
The Discovery of the Potential Attractive Compounds of Bactrocera dorsalis (Hendel)

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Collection, Identification and Enrichment of Olfactory Sensory Proteins in B. dorsalis
2.2.1. RNA Extraction, cDNA Library Preparation and Sequencing
2.2.2. Gene Annotation and Sequence Analysis
2.2.3. Phylogenetic Analyses
2.2.4. GO Functional Enrichment and KEGG Pathway Enrichment Analyses
2.3. Collection of Potential Attractive Compounds
2.4. Molecular Docking
2.5. Screening and Prediction of Core Functional Compounds
2.6. Behavioral Assays
3. Results
3.1. Collection, Identification and Enrichment of Olfactory Sensory Proteins in B. dorsalis
3.1.1. Phylogenetic Analyses
3.1.2. GO Functional Enrichment and KEGG Pathway Enrichment Analyses
3.2. Collection of Potential Attractive Compounds
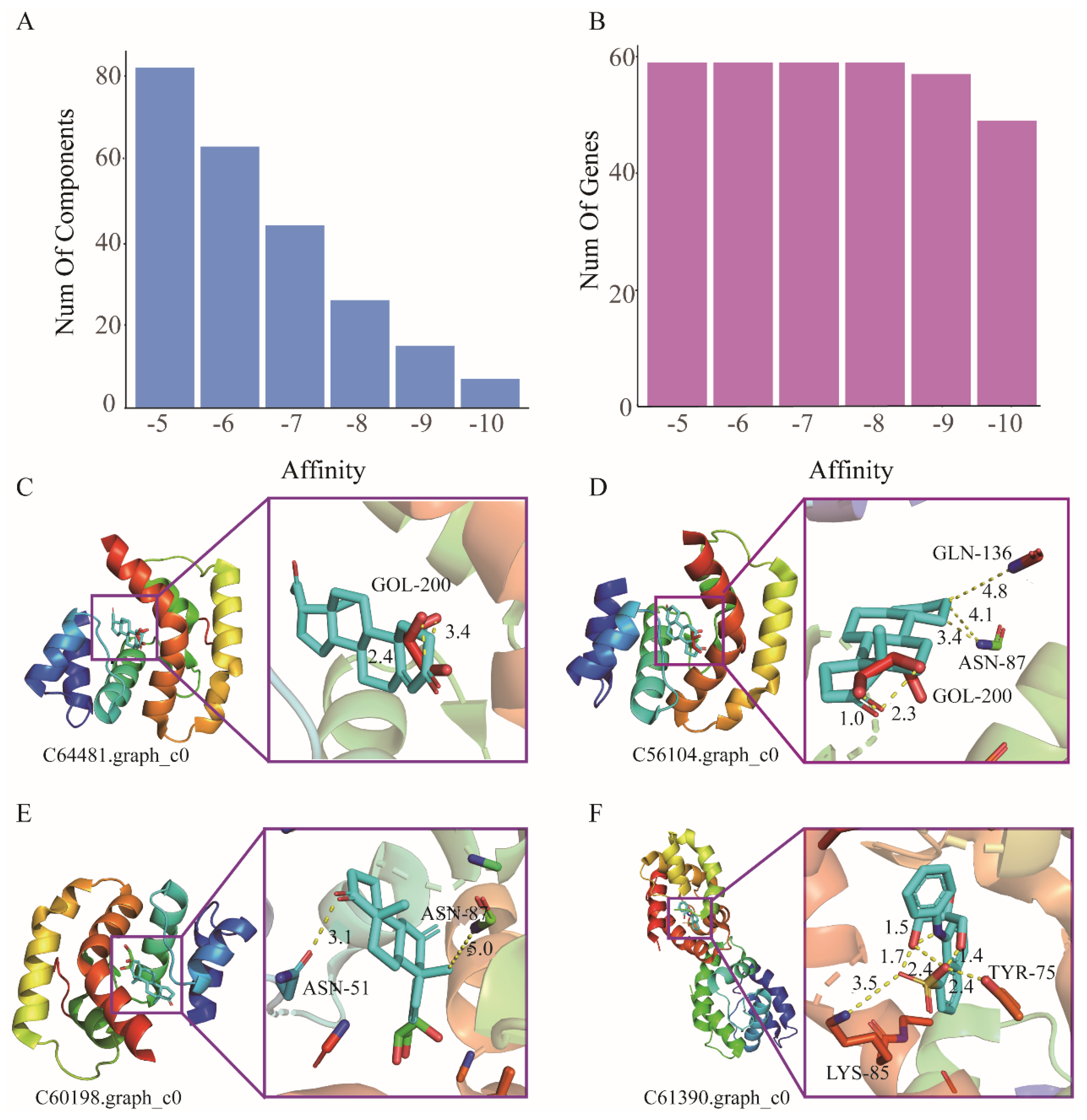
3.3. Molecular Docking
3.4. Screening and Prediction of Core Functional Compounds
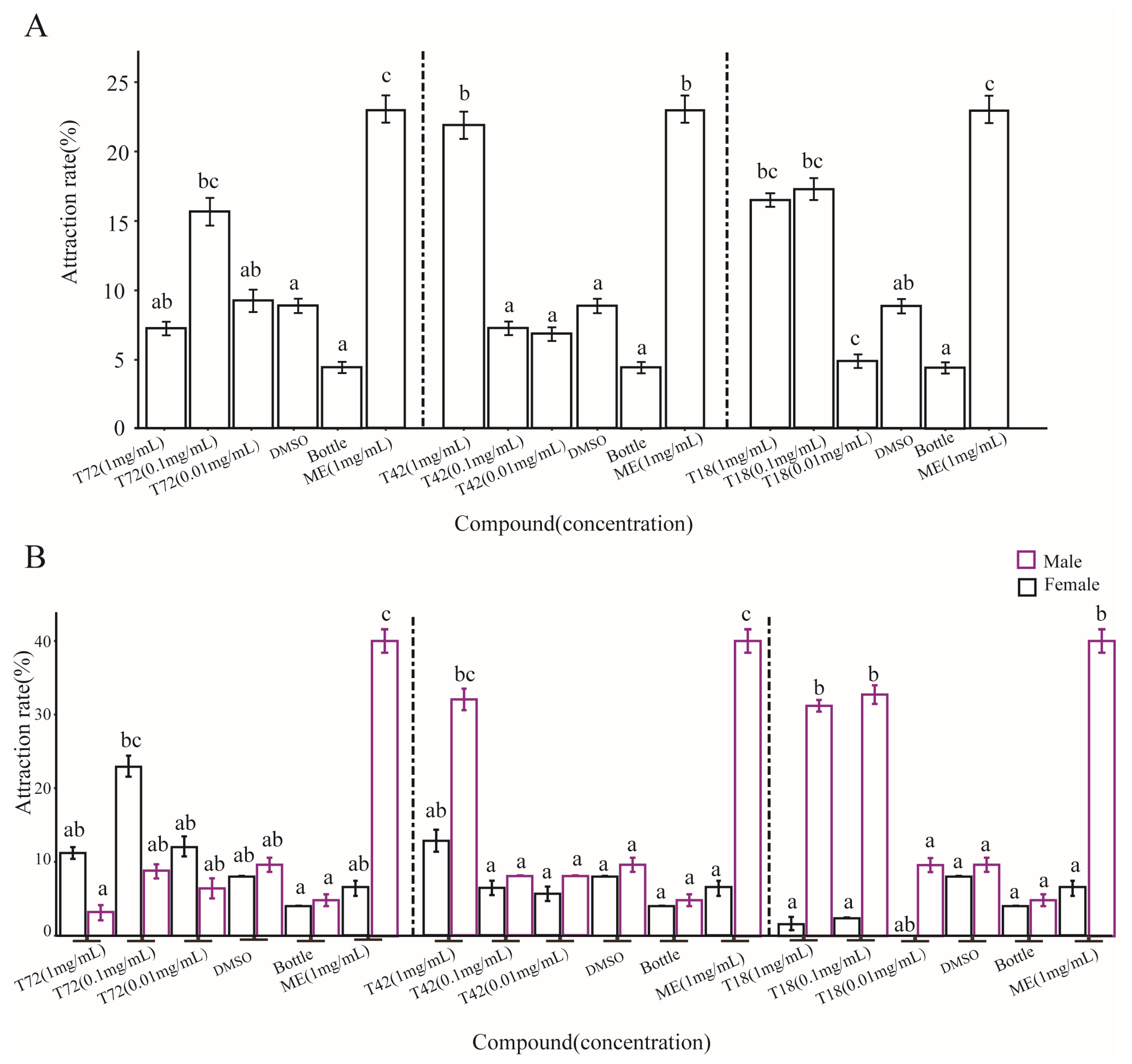
3.5. Behavioral Assays
4. Discussions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| Apisu | Acyrthosiphon pisum |
| B. dorsalis | Bactrocera dorsalis |
| Bcuc | Bactrocera cucurbitae |
| Bcuc | Zeugodacus cucurbitae |
| Bdor | Bactrocera dorsalis |
| Bmori | Bombyx mori |
| Bole | Bactrocera oleae |
| Ccap | Ceratitis capitata |
| CPGP network | Compound–pathway–GO term–olfactory sensory protein network |
| CSP | Chemosensory protein |
| Dana | Drosophila ananassae |
| Dere | Drosophila erecta |
| Dgri | Drosophila grimshawi |
| Dmel | Drosophila melanogaster |
| Dmoj | Drosophila mojavensis |
| Dper | Drosophila persimilis |
| Dpse | Drosophila pseudoananassae |
| Dsec | Drosophila sechellia |
| Dsim | Drosophila simulans |
| Dvir | Drosophila virilis |
| Dwil | Drosophila willistoni |
| Dyak | Drosophila Yakuba |
| GO | Gene ontology |
| GR | Gustatory receptor |
| IR | Ionotropic receptor |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| ME | Methyl eugenol |
| MP | Maximum parsimony |
| OBP | Odorant-binding protein |
| ORF | Open reading frame |
| OR | Olfactory receptor |
| PBP | Pheromone-binding protein |
| SPR | Subtree pruning and regrafting |
| SNMP | Sensory neuron membrane protein |
| TCM | Traditional Chinese Medicine |
References
- Schutze, M.K.; Bourtzis, K.; Cameron, S.L.; Clarke, A.R.; De Meyer, M.; Hee, A.K.W.; Hendrichs, J.; Krosch, M.N.; Mwatawala, M. Integrative Taxonomy versus Taxonomic Authority without Peer Review: The Case of the Oriental Fruit Fly, Bactrocera dorsalis (Tephritidae). Syst. Entomol. 2017, 42, 609–620. [Google Scholar] [CrossRef]
