


Seaweed-Derived Bio-Stimulant (Kelpak®) Enhanced the Morphophysiological, Biochemical, and Nutritional Quality of Salt-Stressed Spinach (Spinacia oleracea L.)

 ,
,  and
and
Abstract
1. Introduction
2. Material and Methods
2.1. Greenhouse Cultivation
2.1.1. Growth Conditions
2.1.2. Preparation of Plant Material
2.1.3. Seaweed Extract Preparation
2.1.4. Seaweed Extract and Saline Treatment
2.2. Biomass Evaluation
2.3. Physiological Attributes
2.4. Oxidative Stress Markers
2.5. Antioxidative Enzymes
2.6. Metabolites and Antioxidant Activity
2.7. Nutritional Constituents
2.8. Statistical Analysis
3. Results
3.1. Biomass Evaluation
3.2. Physiological Attributes
3.3. Oxidative Stress Markers
3.4. Antioxidant Defence Mechanisms
3.5. Nutritional Evaluation
3.5.1. Minerals and Vitamin Constituents
3.5.2. Proximate Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jameel, J.; Anwar, T.; Siddiqi, E.H.; Alomrani, S.O. Alleviation of NaCl Stress in Tomato Varieties by Promoting Morpho-Physiological Attributes and Biochemical Characters. Sci. Hortic. 2024, 323, 112496. [Google Scholar] [CrossRef]
- Mndi, O.; Sogoni, A.; Jimoh, M.O.; Wilmot, C.M.; Rautenbach, F.; Laubscher, C.P. Interactive Effects of Salinity Stress and Irrigation Intervals on Plant Growth, Nutritional Value, and Phytochemical Content in Mesembryanthemum crystallinum L. Agriculture 2023, 13, 1026. [Google Scholar] [CrossRef]
- Amerian, M.; Palangi, A.; Gohari, G.; Ntatsi, G. Enhancing Salinity Tolerance in Cucumber through Selenium Biofortification and Grafting. BMC Plant Biol. 2024, 24, 24. [Google Scholar] [CrossRef] [PubMed]
- Sogoni, A.; Jimoh, M.O.; Barker, A.M.; Keyster, M.; Kambizi, L.; Laubscher, C.P. Salinity Modulates Morpho-Physiology, Biochemical and Antioxidant Defence System in Tetragonia decumbens Mill.: A Neglected Wild Leafy Vegetable in South Africa. Plant Physiol. Rep. 2024, 1–14. [Google Scholar] [CrossRef]
- Kanwal, R.; Maqsood, M.F.; Shahbaz, M.; Naz, N.; Zulfiqar, U.; Ali, M.F.; Jamil, M.; Khalid, F.; Ali, Q.; Sabir, M.A.; et al. Exogenous Ascorbic Acid as a Potent Regulator of Antioxidants, Osmo-Protectants, and Lipid Peroxidation in Pea under Salt Stress. BMC Plant Biol. 2024, 24, 247. [Google Scholar] [CrossRef] [PubMed]
- Rani, J.; Paul, B. Challenges in Arid Region Reclamation with Special Reference to Indian Thar Desert—Its Conservation and Remediation Techniques. Int. J. Environ. Sci. Technol. 2023, 20, 12753–12774. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhowmik, P.C.; Hossain, M.A.; Rahman, M.M.; Prasad, M.N.V.; Ozturk, M.; Fujita, M. Potential Use of Halophytes to Remediate Saline Soils. Biomed Res. Int. 2014, 2014, 589341. [Google Scholar] [CrossRef]
- Qadir, M.; Oster, J.D.; Schubert, S.; Noble, A.D.; Sahrawat, K.L. Phytoremediation of Sodic and Saline-Sodic Soils. Adv. Agron. 2007, 96, 197–247. [Google Scholar]
