U.L. An overview of preharvest factors affecting vitamin C content of citrus fruit. Sci. Hortic. 2017, 216, 12–21. [Google Scholar] [CrossRef]
- Verdugo-Vásquez, N.; Acevedo-Opazo, C.; Valdés-Gómez, H.; Pañitrur-De la Fuente, C.; Ingram, B.; Garcia de Cortazar-Atauri, I.; Tisseyre, B. Identification of main factors affecting the within-field spatial variability of grapevine phenology and total soluble solids accumulation: Towards the vineyard zoning using auxiliary information. Precis. Agric. 2022, 23, 253–277. [Google Scholar] [CrossRef]
- Kaya, O.; Delavar, H.; Ates, F.; Yilmaz, T.; Sahin, M.; Keskin, N. Fine-Tuning Grape Phytochemistry: Examining the Distinct Influence of Oak Ash and Potassium Carbonate Pre-Treatments on Essential Components. Horticulturae 2024, 10, 95. [Google Scholar] [CrossRef]
- Keskin, N.; Bilir Ekbic, H.; Kaya, O.; Keskin, S. Antioxidant activity and biochemical compounds of Vitis vinifera L.(cv.‘Katıkara’) and Vitis labrusca L.(cv.‘Isabella’) grown in Black Sea Coast of Turkey. Erwerbs-Obstbau 2021, 63 (Suppl. S1), 115–122. [Google Scholar] [CrossRef]
- Tebib, K.; Rouanet, J.M.; Besançon, P. Antioxidant effects of dietary polymeric grape seed tannins in tissues of rats fed a high cholesterol-vitamin E-deficient diet. Food Chem. 1997, 59, 135–141. [Google Scholar] [CrossRef]
- Teissedre, P.L. Wine and health. Biochem. Grape Berry 2012, 269, 588. [Google Scholar]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation, and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Liu, Y.; Yue, T.X.; Zhang, Z.W. Comparison between aroma compounds in wines from four Vitis vinifera grape varieties grown in different shoot positions. Food Sci. Technol. 2015, 35, 237–246. [Google Scholar] [CrossRef]
- Yue, X.; Zhao, Y.; Ma, X.; Jiao, X.; Fang, Y.; Zhang, Z.; Ju, Y. Effects of leaf removal on the accumulation of anthocyanins and the expression of anthocyanin biosynthetic genes in Cabernet Sauvignon (Vitis vinifera L.) grapes. J. Sci. Food Agric. 2021, 101, 3214–3224. [Google Scholar] [CrossRef] [PubMed]
