
A Comparison of Watermelon Flesh Texture across Different Ploidy Levels Using Histology and Cell Wall Measurements
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Physiological and Biochemical Indicators and Measurement Methods
2.2.1. Evaluation of Fruit Weight, Rind Hardness, and Flesh TSS Content
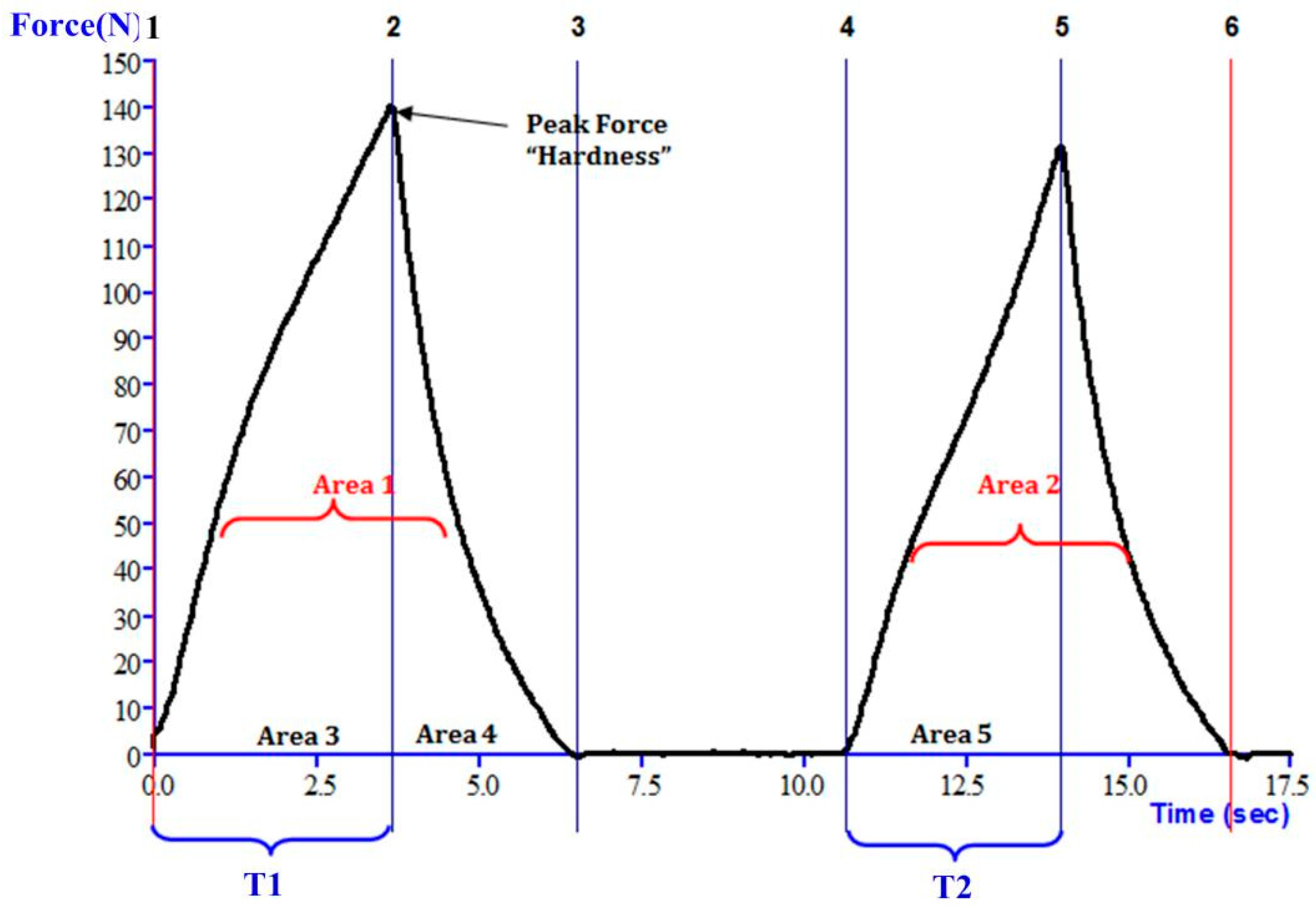
2.2.2. Flesh Texture Measurement