- Mughunth, R.J.; Velmurugan, S.; Mohanalakshmi, M.; Vanitha, K. A Review of Seaweed Extract’s Potential as a Biostimulant to Enhance Growth and Mitigate Stress in Horticulture Crops. Sci. Hortic. 2024, 334, 113312. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production. Plants 2021, 10, 531. [Google Scholar] [CrossRef] [PubMed]
- Deolu-Ajayi, A.O.; van der Meer, I.M.; van der Werf, A.; Karlova, R. The Power of Seaweeds as Plant Biostimulants to Boost Crop Production under Abiotic Stress. Plant Cell Environ. 2022, 45, 2537–2553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yin, J.; Ma, Y.; Peng, Y.; Fenton, O.; Wang, W.; Zhang, W.; Chen, Q. Unlocking the Potential of Biostimulants Derived from Organic Waste and By-Product Sources: Improving Plant Growth and Tolerance to Abiotic Stresses in Agriculture. Environ. Technol. Innov. 2024, 34, 103571. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Sánchez-Hernández, C.V.; Palmeros-Suárez, P.A.; Ocampo-Alvarez, H.; Santacruz-Ruvalcaba, F.; Meza-Canales, I.D.; Becerril-Espinosa, A. Seaweed Extract Improves Growth and Productivity of Tomato Plants under Salinity Stress. Agronomy 2022, 12, 2495. [Google Scholar] [CrossRef]
- Pal, S.C.; Hossain, M.B.; Mallick, D.; Bushra, F.; Abdullah, S.R.; Dash, P.K.; Das, D. Combined Use of Seaweed Extract and Arbuscular Mycorrhizal Fungi for Alleviating Salt Stress in Bell Pepper (Capsicum annuum L.). Sci. Hortic. 2024, 325, 112597. [Google Scholar] [CrossRef]
- Gul, S.; Nawaz, M.F.; Bin Yousaf, M.T.; Haroon U Rashid, M.; Adnan, M.Y.; Tausif, S.; Javed, A.; Abideen, Z.; El Keblawy, A. Brown Macro-Seaweeds Derived Agro-Biostimulant for Zea mays Farming in Saline Conditions: Growth Enhancement and Optimum Biochemical and Ion Feedback. Biocatal. Agric. Biotechnol. 2024, 57, 103105. [Google Scholar] [CrossRef]
- Salehi, B.; Tumer, T.B.; Ozleyen, A.; Peron, G.; Dall’Acqua, S.; Rajkovic, J.; Naz, R.; Nosheen, A.; Mudau, F.N.; Labanca, F.; et al. Plants of the Genus Spinacia: From Bioactive Molecules to Food and Phytopharmacological Applications. Trends Food Sci. Technol. 2019, 88, 260–273. [Google Scholar] [CrossRef]
- Shokri, N.; Hassani, A.; Sahimi, M. Multi-Scale Soil Salinization Dynamics from Global to Pore Scale: A Review. Rev. Geophys. 2024, 62, e2023RG000804. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Cozzolino, E.; Ottaiano, L.; Petropoulos, S.A.; Nocerino, S.; Pelosi, M.E.; Rouphael, Y.; Mori, M.; Di Mola, I. Effect of Biostimulant Application on Plant Growth, Chlorophylls and Hydrophilic Antioxidant Activity of Spinach (Spinacia oleracea L.) Grown under Saline Stress. Horticulturae 2022, 8, 971. [Google Scholar] [CrossRef]