- Ates, F.; Delavar, H.; Dardeniz, A.; Yilmaz, T.; Turan, M.; Kaya, O. Dynamics of berry characteristics, biochemical composition, and physiological responses across ripening stages: Investigating the impact of pollinizer varieties on physiological femaleness in Bozcaada Çavuşu (Vitis vinifera L. cv). J. Plant Growth Regul. 2024, 1–20. [Google Scholar] [CrossRef]
- Lisov, N.; Čakar, U.; Milenković, D.; Čebela, M.; Vuković, G.; Despotović, S.; Petrović, A. The Influence of Cabernet Sauvignon Ripeness, Healthy State and Maceration Time on Wine and Fermented Pomace Phenolic Profile. Fermentation 2023, 9, 695. [Google Scholar] [CrossRef]



{kind=link}
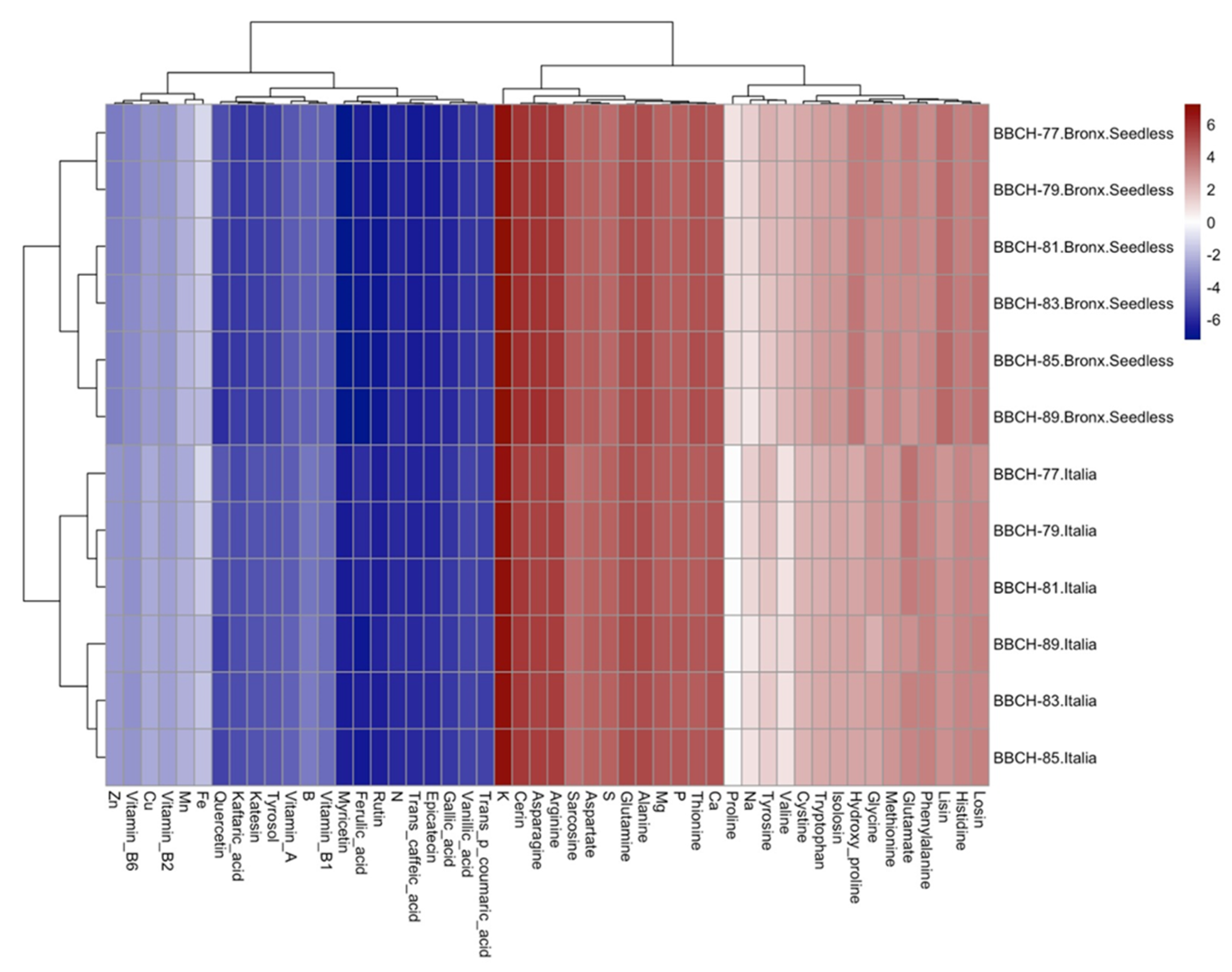{kind=link}
| Berry Development Stages | Berry Weight (g/Berry) | Berry Width (mm) | Berry Length (mm) | Total Soluble Solid (oBrix) | Titratable Acidity (g/L as Tartaric Acid)-TA | Maturity Index (MI-oBrix) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Italia | Bronx Seedless | Italia | Bronx Seedless | Italia | Bronx Seedless | Italia | Bronx Seedless | Italia | Bronx Seedless | Italia | Bronx Seedless | |
| BBCH-77 | 0.68 | 0.41 | 12.35 | 8.89 | 16.15 | 11.65 | 2.87 | 2.70 | 34.98 | 29.78 | 0.88 | 0.95 |
| BBCH-79 | 2.67 | 1.34 | 16.65 | 12.54 | 20.34 | 14.78 | 4.54 | 4.67 | 29.76 | 24.56 | 1.48 | 1.65 |
| BBCH-81 | 4.43 | 1.72 | 18.45 | 13.23 | 22.43 | 16.98 | 9.83 | 10.45 | 18.56 | 15.43 | 5.34 | 6.88 |
| BBCH-83 | 6.41 | 2.58 | 19.34 | 14.15 | 23.45 | 17.76 | 14.34 | 14.67 | 10.45 | 8.34 | 13.34 | 16.67 |
| BBCH-85 | 7.86 | 3.21 | 20.87 | 15.85 | 25.42 | 18.45 | 15.21 | 15,54 | 9.58 | 7.45 | 16.23 | 20.76 |
| BBCH-89 | 8.13 | 3.59 | 21.76 | 16.49 | 26.37 | 19.67 | 16.85 | 17.56 | 6.56 | 5.67 | 25.12 | 32.87 |
| Cultivar X (C) | Aspartate | Glutamate | Asparagine | Cerin | Glutamine | Histidine | Glycine | Thionine | Arginine | Alanine | Tyrosine |