2.2.3. Determination of Cell Wall Polysaccharide Content
2.2.4. Observation of Flesh Cell Microstructure
2.3. Statistical Analysis of Data
3. Results
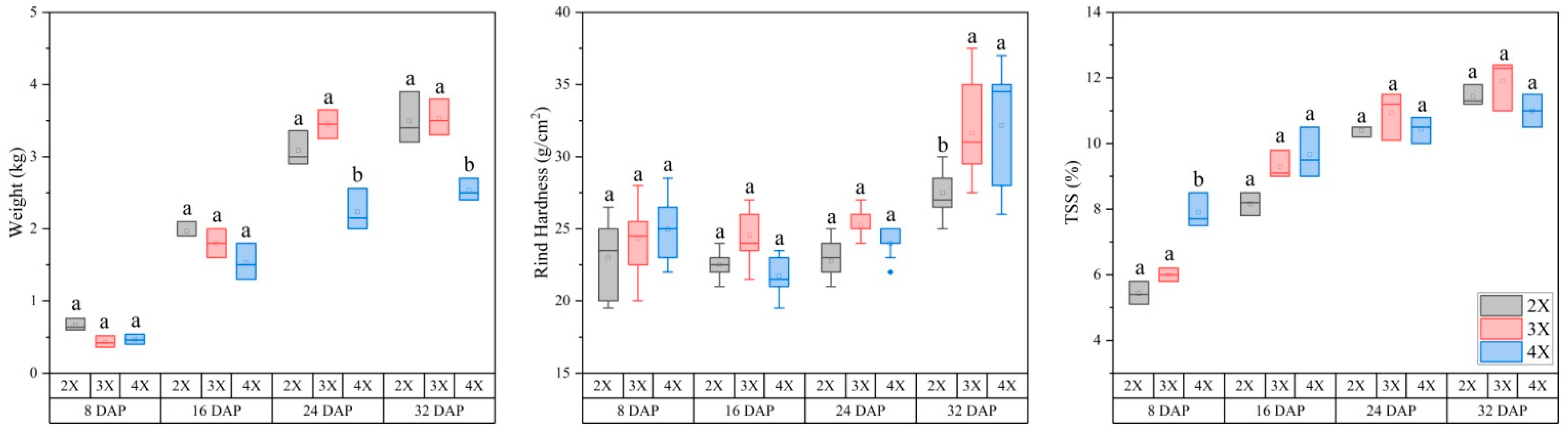
3.1. Differences in Fruit Weight, Rind Hardness, and TSS Content among Different Ploidy Levels of Watermelon
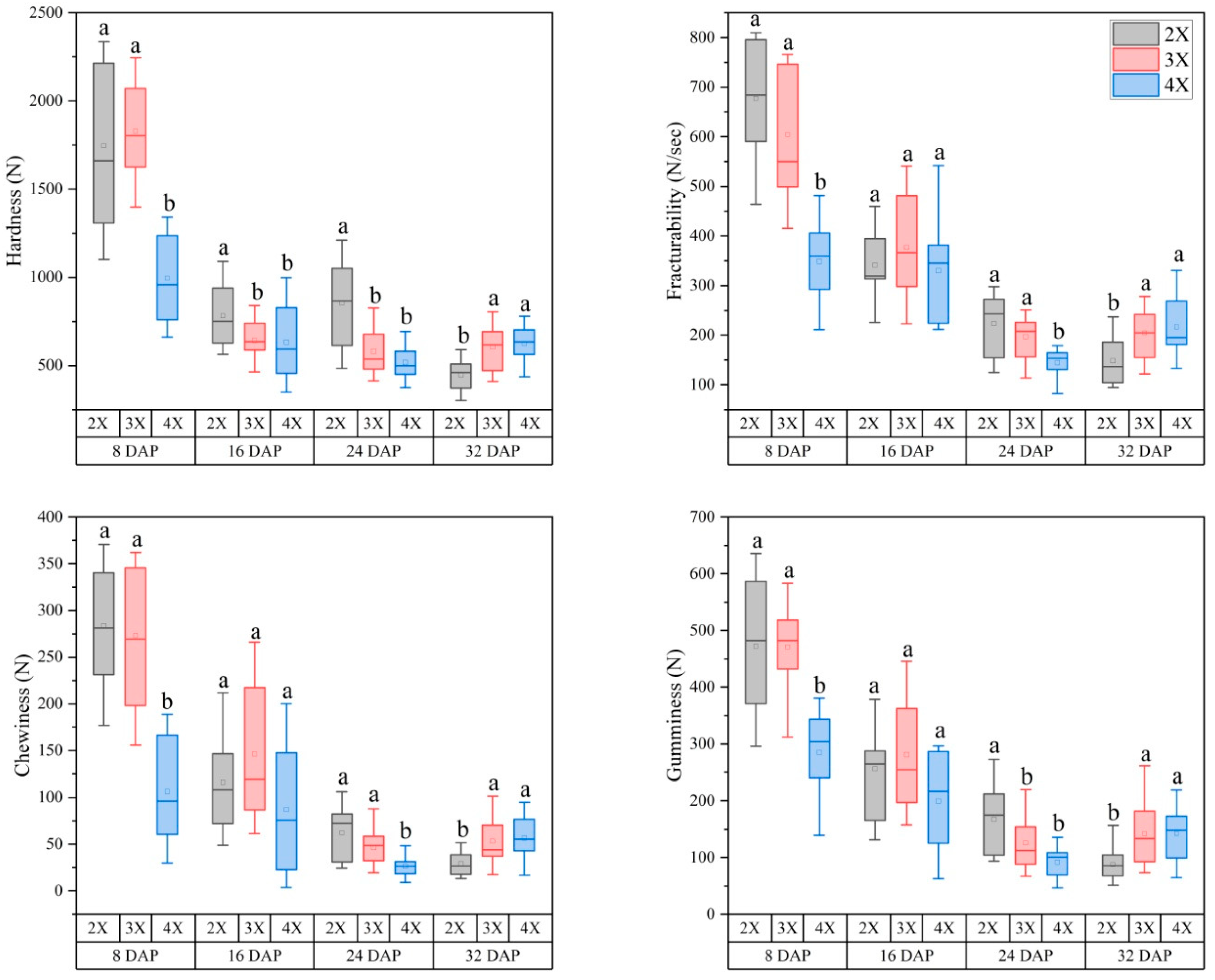
3.2. Variations in the Flesh Texture of Watermelon Fruits with Different Ploidy Levels