- Yavuz, D.; Kılıç, E.; Seymen, M.; Dal, Y.; Kayak, N.; Kal, Ü.; Yavuz, N. The Effect of Irrigation Water Salinity on the Morph-Physiological and Biochemical Properties of Spinach under Deficit Irrigation Conditions. Sci. Hortic. 2022, 304, 111272. [Google Scholar] [CrossRef]
- Aremu, A.O.; Plačková, L.; Gruz, J.; Bíba, O.; Novák, O.; Stirk, W.A.; Doležal, K.; Van Staden, J. Seaweed-Derived Biostimulant (Kelpak®) Influences Endogenous Cytokinins and Bioactive Compounds in Hydroponically Grown Eucomis autumnalis. J. Plant Growth Regul. 2016, 35, 151–162. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant Responses to Drought, Salinity and Extreme Temperatures: Towards Genetic Engineering for Stress Tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Bistgani, Z.E.; Hashemi, M.; DaCosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of Salinity Stress on the Physiological Characteristics, Phenolic Compounds and Antioxidant Activity of Thymus vulgaris L. and Thymus Daenensis Celak. Ind. Crops Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Gokul, A.; Fahiem Carelse, M.; Niekerk, L.A.; Klein, A.; Ludidi, N.; Mendoza-Cozatl, D.; Keyster, M. Exogenous 3,3′-Diindolylmethane Improves Vanadium Stress Tolerance in Brassica napus Seedling Shoots by Modulating Antioxidant Enzyme Activities. Biomolecules 2021, 11, 436. [Google Scholar] [CrossRef]
- González-Orenga, S.; Leandro, M.E.D.A.; Tortajada, L.; Grigore, M.N.; Llorens, J.A.; Ferrer-Gallego, P.P.; Laguna, E.; Boscaiu, M.; Vicente, O. Comparative Studies on the Stress Responses of Two Bupleurum (Apiaceae) Species in Support of Conservation Programmes. Environ. Exp. Bot. 2021, 191, 104616. [Google Scholar] [CrossRef]
- Ali, A.E.E.; Ludidi, N. Antioxidant Responses Are Associated with Differences in Drought Tolerance between Maize and Sorghum. J. Oasis Agric. Sustain. Dev. 2021, 3, 1–12. [Google Scholar] [CrossRef]
- Kaur, N.; Dhawan, M.; Sharma, I.; Pati, P.K. Interdependency of Reactive Oxygen Species Generating and Scavenging System in Salt Sensitive and Salt Tolerant Cultivars of Rice. BMC Plant Biol. 2016, 16, 131. [Google Scholar] [CrossRef]
- Din Muhammad, H.M.; Anjum, M.A.; Naz, S. Silicon-Mediated Alleviation of Salinity Stress in Petunia (Petunia hybrida) by Modulation of Morphological, Physiological and Biochemical Indices. J. Soil Sci. Plant Nutr. 2024, 24, 2221–2231. [Google Scholar] [CrossRef]
- Omran, R.G. Peroxide Levels and the Activities of Catalase, Peroxidase, and Indoleacetic Acid Oxidase during and after Chilling Cucumber Seedlings. Plant Physiol. 1980, 65, 407–408. [Google Scholar] [CrossRef]
- Jasson, T.I.; Jimoh, M.O.; Daniels, C.W.; Nchu, F.; Laubscher, C.P. Enhancement of Antioxidant Potential, Phytochemicals, Nutritional Properties, and Growth of Siphonochilus aethiopicus (Schweinf.) B.L.Burtt with Different Dosages of Compost Tea. Horticulturae 2023, 9, 274. [Google Scholar] [CrossRef]