| Italia | 5200 ± 44 b | 2866 ± 51 a | 9803 ± 13 b | 11,265 ± 29 b | 8004 ± 14 | 2590 ± 4.4 b | 1566 ± 3 b | 5979 ± 11 b | 10,508 ± 27 b | 7974 ± 13 | 753 ± 13 b |
| Bronx Seedless | 5537 ± 34 a | 2500 ± 58 b | 12,995 ± 15 a | 14,045 ± 23 a | 7804 ± 15 | 3044 ± 4.8 a | 2325 ± 4 a | 7374 ± 19 a | 11,691 ± 20 a | 8357 ± 12 | 860 ± 12 a |
| Phenological stage Y (PS) | |||||||||||
| BBCH-77 | 3949 ± 74 f | 2820 ± 12 | 8343 ± 25 e | 9547 ± 504 e | 5859 ± 23 e | 2125 ± 81 e | 2045 ± 43 | 5037 ± 33 d | 8374 ± 48 d | 5857 ± 22 e | 848 ± 11 |
| BBCH-79 | 4431 ± 71 e | 2764 ± 10 | 9378 ± 27 d | 10,617 ± 517 de | 6556 ± 21 de | 2363 ± 23 d | 2004 ± 34 | 5601 ± 31 cd | 9312 ± 49 cd | 6630 ± 24 d | 831 ± 23 |
| BBCH-81 | 4972 ± 72 d | 2709 ± 13 | 10,540 ± 23 c | 11,806 ± 407 cd | 7336 ± 25 cd | 2628 ± 41 cd | 1964 ± 53 | 6229 ± 35 bc | 10,355 ± 46 bc | 7505 ± 26 c | 814 ± 20 |
| BBCH-83 | 5578 ± 73 c | 2654 ± 15 | 11,847 ± 24 b | 13,128 ± 307 bc | 8209 ± 22 bc | 2922 ± 83 bc | 1925 ± 55 | 6926 ± 11 ab | 11,514 ± 68 ab | 8496 ± 29 bc | 798 ± 18 |
| BBCH-85 | 6259 ± 71 b | 2601 ± 16 | 13,316 ± 26 ab | 14,598 ± 506 ab | 9186 ± 21 ab | 3249 ± 45 ab | 1886 ± 34 | 7702 ± 33 ab | 12,804 ± 45 a | 9618 ± 22 ab | 782 ± 19 |
| BBCH-89 | 7022 ± 73 a | 2549 ± 14 | 14,968 ± 25 a | 16,233 ± 517 ab | 10,279 ± 23 a | 3613 ± 74 a | 1849 ± 32 | 8565 ± 35 a | 14,238 ± 46 a | 10,887 ± 26 a | 766 ± 12 |
| Significance | |||||||||||
| C | *** | *** | *** | *** | 0.3490 | *** | *** | *** | ** | 0.0513 | *** |
| PS | *** | 0.4420 | *** | *** | *** | *** | 0.1991 | *** | *** | *** | 0.1045 |
| C × PS | 0.9521 | 0.9999 | 0.8999 | 0.9118 | 1.0000 | 0.9047 | 0.9975 | 0.9755 | 0.9970 | 0.9991 | 1.0000 |
| Cultivar X (C) | Cystine | Valine | Methionine | Tryptophan | Phenylalanine | Isolosin | Losin | Lisin | Hydroxyproline | Sarcosine | Proline |
