

Effect of UV Stress on the Antioxidant Capacity, Photosynthetic Activity, Flavonoid and Steviol Glycoside Accumulation of Stevia rebaudiana Bertoni

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cultivation Conditions
2.2. Determination of Biometric Indicators
2.3. Measurement of Antioxidant Activity
2.4. Determination of the Total Content of Flavonoids in Terms of Quercetin
2.5. Quantification of SG
2.6. Measuring Leaf Gas Exchange and Chlorophyll Fluorescence
2.7. Measuring Vegetation Indices
2.8. Data Analysis
3. Results and Discussion
3.1. The Influence of Additional UV Irradiation on the Morphological Parameters of the Stevia Plant
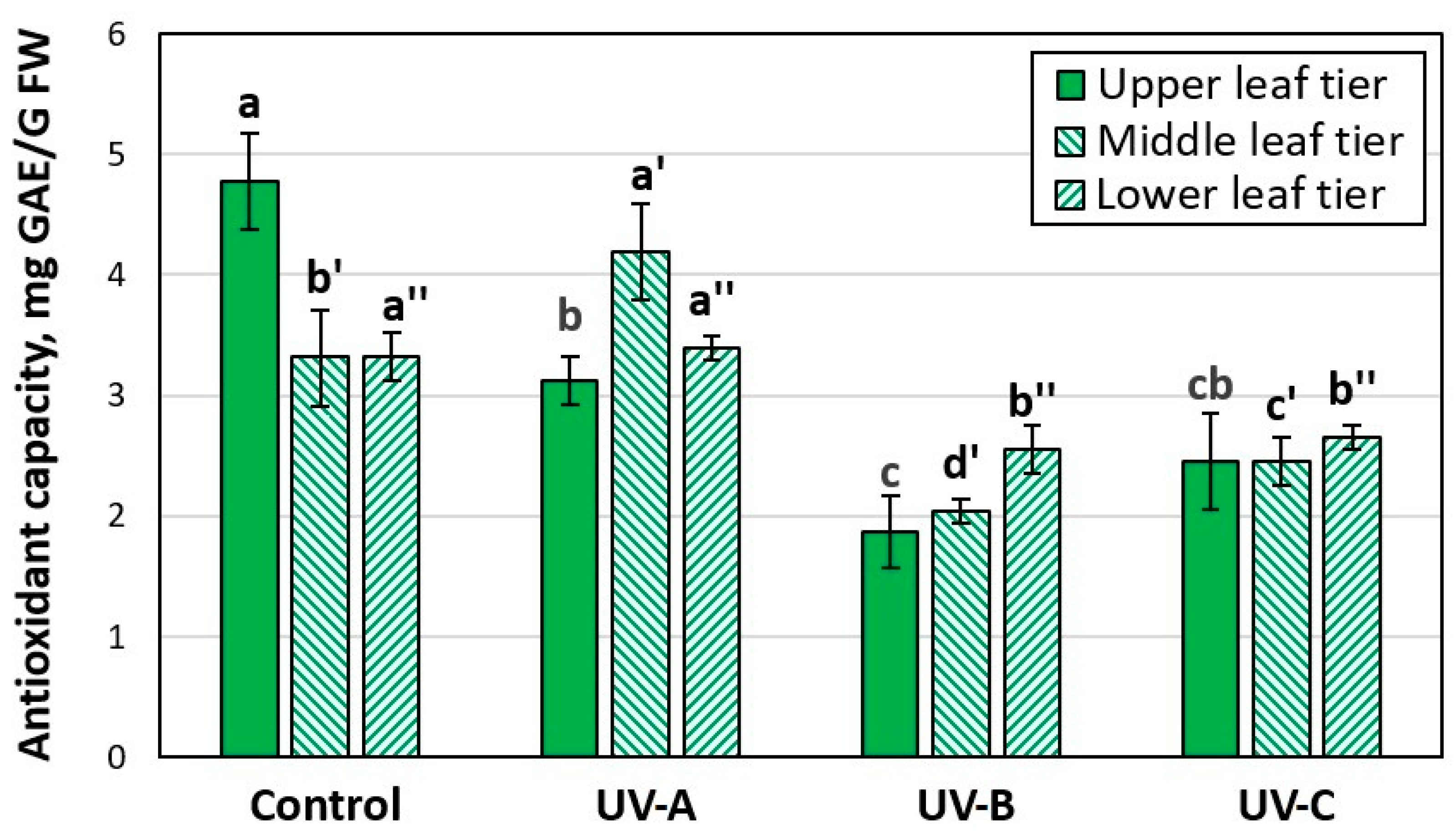
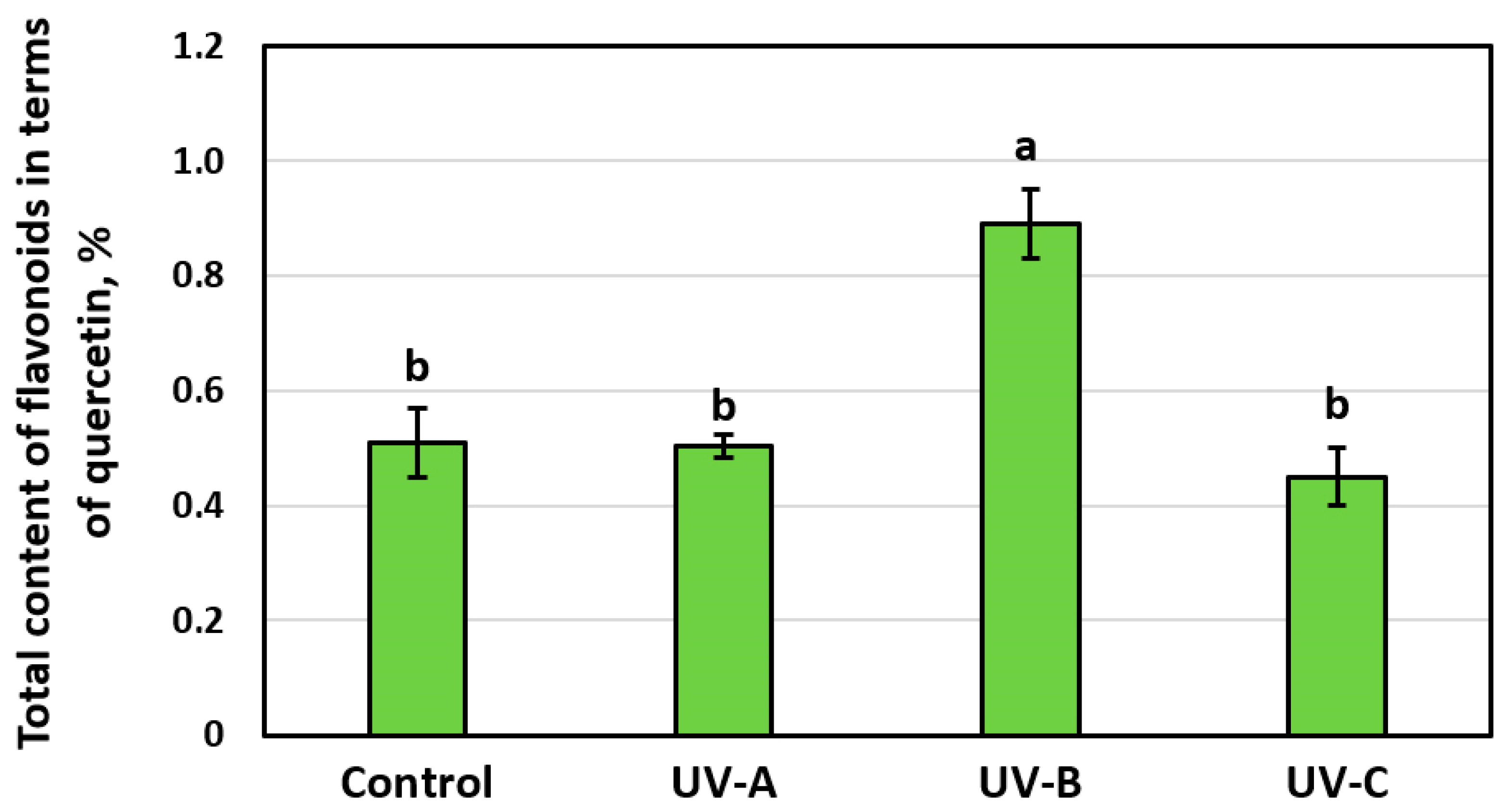
3.2. Effect of Additional UV Irradiation on Antioxidant Activity and Accumulation of SG Flavonoids in the Leaves of the Stevia Plant
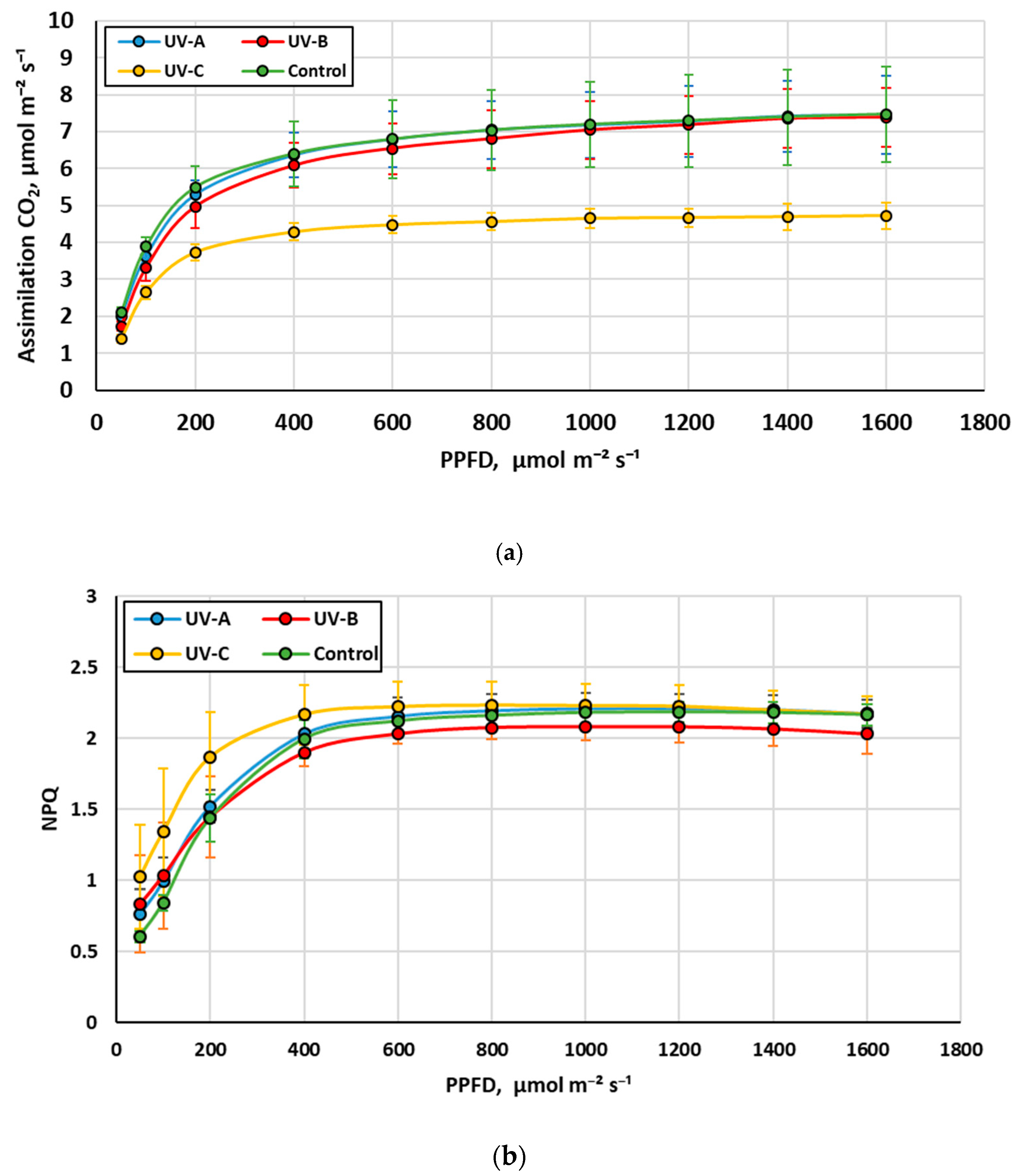
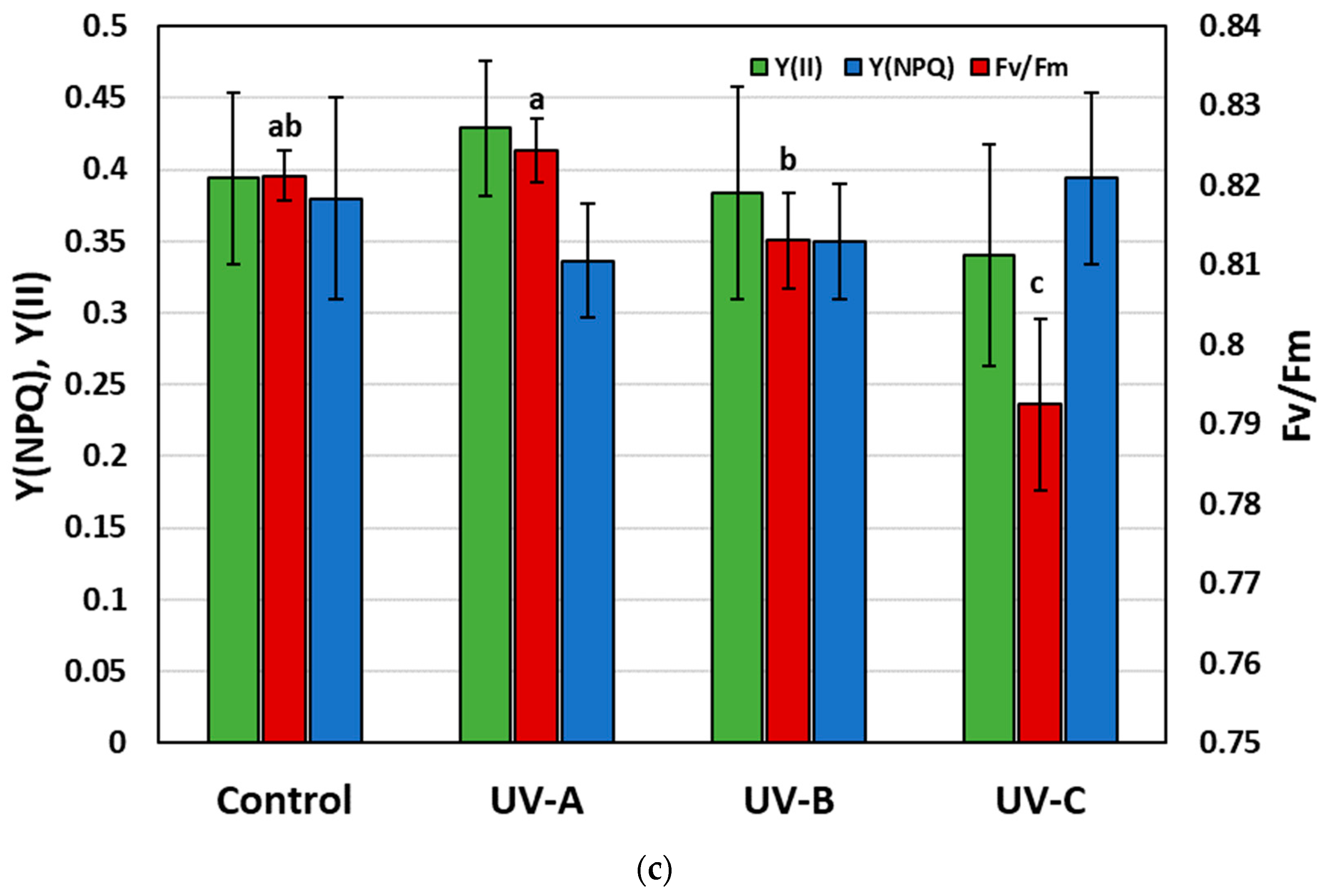
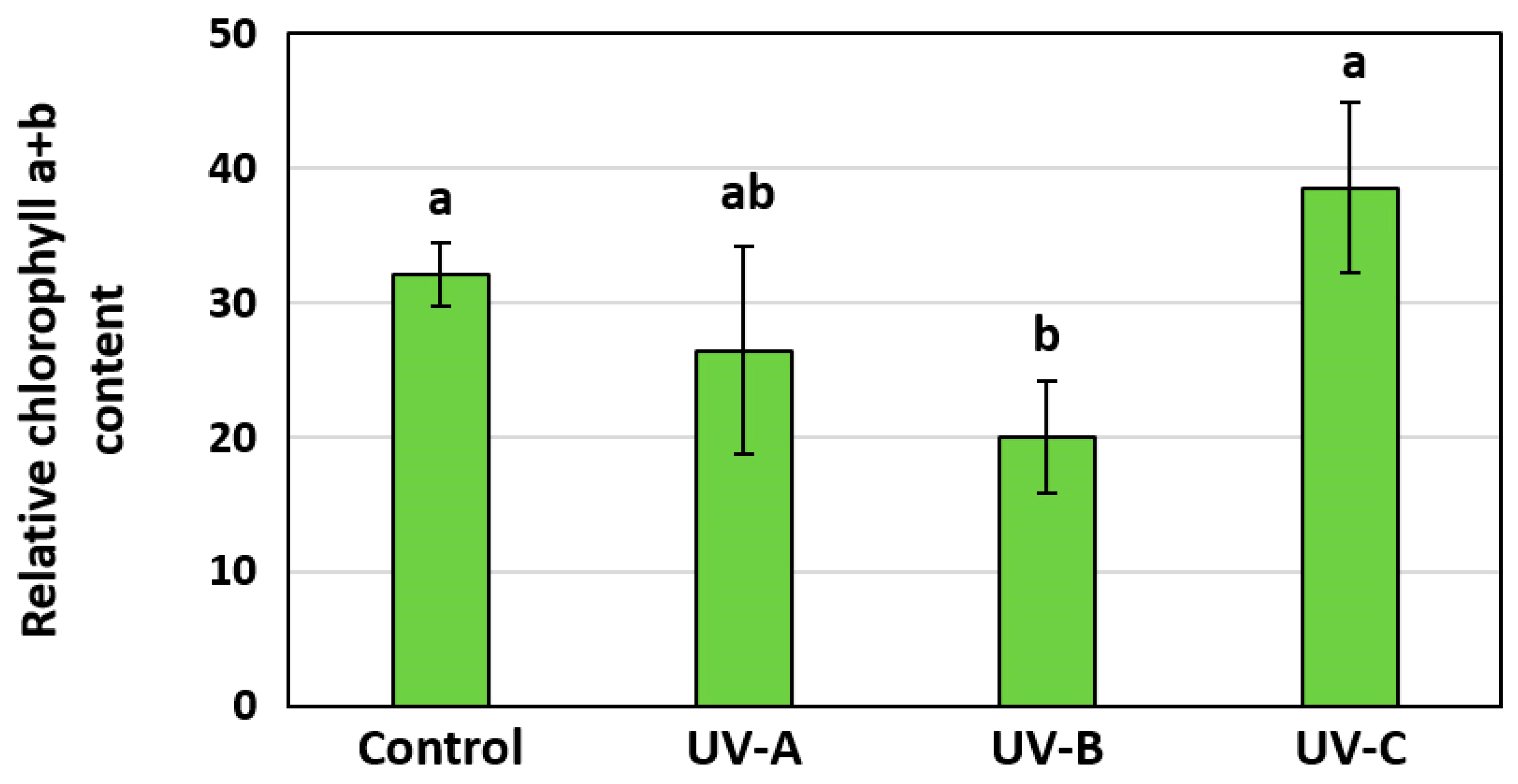
3.3. The Influence of Additional UV Irradiation on the Stress Level, Net Assimilation CO2, and Chlorophyll Fluorescence of Stevia Plants
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Saraiva, A.; Carrascosa, C.; Raheem, D.; Ramos, F.; Raposo, A. Natural sweeteners: The relevance of food naturalness for consumers, food security aspects, sustainability and health impacts. Int. J. Environ. Res. Public Health 2020, 17, 6285. [Google Scholar] [CrossRef]
- Koyama, E.; Kitazawa, K.; Ohori, Y.; Izawa, O.; Kakegawa, K.; Fujino, A.; Ui, M. In vitro metabolism of the glycosidic sweeteners, stevia mixture and enzymatically modified stevia in human intestinal microflora. Food Chem. Toxicol 2003, 41, 359–374. [Google Scholar] [CrossRef]
- Bondarev, N.I.; Sukhanova, M.A.; Reshetnyak, O.V.; Nosov, A.M. Steviol glycoside content in different organs of Stevia rebaudiana and its dynamics during ontogeny. Biol. Plant 2003, 47, 261–264. [Google Scholar] [CrossRef]
- Muanda, F.N.; Soulimani, R.; Diop, B.; Dicko, A. Study on chemical composition and biological activities of essential oil and extracts from Stevia rebaudiana Bertoni leaves. LWT 2011, 44, 1865–1872. [Google Scholar] [CrossRef]
- Santini, A.; Ferrara, L.; Naviglio, D.; Aragon, A.; Ritieni, A. Stevia: Non-caloric natural sweetener. Prog. Nutr. 2008, 10, 3–10. [Google Scholar]