- Zantanta, N.; Kambizi, L.; Etsassala, N.G.E.R.; Nchu, F. Comparing Crop Yield, Secondary Metabolite Contents, and Antifungal Activity of Extracts of Helichrysum odoratissimum Cultivated in Aquaponic, Hydroponic, and Field Systems. Plants 2022, 11, 2696. [Google Scholar] [CrossRef] [PubMed]
- Ngxabi, S.; Jimoh, M.O.; Kambizi, L.; Laubscher, C.P. Growth Characteristics, Phytochemical Contents, and Antioxidant Capacity of Trachyandra ciliata (L.f) Kunth Grown in Hydroponics under Varying Degrees of Salinity. Horticulturae 2021, 7, 244. [Google Scholar] [CrossRef]
- Bulawa, B.; Sogoni, A.; Jimoh, M.O.; Laubscher, C.P. Potassium Application Enhanced Plant Growth, Mineral Composition, Proximate and Phytochemical Content in Trachyandra divaricata Kunth (Sandkool). Plants 2022, 11, 3183. [Google Scholar] [CrossRef]
- Tshayingwe, A.; Jimoh, M.O.; Sogoni, A.; Wilmot, C.M.; Laubscher, C.P. Light Intensity and Growth Media Influence Growth, Nutrition, and Phytochemical Content in Trachyandra divaricata Kunth. Agronomy 2023, 13, 247. [Google Scholar] [CrossRef]
- Nemzer, B.; Al-Taher, F.; Abshiru, N. Extraction and Natural Bioactive Molecules Characterization in Spinach, Kale and Purslane: A Comparative Study. Molecules 2021, 26, 2515. [Google Scholar] [CrossRef]
- Verma, O.; Sharma, S.; Kumar, V.; Singh, T.; Kumar, R.; Auji, R. Salinity Stress Effect on Staple Food Crops and Novel Mitigation Strategies. Biologia 2024, 79, 2359–2374. [Google Scholar] [CrossRef]
- Azeem, M.; Pirjan, K.; Qasim, M.; Mahmood, A.; Javed, T.; Muhammad, H.; Yang, S.; Dong, R.; Ali, B.; Rahimi, M. Salinity Stress Improves Antioxidant Potential by Modulating Physio-Biochemical Responses in Moringa oleifera Lam. Sci. Rep. 2023, 13, 2895. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, M.E.-S.; Shalaby, O.A. Effect of magnetized water and seaweed extract on growth and yield of squash (Cucurbita pepo L.) plants grown under saline conditions. Egypt. J. Desert Res. 2024, 74, 197–215. [Google Scholar] [CrossRef]
- Radwan, A.M.; Ahmed, E.A.; Donia, A.M.; Mustafa, A.E.; Balah, M.A. Priming of Citrullus lanatus var. colocynthoides Seeds in Seaweed Extract Improved Seed Germination, Plant Growth and Performance under Salinity Conditions. Sci. Rep. 2023, 13, 11884. [Google Scholar] [CrossRef]
- Rouphael, Y.; De Micco, V.; Arena, C.; Raimondi, G.; Colla, G.; De Pascale, S. Effect of Ecklonia Maxima Seaweed Extract on Yield, Mineral Composition, Gas Exchange, and Leaf Anatomy of Zucchini Squash Grown under Saline Conditions. J. Appl. Phycol. 2017, 29, 459–470. [Google Scholar] [CrossRef]
- Wilmot, C.M.; Jimoh, M.; Laubscher, C. Stimulatory Effects of an Exogenously Applied Seaweed Extract on the Morphological and Physiological Growth and Yield in Juvenile Amaryllis Belladonna L. Bulbs. Egypt. J. Bot. 2024, 64, 52–68. [Google Scholar] [CrossRef]