- Vargas, R.I.; Pinero, J.C.; Leblanc, L. An Overview of Pest Species of Bactrocera Fruit Flies (Diptera: Tephritidae) and the Integration of Biopesticides with Other Biological Approaches for Their Management with a Focus on the Pacific Region. Insects 2015, 6, 297–318. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Liang, L.; Wang, J.; Zhu, S. Chromosome-Level Genome Assembly of Bactrocera dorsalis Reveals Its Adaptation and Invasion Mechanisms. Commun. Biol. 2022, 5, 25. [Google Scholar]
- Yu, C.; Zhao, R.; Zhou, W.; Pan, Y.; Tian, H.; Yin, Z.; Chen, W. Fruit Fly in a Challenging Environment: Impact of Short-Term Temperature Stress on the Survival, Development, Reproduction, and Trehalose Metabolism of Bactrocera dorsalis (Diptera: Tephritidae). Insects 2022, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; He, Y.; Ren, Y.; Wang, G.; Chu, D. Seasonal and Year-Round Distributions of Bactrocera dorsalis (Hendel) and Its Risk to Temperate Fruits under Climate Change. Insects 2022, 13, 550. [Google Scholar] [CrossRef]
- Dias, N.P.; Zotti, M.J.; Montoya, P.; Carvalho, I.R.; Nava, D.E. Fruit Fly Management Research: A Systematic Review of Monitoring and Control Tactics in the World. Crop Prot. 2018, 112, 187–200. [Google Scholar] [CrossRef]
- Tan, K.H.; Nishida, R. Methyl Eugenol: Its Occurrence, Distribution, and Role in Nature, Especially in Relation to Insect Behavior and Pollination. J. Insect Sci. 2012, 12, 56. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, C.; Qin, Y.; Yin, X.; Dong, X.; Desneux, N.; Zhou, H. Monitoring the Methyl Eugenol Response and Non-Responsiveness Mechanisms in Oriental Fruit Fly Bactrocera dorsalis in China. Insects 2022, 13, 1004. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, X.; Zhang, B.; Cheng, Y. Building Methodology for Discovering and Developing Chinese Medicine Based on Network Biology. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Medica 2011, 36, 228–231. [Google Scholar]
- Wu, L.; Wang, Y.; Fan, X. Tools for Network Pharmacology Study: Network Visualization and Network Analysis. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Medica 2011, 36, 2923–2925. [Google Scholar]
- Wang, Z.-Y.; Wang, X.; Zhang, D.-Y.; Hu, Y.-J.; Li, S. Traditional Chinese Medicine Network Pharmacology: Development in New Era under Guidance of Network Pharmacology Evaluation Method Guidance. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Medica 2022, 47, 7–17. [Google Scholar] [CrossRef]