| Italia | 1030 ± 11 b | 435 ± 22 b | 1843 ± 43 b | 1229 ± 18 b | 2753 ± 40 a | 1420 ± 31 b | 2630 ± 45 b | 2039 ± 34 b | 1391 ± 36 b | 4174 ± 103 b | 257 ± 7 b |
| Bronx Seedless | 1289 ± 16 a | 969 ± 24 a | 2450 ± 40 a | 1769 ± 11 a | 2463 ± 50 b | 1864 ± 32 a | 3772 ± 65 a | 4509 ± 55 a | 3427 ± 56 a | 5730 ± 104 a | 460 ± 7 a |
| Phenological stage Y (PS) | |||||||||||
| BBCH-77 | 875 ± 21 e | 530 ± 35 d | 1620 ± 71 e | 1075 ± 32 e | 1967 ± 83 e | 1239 ± 16 e | 2296 ± 111 e | 2470 ± 95 e | 1728 ± 91 e | 3736 ± 170 e | 271 ± 14 e |
| BBCH-79 | 973 ± 18 d | 589 ± 32 cd | 1801 ± 60 de | 1217 ± 23 d | 2188 ± 87de | 1377 ± 45 de | 2599 ± 114 d | 2747 ± 92 d | 1956 ± 87 de | 4154 ± 183 de | 301 ± 32 de |
| BBCH-81 | 1082 ± 23 c | 655 ± 31 bc | 2003 ± 45 cd | 1378 ± 24 c | 2433 ± 37 cd | 1532 ± 56 cd | 2942 ± 124 cd | 3054 ± 90 cd | 2215 ± 96 cd | 4620 ± 180 cd | 335 ± 13 cd |
| BBCH-83 | 1203 ± 12 bc | 729 ± 33 abc | 2227 ± 65 bc | 1559 ± 31 b | 2705 ± 83 bc | 1703 ± 53 bc | 3331 ± 143 bc | 3397 ± 65 bc | 2507 ± 90 bc | 5137 ± 185 bc | 372 ± 11 bc |
| BBCH-85 | 1338 ± 23 ab | 810 ± 18 ab | 2476 ± 73 ab | 1765 ± 33 a | 3008 ± 59 ab | 1894 ± 54 ab | 3770 ± 134 ab | 3777 ± 92 ab | 2838 ± 77 ab | 5712 ± 179 ab | 414 ± 13 ab |
| BBCH-89 | 1487 ± 26 a | 901 ± 28 a | 2754 ± 71 a | 1998 ± 35 a | 3345 ± 46 a | 2106 ± 51 a | 4268 ± 115 a | 4200 ± 93 a | 3212 ± 59 a | 6352 ± 174 a | 460 ± 22 a |
| Significance | |||||||||||
| C | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| PS | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| C × PS | 0.5546 | 0.9887 | 0.6087 | 0.8765 | 0.9897 | 0.6915 | 0.8976 | 0.9596 | 0.8799 | 0.6099 | 0.9876 |
| Cultivar X (C) | Gallic Acid | Vanillic Acid | Trans-Caffeic Acid | Trans P-Coumaric Acid | Ferulic Acid | Kaftaric Acid | Catechin | Epicatechin | Quercetin | Rutin | Myricetin | Tyrosol |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Italia | 5.1 ± 0.1 a | 6.3 ± 0.0 a | 3.6 ± 0.1 a | 5.9 ± 0.0 a | 3.1 ± 0.03 a | 7.54 ± 0.0 a | 8.5 ± 0.1 a | 3.8 ± 0.1 a | 7.1 ± 0.0 a | 3.2 ± 0.10 a | 2.9 ± 0.0 a | 9.1 ± 0.0 a |