- Boussora, F.; Triki, T.; Bennani, L.; Bagues, M.; Ben Ali, S.; Ferchichi, A.; Ngaz, K.; Guasmi, F. Mineral Accumulation, Relative Water Content and Gas Exchange Are the Main Physiological Regulating Mechanisms to Cope with Salt Stress in Barley. Sci. Rep. 2024, 14, 14931. [Google Scholar] [CrossRef] [PubMed]
- Banakar, M.H.; Amiri, H.; Sarafraz Ardakani, M.R.; Ranjbar, G.H. Susceptibility and Tolerance of Fenugreek (Trigonella Foenum-Graceum L.) to Salt Stress: Physiological and Biochemical Inspections. Environ. Exp. Bot. 2022, 194, 104748. [Google Scholar] [CrossRef]
- Khatri, K.; Rathore, M.S. Salt and Osmotic Stress-Induced Changes in Physio-Chemical Responses, PSII Photochemistry and Chlorophyll a Fluorescence in Peanut. Plant Stress 2022, 3, 100063. [Google Scholar] [CrossRef]
- Krid, A.; El Hallabi, M.; Ennoury, A.; Nhhala, N.; Aberkani, K.; Nhiri, M.; Zerrouk, M.H. The Potential of Seaweed Extracts as a Biostimulant for Improving Salt Stress Tolerance of Solanum Lycopersicum L. S. Afr. J. Bot. 2023, 161, 305–316. [Google Scholar] [CrossRef]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to Drought and Salt Stress in Plants: Unraveling the Signaling Networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed]
- González-Orenga, S.; Grigore, M.-N.; Boscaiu, M.; Vicente, O. Constitutive and Induced Salt Tolerance Mechanisms and Potential Uses of Limonium Mill. Species. Agronomy 2021, 11, 413. [Google Scholar] [CrossRef]
- Rodrigues de Queiroz, A.; Hines, C.; Brown, J.; Sahay, S.; Vijayan, J.; Stone, J.M.; Bickford, N.; Wuellner, M.; Glowacka, K.; Buan, N.R.; et al. The Effects of Exogenously Applied Antioxidants on Plant Growth and Resilience. Phytochem. Rev. 2023, 22, 407–447. [Google Scholar] [CrossRef]
- Ozgur, R.; Uzilday, B.; Sekmen, A.H.; Turkan, I.; Ozgur, R.; Uzilday, B.; Sekmen, A.H.; Turkan, I. Reactive Oxygen Species Regulation and Antioxidant Defence in Halophytes. Funct. Plant Biol. 2013, 40, 832–847. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Plant Salt Tolerance: Adaptations in Halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.; Zulfiqar, A.; Ali, B.; Naseeb, M.A.; Almasaudi, A.S.; Harakeh, S. Iron Sulfate (FeSO4) Improved Physiological Attributes and Antioxidant Capacity by Reducing Oxidative Stress of Oryza Sativa L. Cultivars in Alkaline Soil. Sustainability 2022, 14, 16845. [Google Scholar] [CrossRef]
- Shahid, M.; Singh, U.B.; Farah, M.A.; Al-Anazi, K.M. Phyto-Toxicological Effect of Fipronil to Plant Seedlings: Assessing Germination Attributes, Root-Tip Morphology, Oxidative Stress, and Cellular Respiration Indices. Pestic Biochem. Physiol. 2024, 205, 106135. [Google Scholar] [CrossRef]