- Sun, J.S.; Xiao, S.; Carlson, J.R. The Diverse Small Proteins Called Odorant-Binding Proteins. R. Soc. Open Biol. 2018, 8, 180208. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Li, G.C.; Zhu, J.Y.; Liu, N.Y. Genome-Based Analysis Reveals a Novel SNMP Group of the Coleoptera and Chemosensory Receptors in Rhaphuma Horsfieldi. Genomics 2020, 112, 2713–2728. [Google Scholar] [CrossRef] [PubMed]
- Robertson, H.M. Molecular Evolution of the Major Arthropod Chemoreceptor Gene Families. Annu. Rev. Entomol. 2019, 64, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Karpe, S.D.; Tiwari, V.; Ramanathan, S. InsectOR-Webserver for Sensitive Identification of Insect Olfactory Receptor Genes from Non-Model Genomes. PLoS ONE 2021, 16, e0245324. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, H.; Wang, Z.; Bin, S.; He, H.; Lin, J. Discovery of Chemosensory Genes in the Oriental Fruit Fly, Bactrocera dorsalis. PLoS ONE 2015, 10, e0129794. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Zhao, M.; Zhong, L.; Li, Y.; Li, D.; Deng, Z.; Ma, X. Characterization of the Binding Ability of the Odorant Binding Protein BminOBP9 of Bactrocera Minax to Citrus Volatiles. Pest Manag. Sci. 2021, 77, 1214–1225. [Google Scholar] [CrossRef]
- Yi, X.; Zhao, H.; Wang, P.; Hu, M.; Zhong, G. Bdor\Orco is Important for Oviposition-Deterring Behavior Induced by Both the Volatile and Non-Volatile Repellents in Bactrocera dorsalis (Diptera: Tephritidae). J. Insect Physiol. 2014, 65, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Hong, J.; Wang, C.; Yang, Y.; Zhang, Q.; Ji, Q.; Chen, J. Radiation of Bactrocera dorsalis (Diptera: Tephritidae) Eggs to Improve the Mass Rearing of Diachasmimorpha longicaudata (Hymenoptera: Braconidae). J. Econ. Entomol. 2018, 111, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-Throughput Functional Annotation and Data Mining with the Blast2go Suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2go: A Universal Tool for Annotation, Visualization and Analysis in Functional Genomics Research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.L.; He, P.; Zhang, L.; Fang, S.Q.; Dong, S.L.; Zhang, Y.J.; Li, F. Large-Scale Identification of Odorant-Binding Proteins and Chemosensory Proteins from Expressed Sequence Tags in Insects. BMC Genom. 2009, 10, 632. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating Signal Peptides from Transmembrane Regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Parallelization of the MAFFT Multiple Sequence Alignment Program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef] [PubMed]