| Bronx Seedless | 3.7 ± 0.0 b | 4.84 ± 0.2 b | 2.9 ± 0.0 b | 4.9 ± 0.1 b | 2.4 ± 0.02 b | 5.35 ± 0.1 b | 5.7 ± 0.2 b | 3.2 ± 0.0 b | 5.7 ± 0.0 b | 2.4 ± 0.0 b | 1.7 ± 0.0 b | 6.3 ± 0.2 b |
| Phenological stage Y (PS) | ||||||||||||
| BBCH-77 | 3.2 ± 0.1 f | 4.2 ± 0.0 f | 2.4 ± 0.0 e | 4.1 ± 0.0 f | 2.8 ± 0.1 a | 2.6 ± 0.1 f | 5.3 ± 0.1 e | 2.5 ± 0.0 f | 6.7 ± 0.2 a | 2.1 ± 0.0 e | 1.8 ± 0.1 d | 5.8 ± 0.2 e |
| BBCH-79 | 3.6 ± 0.2 e | 4.7 ± 0.2 e | 2.7 ± 0.0 d | 4.52 ± 0.2 e | 2.82 ± 0.0 ab | 3.9 ± 0.2 e | 5.9 ± 0.1 d | 2.8 ± 0.0 e | 6.6 ± 0.1 ab | 2.3 ± 0.2 de | 1.9 ± 0.3 cd | 6.4 ± 0.3 d |
| BBCH-81 | 4.1 ± 0.0 d | 5.2 ± 0.0 d | 3.0 ± 0.1 c | 5.0 ± 0.3 d | 2.8 ± 0.0 ab | 4.8 ± 0.1 d | 6.6 ± 0.2 cd | 3.2 ± 0.1 d | 6.4 ± 0.1 abc | 2.6 ± 0.0 cd | 2.2 ± 0.1 bcd | 7.2 ± 0.1 c |
| BBCH-83 | 4.6 ± 0.0 c | 5.8 ± 0.1 c | 3.4 ± 0.0 bc | 5.6 ± 0.0 c | 2.7 ± 0.3 ab | 5.1 ± 0.2 c | 6.6 ± 0.2 bc | 3.6 ± 0.1 c | 6.3 ± 0.2 abc | 2.9 ± 0.1 bc | 2.4 ± 0.2 abc | 7.9 ± 0.1 b |
| BBCH-85 | 5.2 ± 0.3 b | 6.4 ± 0.3 b | 3.8 ± 0.3 ab | 6.2 ± 0.0 b | 2.7 ± 0.0 ab | 6.6 ± 0.1 b | 8.1 ± 0.1 ab | 4.1 ± 0.0 b | 6.2 ± 0.1 bc | 3.2 ± 0.0 ab | 2.7 ± 0.1 ab | 8.9 ± 0.3 a |
| BBCH-89 | 5.8 ± 0.3 a | 7.1 ± 0.0 a | 4.2 ± 0.0 a | 6.9 ± 0.0 a | 2.6 ± 0.1 b | 7.4 ± 0.1 a | 9.1 ± 0.1 a | 4.6 ± 0.0 a | 6.0 ± 0.1 c | 3.6 ± 0.1 a | 3.0 ± 0.3 a | 9.9 ± 0.1 a |
| Significance | ||||||||||||
| C | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| PS | *** | *** | *** | *** | 0.0107 * | *** | *** | *** | 0.0050 ** | *** | *** | *** |
| C × PS | 0.9401 | 0.8947 | 0.6219 | 0.8731 | 0.9957 | 0.9122 | 0.0885 | 0.5099 | 0.9978 | 0.8495 | 0.9525 | 0.9019 |
| Cultivar X (C) | N (%) | Ca | K | Mg | Na | P | S | Mn | Cu | Fe | Zn | B |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Italia | 3.9 ± 0.2 a | 6833 ± 4 | 28,184 ± 211 b | 6054 ± 46 a | 480 ± 14 | 5861 ± 33.1 | 5280 ± 68 a | 50.1 ± 1.1 b | 46.8 ± 3.3 a | 81.9 ± 0.3 b | 33.5 ± 3.5 a | 18.0 ± 2.0 a |