- Saddique, M.; Kausar, A.; Iqra, I.; Akhter, N.; Mujahid, N.; Parveen, A.; Zaman, Q.; Hussain, S. Amino Acids Application Alleviated Salinity Stress in Spinach (Spinacia Oleracea L.) by Improving Oxidative Defense, Osmolyte Accumulation, and Nutrient Balance. Turk. J. Agric. For. 2022, 46, 875–887. [Google Scholar] [CrossRef]
- Ors, S.; Suarez, D.L. Spinach Biomass Yield and Physiological Response to Interactive Salinity and Water Stress. Agric. Water Manag. 2017, 190, 31–41. [Google Scholar] [CrossRef]
- Naz, T.; Mazhar Iqbal, M.; Tahir, M.; Hassan, M.M.; Rehmani, M.I.A.; Zafar, M.I.; Ghafoor, U.; Qazi, M.A.; El Sabagh, A.; Sakran, M.I. Foliar Application of Potassium Mitigates Salinity Stress Conditions in Spinach (Spinacia Oleracea L.) through Reducing NaCl Toxicity and Enhancing the Activity of Antioxidant Enzymes. Horticulturae 2021, 7, 566. [Google Scholar] [CrossRef]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hasanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide Dismutase—Mentor of Abiotic Stress Tolerance in Crop Plants. Environ. Sci. Pollut. Res. 2015, 22, 10375–10394. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Ashraf, M. Antioxidants as Modulators of Arsenic-Induced Oxidative Stress Tolerance in Plants: An Overview. J. Hazard. Mater. 2022, 427, 127891. [Google Scholar] [CrossRef]
- Umair Hassan, M.; Chattha, M.U.; Khan, I.; Khan, T.A.; Nawaz, M.; Tang, H.; Noor, M.A.; Asseri, T.A.Y.; Hashem, M.; Guoqin, H. Zinc Seed Priming Alleviates Salinity Stress and Enhances Sorghum Growth by Regulating Antioxidant Activities, Nutrient Homeostasis, and Osmolyte Synthesis. Agronomy 2024, 14, 1815. [Google Scholar] [CrossRef]
- Yarşı, G. Effects of Mycorrhiza, Seaweed and Bionutrient Applied to Reduce the Salt Stress on Nutrient Content, Plant Growth, Malondialdehyde (MDA) and Proline in Pepper. J. Elem. 2023, 28, 533–545. [Google Scholar] [CrossRef]
- Alsherif, E.A.; Yaghoubi Khanghahi, M.; Crecchio, C.; Korany, S.M.; Sobrinho, R.L.; AbdElgawad, H. Understanding the Active Mechanisms of Plant (Sesuvium portulacastrum L.) against Heavy Metal Toxicity. Plants 2023, 12, 676. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.R.; Azhar, W.; Fan, X.; Ulhassan, Z.; Salam, A.; Ashraf, M.; Liu, Y.; Gan, Y. Efficacy of Zinc-Based Nanoparticles in Alleviating the Abiotic Stress in Plants: Current Knowledge and Future Perspectives. Environ. Sci. Pollut. Res. 2023, 30, 110047–110068. [Google Scholar] [CrossRef]
- Nabati, J.; Kafi, M.; Nezami, A.; Rezvani Moghaddam, P.; Masoumi, A.; Zare Mehrjerdi, M. Evaluation of Quantitative and Qualitative Characteristic of Forage Kochia (Kochia scoparia) in Different Salinity Levels and Time. Iran. J. Field Crops Res. 2014, 12, 613–620. [Google Scholar] [CrossRef]