- Vieira, F.G.; Rozas, J. Comparative Genomics of the Odorant-Binding and Chemosensory Protein Gene Families across the Arthropoda: Origin and Evolutionary History of the Chemosensory System. Genome Biol. Evol. 2011, 3, 476–490. [Google Scholar] [CrossRef]
- Siciliano, P.; Scolari, F.; Gomulski, L.M.; Falchetto, M.; Manni, M.; Gabrieli, P.; Field, L.M.; Zhou, J.J.; Gasperi, G.; Malacrida, A.R. Sniffing Out Chemosensory Genes from the Mediterranean Fruit Fly, Ceratitis capitata. PLoS ONE 2014, 9, e85523. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. Clusterprofiler: An R Package for Comparing Biological Themes among Gene Clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y. KEGG Mapper for Inferring Cellular Functions from Protein Sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef]
- Li, J.; Fu, A.; Zhang, L. An Overview of Scoring Functions Used for Protein-Ligand Interactions in Molecular Docking. Interdiscip. Sci. 2019, 11, 320–328. [Google Scholar] [CrossRef]
- Elhenawy, A.A.; Al-Harbi, L.M.; El-Gazzar, M.A.; Khowdiary, M.M.; Ouidate, A.; Alosaimi, A.M.; Elhamid Salim, A. Naproxenylamino Acid Derivatives: Design, Synthesis, Docking, QSAR and Anti-Inflammatory and Analgesic Activity. Biomed. Pharmacother. 2019, 116, 109024. [Google Scholar] [CrossRef]
- Wu, B.; Lan, X.; Chen, X.; Wu, Q.; Yang, Y.; Wang, Y. Researching the Molecular Mechanisms of Taohong Siwu Decoction in the Treatment of Varicocele-Associated Male Infertility Using Network Pharmacology and Molecular Docking: A Review. Medicine 2023, 102, e34476. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.X.; Chen, Y.; Lei, Q.; Peng, Y.Y.; Jiang, H.B. Sublethal Dose of β-Cypermethrin Impairs the Olfaction of Bactrocera dorsalis by Suppressing the Expression of Chemosensory Genes. Insects 2022, 13, 721. [Google Scholar] [CrossRef] [PubMed]
- George, L.F.; Pradhan, S.J.; Mitchell, D.; Josey, M.; Casey, J.; Belus, M.T.; Fedder, K.N.; Dahal, G.R.; Bates, E.A. Ion Channel Contributions to Wing Development in Drosophila melanogaster. G3 Genes Genomes Genet. 2019, 9, 999–1008. [Google Scholar] [CrossRef]
- Prelic, S.; Getahun, M.N.; Kaltofen, S.; Hansson, B.S.; Wicher, D. Modulation of the No-cGMP Pathway Has No Effect on Olfactory Responses in the Drosophila antenna. Front. Cell. Neurosci. 2023, 17, 1180798. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, J. Electrophysiological responses of the oriental fruit fly, Bactrocera dorsalis to host-plant related volatiles. J. Environ. Entomol. 2016, 38, 126–131. [Google Scholar]