3.3. Variations in the Polysaccharide Content within the Cell Walls of Watermelon Flesh across Different Ploidy Levels
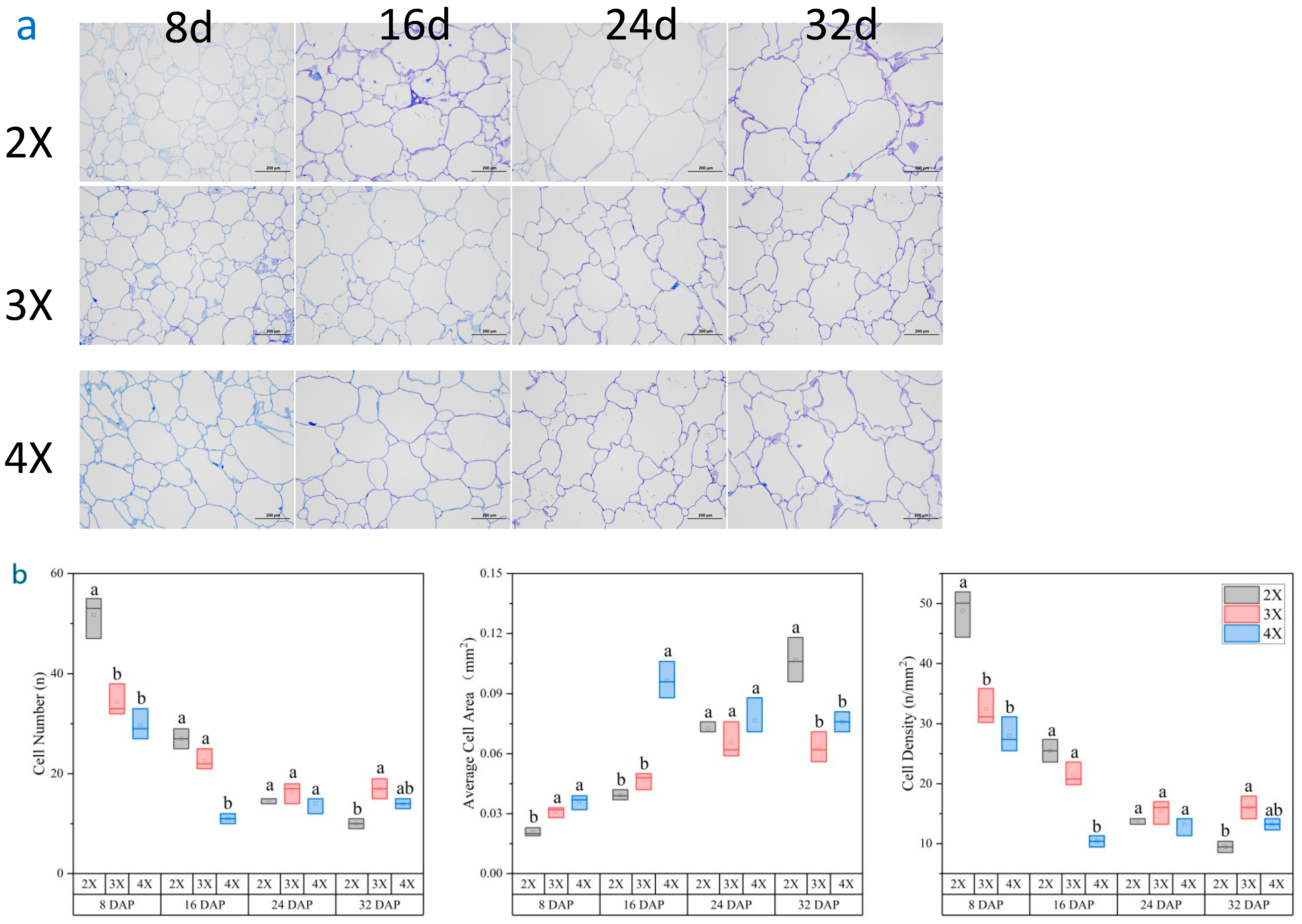
3.4. Differences in the Cellular Structure of Watermelon Flesh of Different Ploidy Levels
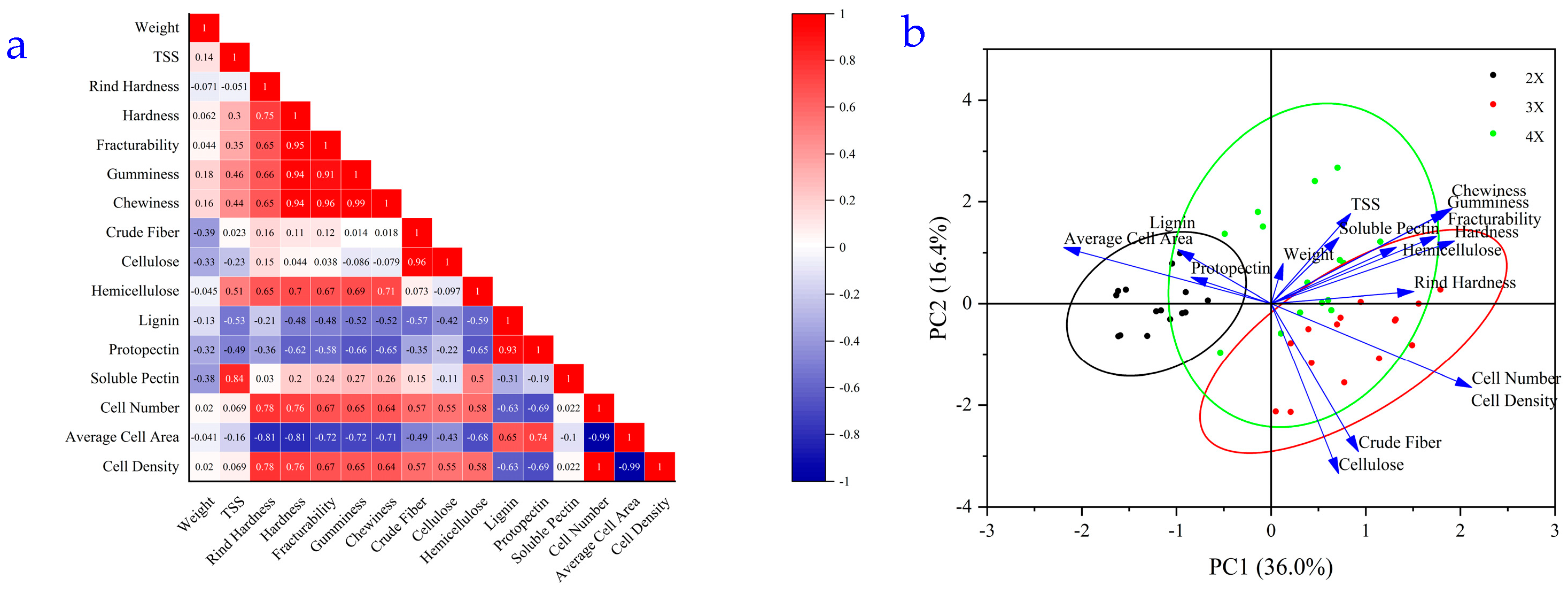
3.5. Correlation Analysis of Flesh Texture with Cell Microstructure and Cell Wall Polysaccharide Content in Watermelon at Different Ploidy Levels
4. Discussion
4.1. Differences in Fruit Weight and TSS Content among Different Ploidy Levels of Watermelon
4.2. Differences in Texture among Different Ploidy Levels of Watermelon
4.3. Differences in the Content of Cell Wall Polysaccharides in the Fruit Flesh among Different Ploidy Levels of Watermelon
4.4. Differences in Cell Structure among Different Ploidy Levels of Watermelon
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Guo, S.; Zhao, S.; Sun, H.; Wang, X.; Wu, S.; Lin, T.; Ren, Y.; Gao, L.; Deng, Y.; Zhang, J.; et al. Resequencing of 414 cultivated and wild watermelon accessions identifies selection for fruit quality traits. Nat. Genet. 2019, 51, 1616–1623. [Google Scholar] [CrossRef] [PubMed]
- FAO. Available online: https://faostat.fao.org/ (accessed on 12 June 2023).