- Sharma, S.; Walia, S.; Singh, B.; Kumar, R. Comprehensive review on agro technologies of low-calorie natural sweetener stevia (Stevia rebaudiana Bertoni): A boon to diabetic patients. J. Sci. Food Agric. 2016, 96, 1867–1879. [Google Scholar] [CrossRef] [PubMed]
- Obst, K.; Ley, J.P.; Reichelt, K.V.; Krammer, G.E.; Engel, K.H. Sensory. Characterisation of Stevia Extracts. Stevia: Science, no fiction. In Proceedings of the 4th Stevia Symposium of the European-Stevia-Association—EUSTAS Symposium, Leuven, Belgium, 29–30 June 2010; pp. 121–134. [Google Scholar]
- Scholten, C.; Haji, F. Methods to influence the taste and sweetness profile of Stevia. Sensory Characterisation of Stevia Extracts. Stevia: Science, no fiction. In Proceedings of the 4th Stevia Symposium of the European-Stevia-Association—EUSTAS Symposium, Leuven, Belgium, 29–30 June 2010; pp. 111–120. [Google Scholar]
- Bergman, M.E.; Davis, B.; Phillips, M.A. Medically useful plant terpenoids: Biosynthesis, occurrence, and mechanism of action. Molecules 2019, 24, 3961. [Google Scholar] [CrossRef]
- Chen, T.H.; Chen, S.C.; Chan, P.; Chu, Y.; Yang, H.Y.; Cheng, J.T. Mechanism of the hypoglycemic effect of stevioside, a glycoside of Stevia rebaudiana. Planta Med. 2005, 71, 108–113. [Google Scholar] [CrossRef]
- Samuel, P.; Ayoob, K.; Magnuson, B.; Wölwer-Rieck, U.; Jeppesen, P.; Rogers, P.; Rowland, I.; Mathews, R. Stevia Leaf to Stevia Sweetener: Exploring Its Science, Benefits, and Future Potential. J. Nutr. 2018, 148, 1186S–1205S. [Google Scholar] [CrossRef]
- Jeppesen, P.; Gregersen, S.; Alstrup, K.; Hermansen, K. Stevioside induces antihyperglycaemic, insulinotropic and glucagonostatic effects in vivo: Studies in the diabetic Goto-Kakizaki (GK) rats. Phytomedicine 2002, 9, 9–14. [Google Scholar] [CrossRef]
- Starratt, A.N.; Kirby, C.W.; Pocs, R.; Brandle, J.E.; Rebaudioside, F.A. Diterpene glycoside from Stevia rebaudiana. Phytochemistry 2002, 59, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Tadhani, M.B.; Patel, V.H.; Subhash, R. In vitro antioxidant activities of Stevia rebaudiana leaves and callus. J. Food Compos. Anal 2007, 20, 323–329. [Google Scholar] [CrossRef]
- Chan, P.; Tomlinson, B.; Chen, Y.J.; Liu, J.C.; Hsieh, M.H.; Cheng, J.T. A double-blind placebo-controlled study of the effectiveness and tolerability of oral stevioside in human hypertension. Br. J. Clin. Pharmacol. 2000, 50, 215–220. [Google Scholar] [CrossRef]