- Zheng, W.; Peng, W.; Zhu, C.; Zhang, Q.; Saccone, G.; Zhang, H. Identification and Expression Profile Analysis of Odorant Binding Proteins in the Oriental Fruit Fly Bactrocera dorsalis. Int. J. Mol. Sci. 2013, 14, 14936–14949. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Smagghe, G.; Lei, Z.; Wang, J.J. Identification of Male- and Female-Specific Olfaction Genes in Antennae of the Oriental Fruit Fly (Bactrocera dorsalis). PLoS ONE 2016, 11, e0147783. [Google Scholar] [CrossRef]
- Yi, X.; Wang, P.; Wang, Z.; Cai, J.; Hu, M.; Zhong, G. Involvement of a Specific Chemosensory Protein from Bactrocera dorsalis in Perceiving Host Plant Volatiles. J. Chem. Ecol. 2014, 40, 267–275. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, X.F.; Fu, L.; Han, Y.Y.; Chen, J.; Lu, Y.Y. BdorOBP2 Plays an Indispensable Role in the Perception of Methyl eugenol by Mature Males of Bactrocera dorsalis (Hendel). Sci. Rep. 2017, 7, 15894. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Z.S.; Zhang, D.J.; Lu, Y.Y. BdorOR88a Modulates the Responsiveness to Methyl Eugenol in Mature Males of Bactrocera dorsalis (Hendel). Front. Physiol. 2018, 9, 987. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, D.D.; Wan, L.; Hu, Z.Y.; He, T.T.; Wang, J.B.; Deng, S.Z.; Wang, X.S. Assessment of Attractancy and Safeness of (E)-Coniferyl Alcohol for Management of Female Adults of Oriental Fruit Fly, Bactrocera dorsalis (Hendel). Pest Manag. Sci. 2022, 78, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lei, Y.; Li, H.; Xu, L.; Yang, H.; Wang, J.; Jiang, H. CRISPR/Cas9 Mutagenesis Abolishes Odorant-Binding Protein BdorOBP56f-2 and Impairs the Perception of Methyl Eugenol in Bactrocera dorsalis (Hendel). Insect Biochem. Mol. Biol. 2021, 139, 103656. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, H.; Wu, S.; Zhao, W.; Hao, G.; Wang, J.; Jiang, H. BdorOBP69a Is Involved in the Perception of the Phenylpropanoid Compound Methyl Eugenol in Oriental Fruit Fly (Bactrocera dorsalis) Males. Insect Biochem. Mol. Biol. 2022, 147, 103801. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Jiang, H.B.; Tang, K.Y.; Yan, Y.; Schetelig, M.F.; Wang, J.J. CRISPR-Mediated Mutagenesis of the Odorant Receptor co-Receptor (Orco) Gene Disrupts Olfaction-Mediated Behaviors in Bactrocera dorsalis. Insect Sci. 2022, 29, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Zhou, X.; Gu, F.; Zhong, G.; Yi, X. Olfactory Plasticity: Variation in the Expression of Chemosensory Receptors in Bactrocera dorsalis in Different Physiological States. Front. Physiol. 2017, 8, 672. [Google Scholar] [CrossRef] [PubMed]