- Bharadwaj, D.N. Polyploidy in Crop Improvement and Evolution. In Plant Biology and Biotechnology; Bahadur, B., Venkat Rajam, M., Sahijram, L., Krishnamurthy, K.V., Eds.; Springer: New Delhi, India, 2015; pp. 619–638. [Google Scholar]
- Soltis, P.S.; Marchant, D.B.; Van de Peer, Y.; Soltis, D.E. Polyploidy and genome evolution in plants. Curr. Opin. Plant Biol. 2015, 35, 119–125. [Google Scholar] [CrossRef]
- Madlung, A. Polyploidy and its effect on evolutionary success: Old questions revisited with new tools. Heredity 2013, 110, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Sattler, M.C.; Carvalho, C.R.; Clarindo, W.R. The polyploidy and its key role in plant breeding. Planta 2015, 243, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.; Fait, A.; Tel-Zur, N. Morphological, cytological and metabolic consequences of autopolyploidization in Hylocereus (Cactaceae) species. BMC Plant Biol. 2013, 13, 173. [Google Scholar] [CrossRef] [PubMed]
- Saminathan, T.; Nimmakayala, P.; Manohar, S.; Malkaram, S.; Almeida, A.; Cantrell, R.; Tomason, Y.; Abburi, L.; Rahman, M.A.; Vajja, V.G.; et al. Differential gene expression and alternative splicing between diploid and tetraploid watermelon. J. Exp. Bot. 2015, 66, 1369–1385. [Google Scholar] [CrossRef]
- Zhang, N.; Bao, Y.N.; Xie, Z.L.; Huang, X.; Sun, Y.H.; Feng, G.; Zeng, H.X.; Ren, J.; Li, Y.H.; Xiong, J.S.; et al. Efficient Characterization of Tetraploid Watermelon. Plants 2019, 8, 419. [Google Scholar] [CrossRef]
- Contador, L.; Shinya, P.; Infante, R. Texture phenotyping in fresh fleshy fruit. Sci. Hortic. 2015, 193, 40–46. [Google Scholar] [CrossRef]
- Predieri, S.; Ragazzini, P.; Rondelli, R. Sensory evaluation and peach fruit quality. Acta Hortic. 2006, 713, 429–434. [Google Scholar] [CrossRef]
- Risse, L.A.; Brecht, J.K.; Sargent, S.A.; Locascio, S.J.; Crall, J.M.; Elmstrom, G.W.; Maynard, D.N. Storage Characteristics of Small Watermelon Cultivars. J. Am. Soc. Hortic. Sci. 1990, 115, 440–443. [Google Scholar] [CrossRef]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.C.; Gao, Y.H.; Zhang, L.J.; Zhou, Y.H. The plant cell wall: Biosynthesis, construction, and functions. J. Integr. Plant Biol. 2021, 63, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.K.T.; Schröder, R.; Sutherland, P.W.; Hallett, I.C.; Hall, M.I.; Prakash, R.; Smith, B.G.; Melton, L.D.; Johnston, J.W. Cell wall structures leading to cultivar differences in softening rates develop early during apple (Malus × domestica) fruit growth. BMC Plant Biol. 2013, 13, 183. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.A.; Wang, W.; Zhang, Q.; Jia, W.S. Cell Wall Integrity Signaling in Fruit Ripening. Int. J. Mol. Sci. 2023, 24, 4054. [Google Scholar] [CrossRef] [PubMed]
- Brummell, D.A. Cell wall metabolism during maturation, ripening and senescence of peach fruit. J. Exp. Bot. 2004, 55, 2029–2039. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.D.; Guo, M.R.; Yang, W.T.; Liu, Y.X.; Wang, Y.; Chen, G.G. The Role of Cell Wall Polysaccharides Disassembly and Enzyme Activity Changes in the Softening Process of Hami melon (L.). Foods 2022, 11, 841. [Google Scholar] [CrossRef] [PubMed]