- Yasukawa, K.; Kitanaka, S.; Seo, S. Inhibitory effect of stevioside on tumor promotion by 12-O-tetradecanoylphorbol-13-acetate in two-stage carcinogenesis in mouse skin. Biol. Pharm. Bull. 2002, 25, 1488–1490. [Google Scholar] [CrossRef]
- Papaefthimiou, M.; Kontou, P.I.; Bagos, P.G.; Braliou, G.G. Antioxidant Activity of Leaf Extracts from Stevia rebaudiana Bertoni Exerts Attenuating Effect on Diseased Experimental Rats: A Systematic Review and Meta-Analysis. Nutrients 2023, 15, 3325. [Google Scholar] [CrossRef]
- Gupta, E.; Purwar, S.; Sundaram, S.; Rai, G. Nutritional and therapeutic values of Stevia rebaudiana: A review. J. Med. Plants Res. 2013, 7, 3343–3353. [Google Scholar]
- Panpatil, V.; Polasa, K. Assessment of stevia (Stevia rebaudiana)-natural sweetener: A review. J. Food Sci. Technol. 2008, 45, 467–473. [Google Scholar]
- Talevi, A. Potential medicinal effects and applications of stevia constituents. Phytochem. Rev. 2022, 21, 161–178. [Google Scholar] [CrossRef]
- Khandaker, M.M.; Liyana, N.; Majrashi, A.; Dalorima, T.; Alias, N.; Mat, N. Effects of kinetin on the morpho-physiological and biochemical characteristics of stevia (Stevia rebaudiana). Bulg. J. Agric. Sci. 2018, 24, 638–647. [Google Scholar]
- Ucar, E.; Ozyigit, Y.; Eruygur, N.; Güven, D.; Yur, S.; Turgut, K.; Özek, T.; Kütük, N. The Effect of the Plant Age and Growth Period on the Nutritional Substance, Chlorophyll and Steviol Glycoside Rates in Stevia (Stevia Rebaudiana Bertoni) Leaves. Commun. Soil Sci. Plant Anal. 2018, 49, 291–302. [Google Scholar] [CrossRef]
- Samsulrizal, N.H.; Khadzran, K.S.; Shaarani, S.H.; Noh, A.L.; Sundram, T.C.; Naim, M.A.; Zainuddin, Z. De novo transcriptome dataset of Stevia rebaudiana accession MS007. Data Brief 2019, 28, 1–3. [Google Scholar] [CrossRef]
- Tarakanov, I.G. Light control of growth and development in vegetable plants with various life strategies. Acta Hortic. 2006, 711, 315–321. [Google Scholar] [CrossRef]
- Tarakanov, I.; Yakovleva, O.; Konovalova, I.; Paliutina, G.; Anisimov, A. Light-emitting diodes: On the way to combinatorial lighting technologies for basic research and crop production. Acta Hortic. 2012, 956, 171–178. [Google Scholar] [CrossRef]
- Nakonechnaya, O.V.; Gafitskaya, I.V.; Burkovskaya, E.V.; Khrolenko, Y.A.; Grishchenko, O.V.; Zhuravlev, Y.N.; Sussotin, E.P.; Kulchin, Y.N. Effect of light intensity on the morphogenesis of Stevia rebaudiana under in vitro conditions. Russ. J. Plant Physiol. 2019, 66, 656–663. [Google Scholar] [CrossRef]