- Hedayati-Firoozabadi, A.; Kazemeini, S.A.; Pirasteh-Anosheh, H.; Ghadiri, H.; Pessarakli, M. Forage Yield and Quality as Affected by Salt Stress in Different Ratios of Sorghum bicolor-Bassia indica Intercropping. J. Plant Nutr. 2020, 43, 2579–2589. [Google Scholar] [CrossRef]
- Athar, H.U.R.; Zulfiqar, F.; Moosa, A.; Ashraf, M.; Zafar, Z.U.; Zhang, L.; Ahmed, N.; Kalaji, H.M.; Nafees, M.; Hossain, M.A.; et al. Salt Stress Proteins in Plants: An Overview. Front. Plant Sci. 2022, 13, 999058. [Google Scholar] [CrossRef] [PubMed]
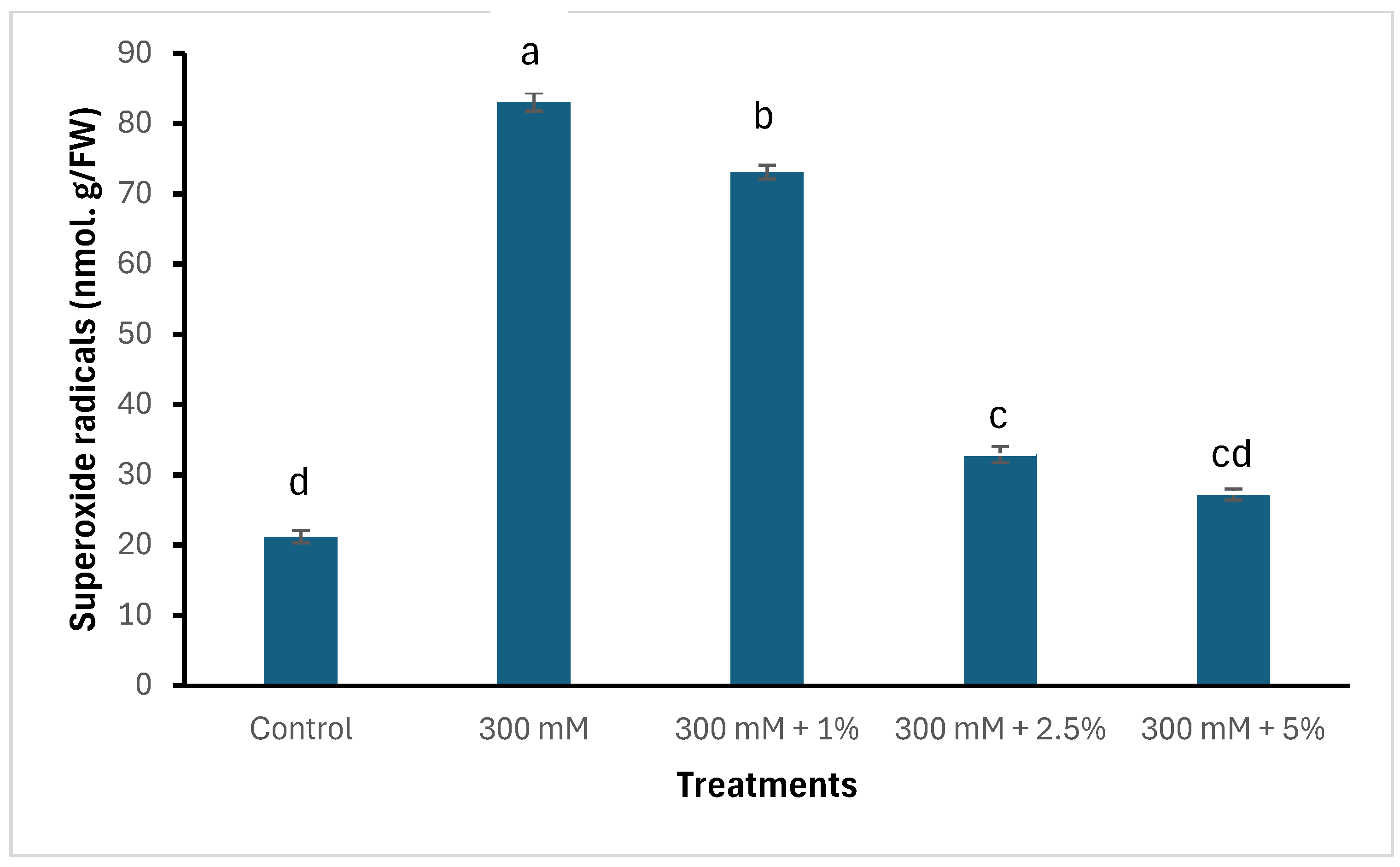


{kind=link}
{kind=link}
| Treatments | SFW (g) | SDW (g) | RFW (g) | RDW (g) | TFW (g) | TDW (g) |
|---|---|---|---|---|---|---|
| Control | 237 ± 1.7 a | 22.2 ± 0.3 a | 22.4 ± 1.1 a | 9.7 ± 0.7 a | 259.2 ± 1.8 a | 32.1 ± 0.6 a |
| 300 mM NaCl | 92.5 ± 1.1 d | 8.7 ± 1.6 d | 10 ± 0.8 c | 1.9 ± 0.9 e | 101.2 ± 1.6 d | 11.9 ± 1 e |
| 300 mM + 1% SWE | 144.9 ± 1.6 c | 12.2 ± 0.4 c | 17.7 ± 0.3 b | 3.5 ± 0.4 d | 157.1 ± 0.8 c | 21.2 ± 0.7 d |
| 300 mM + 2.5% SWE | 191.4 ± 2.1 b | 16.1 ± 0.9 b | 21.7 ± 1.2 a | 4.4 ± 0.1 c | 207.5 ± 1.2 b | 26.1 ± 0.9 c |
| 300 mM + 5% SWE | 214.3 ± 1.1 a | 21.3 ± 1.5 a | 21.5 ± 0.9 a | 7.8 ± 0.7 b | 235.6 ± 1.1 a | 29.4 ± 0.2 b |
| Treatments | Chl a (mg/g FW) | Chl b (mg/g FW) | Chl a + b (mg/g FW) | RWC % |
|---|---|---|---|---|
| Control | 6.7 ± 0.5 a | 4.1 ± 0.2 a | 10.8 ± 0.3 a | 90.5 ± 1.3 a |
| 300 mM NaCl | 2.7 ± 0.1 c | 0.8 ± 0.6 d | 3.5 ± 0.8 d | 65 ± 1 d |
| 300 mM + 1% SWE | 4.7 ± 0.3 b | 2.1 ± 0.5 c | 6.8 ± 0.6 c | 68 ± 1.4 c |
| 300 mM + 2.5% SWE | 5 ± 0.2 b | 3.1 ± 0.9 b | 8.1 ± 0.3 b | 79.9 ± 1 b |
| 300 mM + 5% SWE | 6.5 ± 0.4 a | 3.9 ± 0.3 a | 10.4 ± 0.3 a | 88.8 ± 1.2 a |
| Treatments | SOD Activity (U/g−1 Protein) | CAT Activity (U/g−1 Protein) | POD Activity (U/ g−1 Protein) | Polyphenols (mg GAE/g) | Flavonols (mg QE/g) | DPPH (µmol TE/g) |
|---|---|---|---|---|---|---|
| Control | 25.9 ± 0.3 d | 9.3 ± 0.4 d | 11.8 ± 1.6 d | 5.1 ± 0.1 c | 0.7 ± 0.1 c | 27.3 ± 1.3 d |
| 300 mM NaCl | 70.6 ± 0.4 c | 22.6 ± 0.3 a | 24.7 ± 1.8 a | 7.4 ± 0.7 a | 1.7 ± 0.02 a | 51.5 ± 1 a |
| 300 mM + 1% SWE | 70.5 ± 1.3 c | 19 ± 0.5 b | 18.4 ± 1.4 b | 6.6 ± 0.5 b | 1.1 ± 0.01 b | 39.6 ± 1.4 b |
| 300 mM + 2.5% SWE | 73.6 ± 0.8 b | 12.1 ± 0.1 c | 15.1 ± 0.8 c | 6.4 ± 0.2 b | 1 ± 0.01 bc | 31.9 ± 1.9 c |
| 300 mM + 5% SWE | 79.9 ± 0.5 a | 10.2 ± 0.7 cd | 12.6 ± 1.3 d | 5.1 ± 0.9 c | 0.9 ± 0.04 bc | 28.5 ± 1.1 d |
| Treatments | N (mg/100g) | P (mg/100g) | K (mg/100g) | Mg (mg/100g) | Na (mg/100g) | Ca (mg/100g) |
|---|---|---|---|---|---|---|
| Control | 4204 ± 4 a | 612 ± 12.5 b | 6837 ± 16 a | 773 ± 12 a | 3625 ± 4.2 d | 528 ± 11 a |
| 300 mM NaCl | 3107 ± 7 e | 407 ± 9.3 d | 4122 ± 25 e | 507 ± 11 d | 7812 ± 12.5 a | 242 ± 9 d |
| 300 mM + 1% SWE | 3521 ± 11.3 d | 505 ± 5 c | 5065 ± 5 d | 702 ± 14 c | 4662 ± 2.5 b | 342 ± 14 c |