| Bronx Seedless | 3.6 ± 0.0 b | 6812 ± 5 | 31,089 ± 215 a | 5900 ± 35 b | 506 ± 11 | 5758 ± 38.6 | 4694 ± 66 b | 56.0 ± 1.6 a | 34.2 ± 3.8 b | 87.1 ± 0.2 a | 20.8 ± 3.8 b | 11.7 ± 1.0 b |
| Phenological stage Y (PS) | ||||||||||||
| BBCH-77 | 2.8 ± 0.0 f | 5147 ± 10 f | 22,359 ± 37 f | 4397 ± 83 f | 518 ± 21 | 4252 ± 66 f | 3762 ± 118 e | 39.3 ± 2.3 d | 30.6 ± 6.1 | 88.8 ± 0.4 a | 20.5 ± 6.2 | 11.2 ± 3.4 |
| BBCH-79 | 3.1 ± 0.1 e | 5724 ± 13 e | 24,863 ± 373 e | 4933 ± 80 e | 508 ± 22 | 4779 ± 64 e | 4184 ± 111 d | 44.0 ± 2.3 cd | 34.0 ± 6.6 | 87.1 ± 0.5 ab | 22.7 ± 5.6 | 12.5 ± 2.4 |
| BBCH-81 | 3.5 ± 0.0 d | 6365 ± 20 d | 27,648 ± 323 d | 5535 ± 81 d | 498 ± 23 | 5372 ± 65 d | 4652 ± 132 c | 49.2 ± 2.5 bc | 37.8 ± 6.0 | 85.3 ± 0.7 bc | 25.3 ± 6.4 | 13.9 ± 3.3 |
| BBCH-83 | 3.9 ± 0.2 c | 7077 ± 12 c | 30,744 ± 333 c | 6210 ± 86 c | 488 ± 24 | 6038 ± 62 c | 5174 ± 121 b | 55.1 ± 2.4 abc | 42.0 ± 6.3 | 83.6 ± 0.6 cd | 28.1 ± 5.6 | 15.4 ± 2.5 |
| BBCH-85 | 4.3 ± 0.0 b | 7870 ± 21 b | 34,188 ± 345 b | 6968 ± 70 b | 478 ± 19 | 6787 ± 55 b | 5753 ± 112 ab | 61.6 ± 2.5 ab | 46.7 ± 6.4 | 81.9 ± 0.3 d e | 31.3 ± 5.6 | 17.1 ± 3.5 |
| BBCH-89 | 4.8 ± 0.3 a | 8752 ± 13 a | 38,017 ± 356 a | 7818 ± 82 a | 469 ± 18 | 7628 ± 49 a | 6397 ± 122 a | 69.0 ± 2.8 a | 52.0 ± 6.2 | 80.3 ± 0.5 e | 34.8 ± 6.0 | 19.1 ± 3.1 |
| Significance | ||||||||||||
| C | *** | 0.7951 | *** | *** | 0.2078 | 0.0708 | *** | 0.0203 * | 0.0283 * | *** | 0.0285 * | 0.0393 * |
| PS | *** | *** | *** | *** | 0.7081 | *** | *** | *** | 0.2414 | *** | 0.6661 | 0.6296 |
| C × PS | 0.7235 | 1.000 | 0.7046 | 0.9989 | 1.000 | 0.9996 | 0.9413 | 0.9986 | 0.9992 | 0.9999 | 0.9992 | 0.9995 |
| Cultivar X (C) | Vitamin A | Vitamin B1 | Vitamin B2 | Vitamin B6 | Vitamin C |
|---|---|---|---|---|---|
| Italia | 11.0 ± 0.2 a | 14.1 ± 0.2 a | 36.9 ± 0.6 a | 29.5 ± 0.3 a | 3.9± 0.0 a |