- Muthu, S.; Schuurmans, F.J.; Pashley, M.D. Red, green, and blue LEDs for white light illumination. IEEE J. Sel. Top. Quantum. Electron. 2002, 8, 333–338. [Google Scholar] [CrossRef]
- Rivera-Avilez, J.A.; Jarma-Orozco, A.; Pompelli, M.F. Stevia rebaudiana Bertoni: The Interaction of Night Interruption on Gas Exchange, Flowering Delay, and Steviol Glycosides Synthesis. Horticulturae 2021, 7, 543. [Google Scholar] [CrossRef]
- Stijn, C.; Stefaan, W.; Geuns, J. Stimulation of steviol glycoside accumulation in Stevia rebaudiana by red LED light. J. Plant Physiol. 2012, 169, 749–752. [Google Scholar]
- Abdulameer, D.; Osman, M.; Sulaiman, Z.; Yusop, M.; Abdullah, S.; Azizi, P.; Muttaleb, Q. Assessment of Stevia rebaudiana Bertoni Genotypes via Morpho-Agronomic Traits under Two Light Conditions. Am. J. Plant Sci. 2018, 9, 1403–1413. [Google Scholar] [CrossRef][Green Version]
- Bondarev, N.I.; Reshetnyak, O.V.; Nosov, A.M. Influence of photoperiod and irradiation intensity on the development of Stevia rebaudiana shoots in vitro and synthesis of steviol glycosides. Izv. TSKhA 2008, 4, 102–107. [Google Scholar]
- Ahmad, N.; Rab, A.; Ahmad, N. Light-induced biochemical variations in secondary metabolite production and antioxidant activity in callus cultures of Stevia rebaudiana (Bert). J. Photochem. Photobiol. 2016, 154, 6–51. [Google Scholar] [CrossRef]
- Yoneda, Y.; Nakashima, H.; Miyasaka, J.; Ohdoi, K.; Shimizu, H.J.P. Impact of blue, red, and far-red light treatments on gene 1093 expression and steviol glycoside accumulation in Stevia rebaudiana. Phytochemistry 2017, 137, 57–65. [Google Scholar] [CrossRef]
- Shevchenko, Y.; Wendt, A.; Smetanska, I. Sprout culture of Stevia rebaudiana Bertoni. In Proceedings of the 4th Stevia Symposium of the European-Stevia-Association—EUSTAS Symposium. Stevia: Science, Leuven, Belgium, 29–30 June 2010; pp. 5–26. [Google Scholar]
- Rengasamy, N.; Othman, R.Y.; Che, H.S.; Harikrishna, J.A. Beyond the PAR spectra: Impact of light quality on the germination, flowering, and metabolite content of Stevia rebaudiana (Bertoni). J. Sci. Food Agric. 2021, 102, 299–311. [Google Scholar] [CrossRef]
- Ceunen, S.; Geuns, J.M.J.P. Spatio-temporal variation of the diterpene steviol in Stevia rebaudiana grown under different photoperiods. Phytochemistry 2013, 89, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Kovylyaeva, G.I.; Bakaleinik, G.A.; Strobykina, I.Y.; Gubskaya, V.I.; Sharipova, R.R.; Al’Fonsov, V.A.; Kataev, V.E.; Tolstikov A., G. Glycosides from Stevia rebaudiana. Chem. Nat. Compd. 2007, 43, 81–85. [Google Scholar] [CrossRef]