- Kah-Wei Hee, A.; Tan, K.H. Transport of Methyl Eugenol-Derived Sex Pheromonal Components in the Male Fruit Fly, Bactrocera dorsalis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 143, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Rasool, A.; Fatima, S.; Shah, S.H.; Munis, M.F.H.; Irshad, A.; Shelly, T.E.; Haq, I.U. Methyl Eugenol Aromatherapy: A Delivery System Facilitating the Simultaneous Application of Male Annihilation and Sterile Insect Technique against the Peach Fruit Fly. Pest Manag. Sci. 2024, 80, 1465–1473. [Google Scholar] [CrossRef]
- Sim, S.B.; Curbelo, K.M.; Manoukis, N.C.; Cha, D.H. Evaluating Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) Response to Methyl Eugenol: Comparison of Three Common Bioassay Methods. J. Econ. Entomol. 2022, 115, 556–564. [Google Scholar] [CrossRef]
- Buczkowski, G. A comparison of Insecticide Susceptibility Levels in 12 Species of urban Pest Ants with Special Focus on the Odorous House Ant, Tapinoma Sessile. Pest Manag. Sci. 2021, 77, 2948–2954. [Google Scholar] [CrossRef] [PubMed]
- Nugnes, F.; Russo, E.; Viggiani, G.; Bernardo, U. First Record of an Invasive Fruit Fly Belonging to Bactrocera dorsalis Complex (Diptera: Tephritidae) in Europe. Insects 2018, 9, 182. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | English Name | CAS | ID | English Name | CAS |
|---|---|---|---|---|---|
| T01 | (4R)-limonene | 5989-27-5 | T44 | Pentolinium | 144-44-5 |
| T02 | Toluene | 108-88-3 | T45 | Ginkgolide-J | 107438-79-9 |
| T03 | 1,3,5-Trimethoxybenzene | 621-23-8 | T46 | Decamethonium | 156-74-1 |
| T04 | Chamazulene | 529-05-5 | T47 | Isoquinoline | 119-65-3 |
| T05 | Alpha-Pinene | 80-56-8 | T48 | Cinnamyl alcohol | 4407-36-7 |
| T06 | Anhydrovitamin A | 1224-78-8 | T49 | Enbucrilate | 6606-65-1 |
| T07 | Calcium undecylenate | 1322-14-1 | T50 | Caprylyl glycol | 1117-86-8 |
| T08 | Farnesol | 4602-84-0 | T51 | Carbazole | 86-74-8 |
| T09 | Spermine | 71-44-3 | T52 | Prifinium | 10236-81-4 |
| T10 | Androstenedione | 63-05-8 | T53 | Lauric acid | 143-07-7 |
| T11 | Terpineol | 8000-41-7 | T54 | MDL72527 | 99207-33-7 |
| T12 | Adamantane | 281-23-2 | T55 | 17alpha-methyl-3beta | 571-03-9 |
| T13 | Verbenone | 80-57-9 | T56 | Boldione | 897-06-3 |