| Bronx Seedless | 10.4 ± 0.0 b | 11.5 ± 0.1 b | 31.6 ± 0.5 b | 24.7 ± 0.2 b | 3.5± 0.1 b |
| Phenological stage Y (PS) | |||||
| BBCH-77 | 8.1 ± 0.2 f | 10.5 ± 0.4 d | 25.6 ± 1.1 e | 20.6 ± 0.3 e | 2.7 ± 0.0 f |
| BBCH-79 | 8.9 ± 0.1 e | 11.3 ± 0.3 cd | 28.6 ± 1.3 de | 22.8 ± 0.5 d | 3.1 ± 0.1 e |
| BBCH-81 | 9.9 ± 0.2 d | 12.2 ± 0.4 bc | 31.9 ± 1.0 cd | 25.3 ± 0.4 c | 3.5 ± 0.0 d |
| BBCH-83 | 11.1 ± 0.1 c | 13.2 ± 0.2 ab | 35.6 ± 1.4 bc | 28.1 ± 0.5 b | 3.9 ± 0.1 c |
| BBCH-85 | 12.3 ± 0.2 b | 14.3 ± 0.1 a | 39.7 ± 1.1 ab | 31.2 ± 0.2 ab | 4.4 ± 0.0 b |
| BBCH-89 | 13.7 ± 0.1 a | 15.4 ± 0.4 a | 44.3 ± 1.0 a | 34.6 ± 0.3 a | 4.9 ± 0.0 a |
| Significance | |||||
| C | *** | *** | *** | *** | *** |
| PS | *** | *** | *** | *** | *** |
| C × PS | 0.9267 | 0.9564 | 0.9372 | 0.8664 | 0.9873 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatterman-Valenti, H.; Kaya, O.; Yilmaz, T.; Ates, F.; Turan, M. Phenolic, Amino Acid, Mineral, and Vitamin Contents during Berry Development in ‘Italia’ and ‘Bronx Seedless’ Grape Cultivars. Horticulturae 2024, 10, 429. https://doi.org/10.3390/horticulturae10050429
Hatterman-Valenti H, Kaya O, Yilmaz T, Ates F, Turan M. Phenolic, Amino Acid, Mineral, and Vitamin Contents during Berry Development in ‘Italia’ and ‘Bronx Seedless’ Grape Cultivars. Horticulturae. 2024; 10(5):429. https://doi.org/10.3390/horticulturae10050429
Chicago/Turabian StyleHatterman-Valenti, Harlene, Ozkan Kaya, Turhan Yilmaz, Fadime Ates, and Metin Turan. 2024. "Phenolic, Amino Acid, Mineral, and Vitamin Contents during Berry Development in ‘Italia’ and ‘Bronx Seedless’ Grape Cultivars" Horticulturae 10, no. 5: 429. https://doi.org/10.3390/horticulturae10050429
APA StyleHatterman-Valenti, H., Kaya, O., Yilmaz, T., Ates, F., & Turan, M. (2024). Phenolic, Amino Acid, Mineral, and Vitamin Contents during Berry Development in ‘Italia’ and ‘Bronx Seedless’ Grape Cultivars. Horticulturae, 10(5), 429. https://doi.org/10.3390/horticulturae10050429