- Denev, P.N.; Uchkunov, I.I.; Uchkunov, V.I.; Kratchanova, M.G. Content of steviol glycosides in stevia (Stevia Rebaudiana B.) genotypes cultivated in Bulgaria. Bulg. Chem. Commun. 2017, 6, 121–125. [Google Scholar]
- Lee, M.-J.; Son, J.E.; Oh, M.-M. Growth and phenolic compounds of Lactuca sativa L. grown in a closed-type plant production system with UV-A, -B, or -C lamp. J. Sci. Food Agric. 2014, 94, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.A.; Tungmunnithum, D.; Garros, L.; Drouet, S.; Hano, C.; Abbasi, B.H. Effect of Ultraviolet-C Radiation and Melatonin Stress on Biosynthesis of Antioxidant and Antidiabetic Metabolites Produced in In Vitro Callus Cultures of Lepidium sativum L. Int. J. Mol. Sci. 2019, 20, 1787. [Google Scholar] [CrossRef] [PubMed]
- Nazir, M.; Asad Ullah, M.; Mumtaz, S.; Siddiquah, A.; Shah, M.; Drouet, S.; Hano, C.; Abbasi, B.H. Interactive Effect of Melatonin and UV-C on Phenylpropanoid Metabolite Production and Antioxidant Potential in Callus Cultures of Purple Basil (Ocimum basilicum L. var purpurascens). Molecules 2020, 25, 1072. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Rao, G.J.S.T. Stevia: The herbal sugar of 21st century. Sugar Tech 2005, 7, 17–24. [Google Scholar] [CrossRef]
- Huxley, A.; Griffiths, M.; Levy, M. The New RHS Dictionary of Gardening; Royal Horticultural Society: London, UK, 1992; pp. 315–318. [Google Scholar]
- Gins, M.S.; Gins, W.K.; Baykov, A.A.; Kononkov, P.F.; Romanova, E.V. Methodology for analyzing the total content of antioxidants in leaf and leafy vegetable crops. In Educational Manual; RUDN Publ.: Moscow, Russia, 2013; p. 40. [Google Scholar]
- Fedoseeva, G.M.; Mirovich, V.M.; Goryachkina, E.G.; Perelomova, M.V. Phytochemical analysis of plant raw materials containing flavonoids. In Methodological Manual on Pharmacognosy; ISMU Publ.: Irkutsk, Russia, 2009; p. 67. [Google Scholar]
- Semenova, N.A.; Smirnov, A.A.; Ivanitskikh, A.S.; Izmailov, A.Y.; Dorokhov, A.S.; Proshkin, Y.A.; Yanykin, D.V.; Sarimov, R.R.; Gudkov, S.V.; Chilingaryan, N.O. Impact of UV radiation on the biosynthesis of pigments and essential oils of sweet basil (Ocimum basilicum L.). Appl. Sci. 2022, 12, 7190. [Google Scholar] [CrossRef]