| T14 | Mitemcinal | 154738-42-8 | T57 | Hexamethonium | 60-26-4 |
| T15 | Dodecyltrimethylammonium | 10182-91-9 | T58 | Ginkgolide-C | 15291-76-6 |
| T16 | Anethole | 4180-23-8 | T59 | Cetyltrimethylammonium naproxenate | 102580-74-5 |
| T17 | Levoverbenone | 1196-01-6 | T60 | Benzyl formate | 104-57-4 |
| T18 | 19-norandrostenedione | 734-32-7 | T61 | Hydroxytyrosol | 10597-60-1 |
| T19 | Terpinyl acetate | 8007-35-0 | T62 | Amyl acetate | 628-63-7 |
| T20 | Nonan-1-Ol | 28473-21-4 | T63 | Tetraethylammonium | 66-40-0 |
| T21 | Beta-Pinene | 127-91-3 | T64 | Capric acid | 334-48-5 |
| T22 | Bretylium | 59-41-6 | T65 | Octamethylenediamine | 373-44-4 |
| T23 | Soneclosan | 3380-30-1 | T66 | 4-Androstenediol | 1156-92-9 |
| T24 | Guaiazulen | 489-84-9 | T67 | 5-androstenedione | 571-36-8 |
| T25 | Undecylenic acid | 112-38-9 | T68 | (S)-oct-1-en-3-ol | 24587-53-9 |
| T26 | Levomenol | 23089-26-1 | T69 | Dimethyl carbate | 39589-98-5 |
| T27 | Spermidine | 124-20-9 | T70 | Ginkgolide-M | 15291-78-8 |
| T28 | Geraniol | 106-24-1 | T71 | Quaternium-24 | 32426-11-2 |
| T29 | Terpinen-4-ol | 562-74-3 | T72 | Exemestane | 107868-30-4 |
| T30 | Camphane | 464-15-3 | T73 | Bornyl acetate | 76-49-3 |
| T31 | Fusicoccin | 20108-30-9 | T74 | 17beta-diol | 1852-53-5 |
| T32 | Dioctyldimonium | 20256-55-7 | T75 | Iodobenzene | 591-50-4 |
| T33 | Vanillyl alcohol | 498-00-0 | T76 | Carbaryl | 63-25-2 |
| T34 | 2-octyl cyanoacrylate | 133978-15-1 | T77 | N-Tridecanoic Acid | 638-53-9 |
| T35 | 1-Dodecanol | 112-53-8 | T78 | Agmatine | 306-60-5 |
| T36 | 1,2-dichlorobenzene | 95-50-1 | T79 | Bolandiol | 19793-20-5 |
| T37 | Triclosan | 3380-34-5 | T80 | Atamestane | 96301-34-7 |
| T38 | Diphemanil | 15394-62-4 | T81 | Dibromothymoquinone | 29096-93-3 |
| T39 | Palmitoleic Acid | 373-49-9 | T82 | Midecamycin | 35457-80-8 |
| T40 | Perillyl alcohol | 18457-55-1 | T83 | Cetrimonium | 6899-10-1 |
| T41 | Bis(6-aminohexyl)amine | 143-23-7 | T84 | Benzyl benzoate | 120-51-4 |
| T42 | Progesterone | 57-83-0 | T85 | CA4P | 222030-63-9 |
| T43 | Duroquinone | 527-17-3 | T86 | Isopentyl 2-cyanoacrylate | 19475-26-4 |
| Compound ID | Compound Name | CAS | Average Affinity (kcal/mol) | Degree |
|---|---|---|---|---|
| T70 | Ginkgolide-M | 15291-78-8 | −10.12 | 107 |
| T14 | Mitemcinal | 154738-42-8 | −8.77 | 107 |
| T82 | Midecamycin | 35457-80-8 | −6.69 | 106 |
| T72 | Exemestane | 107868-30-4 | −6.59 | 105 |
| T55 | 17alpha-methyl-3beta | 571-03-9 | −6.55 | 105 |
| T56 | Boldione | 897-06-3 | −6.50 | 105 |