- Soteriou, G.A.; Siomos, A.S.; Gerasopoulos, D.; Rouphael, Y.; Georgiadou, S.; Kyriacou, M.C. Biochemical and histological contributions to textural changes in watermelon fruit modulated by grafting. Food Chem. 2017, 237, 133–140. [Google Scholar] [CrossRef]
- Liao, N.; Hu, Z.; Li, Y.; Hao, J.; Chen, S.; Xue, Q.; Ma, Y.; Zhang, K.; Mahmoud, A.; Ali, A.; et al. Ethylene-responsive factor 4is associated with the desirable rind hardness trait conferring cracking resistance in fresh fruits of watermelon. Plant Biotechnol. J. 2019, 18, 1066–1077. [Google Scholar] [CrossRef]
- Gao, Y.; Guo, Y.; Su, Z.Y.; Yu, Y.; Zhu, Z.C.; Gao, P.; Wang, X.Z. Transcriptome analysis of genes related to fruit texture in watermelon. Sci. Hortic. 2020, 262, 109075. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, Y.; Cui, H.; Zhang, L.; Sha, T.; Wang, C.; Fan, C.; Luan, F.; Wang, X. Linkage Mapping and Comparative Transcriptome Analysis of Firmness in Watermelon (Citrullus lanatus). Front. Plant Sci. 2020, 11, 831. [Google Scholar] [CrossRef]
- Anees, M.; Gao, L.; Umer, M.J.; Yuan, P.L.; Zhu, H.J.; Lu, X.Q.; He, N.; Gong, C.S.; Kaseb, M.O.; Zhao, S.J.; et al. Identification of Key Gene Networks Associated with Cell Wall Components Leading to Flesh Firmness in Watermelon. Front. Plant Sci. 2021, 12, 630243. [Google Scholar] [CrossRef] [PubMed]
- Anees, M.; Gao, L.; Gong, C.; Umer, M.J.; Yuan, P.; Zhu, H.; Lu, X.; He, N.; Kaseb, M.O.; Yang, D.; et al. Aux/IAA gene Cla004102, is involved in synergistic regulation of various endogenous hormones, regulating flesh firmness in watermelon. Sci. Hortic. 2023, 310, 111719. [Google Scholar] [CrossRef]
- Poles, L.; Gentile, A.; Giuffrida, A.; Valentini, L.; Endrizzi, I.; Aprea, E.; Gasperi, F.; Distefano, G.; Artioli, G.; La Malfa, S.; et al. Role of fruit flesh cell morphology and MdPG1 allelotype in influencing juiciness and texture properties in apple. Postharvest Biol. Technol. 2020, 164, 111161. [Google Scholar] [CrossRef]
- Kim, M.S.; Duizer, L.M.; Grygorczyk, A. Application of a texture analyzer friction rig to evaluate complex texture attributes in apples. Postharvest Biol. Technol. 2022, 186, 111820. [Google Scholar] [CrossRef]
- Bianchi, T.; Guerrero, L.; Gratacós-Cubarsí, M.; Claret, A.; Argyris, J.; Garcia-Mas, J.; Hortós, M. Textural properties of different melon (Cucumis melo L.) fruit types: Sensory and physical-chemical evaluation. Sci. Hortic. 2016, 201, 46–56. [Google Scholar] [CrossRef]
- Zhang, W.; Cui, D.; Ying, Y. Nondestructive measurement of pear texture by acoustic vibration method. Postharvest Biol. Technol. 2014, 96, 99–105. [Google Scholar] [CrossRef]
- Ciacciulli, A.; Cirilli, M.; Chiozzotto, R.; Attanasio, G.; Da Silva Linge, C.; Pacheco, I.; Rossini, L.; Bassi, D. Linkage and association mapping for the slow softening (SwS) trait in peach (P. persica L. Batsch) fruit. Tree Genet. Genomes 2018, 14, 93. [Google Scholar] [CrossRef]
- Pieniazek, F.; Messina, V. Texture and color analysis of freeze-dried potato (cv. Spunta) using instrumental and image analysis techniques. Int. J. Food Prop. 2016, 20, 1422–1431. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, S.; Lu, X.; He, N.; Gao, L.; Dou, J.; Bie, Z.; Liu, W. Genome duplication improves the resistance of watermelon root to salt stress. Plant Physiol. Biochem. 2018, 133, 11–21. [Google Scholar] [CrossRef]