- Hao, J.; Lou, P.; Han, Y.; Zheng, L.; Lu, J.; Chen, Z.; Ni, J.; Yang, Y.; Xu, M. Ultraviolet-B Irradiation Increases Antioxidant Capacity of Pakchoi (Brassica rapa L.) by Inducing Flavonoid Biosynthesis. Plants 2022, 11, 766. [Google Scholar] [CrossRef]
- Kootstra, A. Protection from UV-B-induced DNA damage by flavonoids. Plant Mol. Biol. 1994, 26, 771–774. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Pollastri, S.; Tattini, M. Functional roles of flavonoids in photoprotection: New evidence, lessons from the past. Plant Physiol. Biochem. 2013, 72, 35–45. [Google Scholar] [CrossRef]
- Agati, G.; Biricolti, S.; Guidi, L.; Ferrini, F.; Fini, A.; Tattini, M. The biosynthesis of flavonoids is enhanced similarly by UV radiation and root zone salinity in L. vulgare leaves. J. Plant Physiol. 2011, 168, 204–212. [Google Scholar] [CrossRef]
- Scattino, C.; Castagna, A.; Neugart, S.; Chan, H.; Schreiner, M.; Crisosto, C.; Tonutti, P.; Ranieri, A. Post-harvest UV-B irradiation induces changes of phenol contents and corresponding biosynthetic gene expression in peaches and nectarines. Food Chem. 2014, 163, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Ghasemzadeh, A.; Ashkani, S.; Baghdadi, A.; Pazoki, A.; Jaafar, H.; Rahmat, A. Improvement in Flavonoids and Phenolic Acids Production and Pharmaceutical Quality of Sweet Basil (Ocimum basilicum L.) by Ultraviolet-B Irradiation. Molecules 2016, 21, 1203. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hu, L.; Jiang, D.; Xi, W. Effect of Post-Harvest LED and UV Light Irradiation on the Accumulation of Flavonoids and Limonoids in the Segments of Newhall Navel Oranges (Citrus sinensis Osbeck). Molecules 2019, 24, 1755. [Google Scholar] [CrossRef] [PubMed]
- Kohzuma, K.; Tamaki, M.; Hikosaka, K. Corrected photochemical reflectance index (PRI) is an effective tool for detecting environmental stresses in agricultural crops under light conditions. J. Plant Res. 2021, 134, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Garbulsky, M.F.; Peñuelas, J.; Gamon, J.; Inoue, Y.; Filella, I. The photochemical reflectance index (PRI) and the remote sensing of leaf, canopy and ecosystem radiation use efficiencies: A review and meta-analysis. Remote Sens. Environ. 2011, 115, 281–297. [Google Scholar] [CrossRef]