| T67 | 5-androstenedione | 571-36-8 | −6.42 | 105 |
| T42 | Progesterone | 57-83-0 | −6.38 | 104 |
| T80 | Atamestane | 96301-34-7 | −6.51 | 104 |
| T18 | 19-norandrostenedione | 734-32-7 | −6.44 | 104 |
| T74 | 17beta-diol | 1852-53-5 | −6.52 | 103 |
| T10 | Androstenedione | 63-05-8 | −6.51 | 103 |
| T79 | Bolandiol | 19793-20-5 | −6.49 | 102 |
| T06 | Anhydrovitamin A | 1224-78-8 | −6.25 | 101 |
| T66 | 4-Androstenediol | 1156-92-9 | −6.48 | 101 |
| T38 | Diphemanil | 15394-62-4 | −6.20 | 101 |
| T24 | Guaiazulen | 489-84-9 | −6.39 | 101 |
| T04 | Chamazulene | 529-05-5 | −6.24 | 98 |
| T51 | Carbazole | 86-74-8 | −6.18 | 96 |
| T52 | Prifinium | 10236-81-4 | −6.06 | 92 |
| Concentration | Number of B. dorsalis Mean ± SE | Male Proportion |
|---|---|---|
| 1 mg/mL Exemestane | 3.60 ± 0.49 ab | 22.22% |
| 0.1 mg/mL Exemestane | 7.80 ± 0.98 bc | 28.20% |
| 0.01 mg/mL Exemestane | 4.60 ± 0.80 ab | 24.78% |
| 1 mg/mL Progesterone | 11.00 ± 1.26 c | 70.90% |
| 0.1 mg/mL Progesterone | 3.60 ± 0.49 ab | 55.55% |
| 0.01 mg/mL Progesterone | 3.40 ± 0.49 ab | 58.82% |
| 1 mg/mL 19-norandrostenedione | 8.00 ± 0.40 bc | 95.00% |
| 0.1 mg/mL 19-norandrostenedione | 8.60 ± 1.01 bc | 93.02% |
| 0.01 mg/mL 19-norandrostenedione | 2.40 ± 0.49 a | 100.00% |
| DMSO Solvent (0 mg/mL) | 4.40 ± 0.49 ab | 45.00% |
| Empty bottle | 2.20 ± 0.40 a | 45.00% |
| 1 mg/mL ME | 11.40 ± 1.01 c | 85.96% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Cao, F.; Zou, Y.; Li, Y.; Wen, J.; Fu, Y.; Su, H.; Bai, X.; Xie, X.; Zeng, L.; et al. The Discovery of the Potential Attractive Compounds of Bactrocera dorsalis (Hendel). Horticulturae 2024, 10, 299. https://doi.org/10.3390/horticulturae10030299
Chen Y, Cao F, Zou Y, Li Y, Wen J, Fu Y, Su H, Bai X, Xie X, Zeng L, et al. The Discovery of the Potential Attractive Compounds of Bactrocera dorsalis (Hendel). Horticulturae. 2024; 10(3):299. https://doi.org/10.3390/horticulturae10030299
Chicago/Turabian StyleChen, Yupeng, Fengqin Cao, Yan Zou, Yi Li, Jian Wen, Yu Fu, Hongai Su, Xue Bai, Xiaowei Xie, Ling Zeng, and et al. 2024. "The Discovery of the Potential Attractive Compounds of Bactrocera dorsalis (Hendel)" Horticulturae 10, no. 3: 299. https://doi.org/10.3390/horticulturae10030299
APA StyleChen, Y., Cao, F., Zou, Y., Li, Y., Wen, J., Fu, Y., Su, H., Bai, X., Xie, X., Zeng, L., Liang, G., Wang, D., & Lu, Y. (2024). The Discovery of the Potential Attractive Compounds of Bactrocera dorsalis (Hendel). Horticulturae, 10(3), 299. https://doi.org/10.3390/horticulturae10030299