- Oyeyinka, B.O.; Afolayan, A.J. Comparative Evaluation of the Nutritive, Mineral, and Antinutritive Composition of Musa sinensis L. (Banana) and Musa paradisiaca L. (Plantain) Fruit Compartments. Plants 2019, 8, 598. [Google Scholar] [CrossRef]
- Davis, A.; Webber, C.; Wenge, L.; Perkins, P.; Levi, A.; King, S. Watermelon Quality Traits as Affected by Ploidy. HortScience 2013, 48, 1113–1118. [Google Scholar] [CrossRef]
- Henderson, W. Effect of Cultivar, Polyploidy and “Reciprocal” Hybridization on Characters Important in Breeding Triploid Seedless Watermelon Hybrids1. J. Am. Soc. Hortic. Sci. 1977, 102, 293–297. [Google Scholar] [CrossRef]
- Mohammadi, V.; Talebi, S.; Ahmadnasab, M.; Mollahassanzadeh, H. The effect of induced polyploidy on phytochemistry, cellular organelles and the expression of genes involved in thymol and carvacrol biosynthetic pathway in thyme (Thymus vulgaris). Front. Plant Sci. 2023, 14, 1228844. [Google Scholar] [CrossRef] [PubMed]
- Perkins-Veazie, P.; Collins, J.K.; Davis, A.R.; Roberts, W. Carotenoid content of 50 watermelon cultivars. J. Agric. Food Chem. 2006, 54, 2593–2597. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Sun, Y.; Chen, F.; Yu, L.; Mao, Q.; Wang, L.; Guo, X.; Liu, C. Analysis of microstructures and macrotextures for different apple cultivars based on parenchyma morphology. Microsc. Res. Tech. 2016, 79, 304–312. [Google Scholar] [CrossRef]
- Li, H.; Pidakala, P.; Billing, D.; Burdon, J. Kiwifruit firmness: Measurement by penetrometer and non-destructive devices. Postharvest Biol. Technol. 2016, 120, 127–137. [Google Scholar] [CrossRef]
- Ramsey, J.; Schemske, D.W. Pathways, Mechanisms, and Rates of Polyploid Formation in Flowering Plants. Annu. Rev. Ecol. Syst. 1998, 29, 467–501. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.; He, N.; Anees, M.; Yang, D.; Kong, W.; Zhang, J.; Yuan, L.; Luo, X.; Zhu, H.; Liu, W. A Comparison of Watermelon Flesh Texture across Different Ploidy Levels Using Histology and Cell Wall Measurements. Horticulturae 2024, 10, 112. https://doi.org/10.3390/horticulturae10020112
Lu X, He N, Anees M, Yang D, Kong W, Zhang J, Yuan L, Luo X, Zhu H, Liu W. A Comparison of Watermelon Flesh Texture across Different Ploidy Levels Using Histology and Cell Wall Measurements. Horticulturae. 2024; 10(2):112. https://doi.org/10.3390/horticulturae10020112
Chicago/Turabian StyleLu, Xuqiang, Nan He, Muhammad Anees, Dongdong Yang, Weikang Kong, Jiwen Zhang, Luming Yuan, Xiaowen Luo, Hongju Zhu, and Wenge Liu. 2024. "A Comparison of Watermelon Flesh Texture across Different Ploidy Levels Using Histology and Cell Wall Measurements" Horticulturae 10, no. 2: 112. https://doi.org/10.3390/horticulturae10020112
APA StyleLu, X., He, N., Anees, M., Yang, D., Kong, W., Zhang, J., Yuan, L., Luo, X., Zhu, H., & Liu, W. (2024). A Comparison of Watermelon Flesh Texture across Different Ploidy Levels Using Histology and Cell Wall Measurements. Horticulturae, 10(2), 112. https://doi.org/10.3390/horticulturae10020112