- Guidi, L.; Brunetti, C.; Fini, A.; Agati, G.; Ferrini, F.; Gori, A.; Tattini, M. UV radiation promotes flavonoid biosynthesis, while negatively affecting the biosynthesis and the de-epoxidation of xanthophylls: Consequence for photoprotection. Environ. Exp. Bot. 2016, 127, 14–25. [Google Scholar] [CrossRef]
- Hosseini, S.; Carapetian, J.; Khara, J. The effects of UV radiation on some structural and ultrastructural parameters in pepper (Capsicum longum A. DC.). Turk. J. Biol. 2011, 35, 69–77. [Google Scholar]
- Peguero-Pina, J.J.; Morales, F.; Flexas, J.; Gil-Pelegrín, E.; Moya, I. Photochemistry, remotely sensed physiological reflectance index and de-epoxidation state of the xanthophyll cycle in Quercus coccifera under intense drought. Oecologia 2008, 156, 1–11. [Google Scholar] [CrossRef]
- Urban, L.; Chabane Sari, D.; Orsal, B.; Lopes, M.; Miranda, R.; Aarrouf, J. UV-C light and pulsed light as alternatives to chemical and biological elicitors for stimulating plant natural defenses against fungal diseases. Sci. Hortic. 2018, 235, 452–459. [Google Scholar] [CrossRef]
- Darras, A.I.; Tsikaloudakis, G.; Lycoskoufis, I.; Dimitriadis, C.; Karamousantas, D. Low doses of UV-C irradiation affects growth, fruit yield and photosynthetic activity of tomato plants. Sci. Hortic. 2020, 267, 109357. [Google Scholar]
- Darras, A.; Vlachodimitropoulou, A.; Dimitriadis, C. Regulation of corm sprouting, growth and flowering of pot Freesia hybrida L. plants by cold and UV-C irradiation forcing. Sci. Hortic. 2019, 252, 110–112. [Google Scholar] [CrossRef]
- Vàsquez, H.; Ouhibi, C.; Lizzi, Y.; Azzouz, N.; Forges, M.; Bardin, M.; Nicot, P.; Urban, L.; Aarrouf, J. Pre-harvest hormetic doses of UV-C radiation can decrease susceptibility of lettuce leaves (Lactuca sativa L.) to Botrytis cinerea L. Scientia. Horticulturae 2017, 222, 32–39. [Google Scholar] [CrossRef]
- Xu, Y.; Charles, M.; Luo, Z.; Roussel, D.; Rolland, D. Potential link between fruit yield, quality parameters and phytohormonal changes in preharvest UV-C treated strawberry. Plant Physiol. Biochem. 2017, 116, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, I.; Crizel, G.; Severo, J.; Renard, C.; Chaves, F.C.; Rombaldi, C.V. Preharvest UV-C radiation influences physiological, biochemical, and transcriptional changes in strawberry cv. Camarosa. Plant Physiol. Biochem. 2016, 108, 391–399. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Light Variant | Photon Flux Density (µmol Photons·m−2·s−1) | ||||||
|---|---|---|---|---|---|---|---|
| Blue Light | Green Light | Red Light | Far Red Light | UV-C | UV-B | UV-A | |
| Control | 20 | 44 | 86 | 9 | - | - | - |
| UV-A | 20 | 44 | 86 | 9 | - | - | 1.5 |
| UV-B | 20 | 44 | 86 | 9 | - | 0.015 | - |
| UV-C | 20 | 44 | 86 | 9 | 0.010 | - | - |
| Growth Light Variant | Plant Height (cm) | Fresh Weight (g) | Dry Weight/Fresh Weight (%) | Leaf Surface Area (cm2) | |
|---|---|---|---|---|---|
| Stems | Leaves | ||||
| Control | 60.6 ± 3.5 b | 16.7 ± 4.3 ab | 28.2 ± 4.5 b | 18.0 ± 1.2 a | 1225.4 ± 123.0 b |
| UV-A | 68.3 ± 4.6 b | 20.8 ± 5.1 b | 31.7 ± 4.8 b | 17.5 ± 1.6 a | 1597.4 ± 138.3 c |
| UV-B | 63.8 ± 2.8 b | 20.0 ± 5.3 b | 32.1 ± 5.1 b | 19.3 ± 1.3 a | 1544.9 ± 152.2 c |
| UV-C | 49.0 ± 4.2 a | 11.4 ± 3.8 a | 11.6 ± 6.9 a | 28.6 ± 2.5 b | 542.9 ± 225.0 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semenova, N.A.; Ivanitskikh, A.S.; Uyutova, N.I.; Smirnov, A.A.; Proshkin, Y.A.; Burynin, D.A.; Kachan, S.A.; Sokolov, A.V.; Dorokhov, A.S.; Chilingaryan, N.O. Effect of UV Stress on the Antioxidant Capacity, Photosynthetic Activity, Flavonoid and Steviol Glycoside Accumulation of Stevia rebaudiana Bertoni. Horticulturae 2024, 10, 210. https://doi.org/10.3390/horticulturae10030210
Semenova NA, Ivanitskikh AS, Uyutova NI, Smirnov AA, Proshkin YA, Burynin DA, Kachan SA, Sokolov AV, Dorokhov AS, Chilingaryan NO. Effect of UV Stress on the Antioxidant Capacity, Photosynthetic Activity, Flavonoid and Steviol Glycoside Accumulation of Stevia rebaudiana Bertoni. Horticulturae. 2024; 10(3):210. https://doi.org/10.3390/horticulturae10030210
Chicago/Turabian StyleSemenova, Natalia A., Alina S. Ivanitskikh, Nadezhda I. Uyutova, Alexander A. Smirnov, Yuri A. Proshkin, Dmitry A. Burynin, Sergey A. Kachan, Alexander V. Sokolov, Alexey S. Dorokhov, and Narek O. Chilingaryan. 2024. "Effect of UV Stress on the Antioxidant Capacity, Photosynthetic Activity, Flavonoid and Steviol Glycoside Accumulation of Stevia rebaudiana Bertoni" Horticulturae 10, no. 3: 210. https://doi.org/10.3390/horticulturae10030210
APA StyleSemenova, N. A., Ivanitskikh, A. S., Uyutova, N. I., Smirnov, A. A., Proshkin, Y. A., Burynin, D. A., Kachan, S. A., Sokolov, A. V., Dorokhov, A. S., & Chilingaryan, N. O. (2024). Effect of UV Stress on the Antioxidant Capacity, Photosynthetic Activity, Flavonoid and Steviol Glycoside Accumulation of Stevia rebaudiana Bertoni. Horticulturae, 10(3), 210. https://doi.org/10.3390/horticulturae10030210