| 300 mM + 2.5% SWE | 3651 ± 6.5 c | 522 ± 18 c | 6554 ± 4 c | 722 ± 15 b | 3826 ± 13 c | 407 ± 16 b |
| 300 mM + 5% SWE | 3728 ± 19.1 b | 662 ± 14 a | 6645 ± 15 b | 767 ± 11 a | 3629 ± 16 d | 514 ± 12 a |
| Treatments | Zn (mg/100g) | Mn (mg/100g) | Cu (mg/100g) | Fe (mg/100g) | Vitamin C (mg/100g) | Vitamin E (µg/100g) |
|---|---|---|---|---|---|---|
| Control | 8.6 ± 0.1 b | 11.2 ± 0.4 e | 0.3 ± 0.02 a | 44.2 ± 0.1 e | 51.3 ± 0.5 a | 15.3 ± 0.08 a |
| 300 mM NaCl | 5.5 ± 0.05 d | 12 ± 0.02 d | 0 ± 0.0 d | 72.7 ± 0.2 d | 20.7 ± 0.2 d | 4.3 ± 0.1 d |
| 300 mM + 1% SWE | 7.6 ± 0.01 c | 16.6 ± 0.5 c | 0 ± 0.0 d | 91.7 ± 0.8 c | 34.9 ± 0.4 c | 8.3 ± 0.9 c |
| 300 mM + 2.5% SWE | 8 ± 0.1 b | 18.6 ± 0.8 b | 0.1 ± 0.0 c | 121.1 ± 0.1 b | 40.6 ± 0.5 b | 10.2 ± 0.1 b |
| 300 mM + 5% SWE | 10.1 ± 0.3 a | 23.1 ± 1.9 a | 0.3 ± 0.0 b | 173 ± 0.8 a | 51.1 ± 1.3 a | 15.2 ± 1.1 a |
| Treatments | Ash% | Crude fat% | Protein% | Moisture% | NFC% | NDF% |
|---|---|---|---|---|---|---|
| Control | 33.9 ± 2.1 e | 1.5 ± 0.05 c | 26.4 ± 1.6 a | 7.1 ± 0.5 a | 32.1 ± 1.2 a | 27.8 ± 0.2 d |
| 300 mM NaCl | 54.5 ± 1.3 a | 1.1 ± 0.1 d | 18.1 ± 0.8 d | 4.1 ± 0.9 e | 16.8 ± 0.3 e | 42.2 ± 1 a |
| 300 mM + 1% SWE | 41.2 ± 1.1 b | 1.5 ± 0.3 c | 20.2 ± 0.4 c | 5.4 ± 0.2 d | 19.3 ± 0.9 d | 35.3 ± 0.4 b |
| 300 mM + 2.5% SWE | 38.5 ± 1.4 c | 2.2 ± 0.8 b | 21.8 ± 0.8 b | 5.9 ± 0.3 c | 22.4 ± 1 c | 33.8 ± 0.9 c |
| 300 mM + 5% SWE | 36.8 ± 1.5 d | 2.7 ± 0.05 a | 26 ± 0.3 a | 6.2 ± 0.8 b | 30 ± 1.1 b | 27.5 ± 1.1 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sogoni, A.; Ngcobo, B.L.; Jimoh, M.O.; Kambizi, L.; Laubscher, C.P. Seaweed-Derived Bio-Stimulant (Kelpak®) Enhanced the Morphophysiological, Biochemical, and Nutritional Quality of Salt-Stressed Spinach (Spinacia oleracea L.). Horticulturae 2024, 10, 1340. https://doi.org/10.3390/horticulturae10121340
Sogoni A, Ngcobo BL, Jimoh MO, Kambizi L, Laubscher CP. Seaweed-Derived Bio-Stimulant (Kelpak®) Enhanced the Morphophysiological, Biochemical, and Nutritional Quality of Salt-Stressed Spinach (Spinacia oleracea L.). Horticulturae. 2024; 10(12):1340. https://doi.org/10.3390/horticulturae10121340
Chicago/Turabian StyleSogoni, Avela, Bonga Lewis Ngcobo, Muhali Olaide Jimoh, Learnmore Kambizi, and Charles Petrus Laubscher. 2024. "Seaweed-Derived Bio-Stimulant (Kelpak®) Enhanced the Morphophysiological, Biochemical, and Nutritional Quality of Salt-Stressed Spinach (Spinacia oleracea L.)" Horticulturae 10, no. 12: 1340. https://doi.org/10.3390/horticulturae10121340
APA StyleSogoni, A., Ngcobo, B. L., Jimoh, M. O., Kambizi, L., & Laubscher, C. P. (2024). Seaweed-Derived Bio-Stimulant (Kelpak®) Enhanced the Morphophysiological, Biochemical, and Nutritional Quality of Salt-Stressed Spinach (Spinacia oleracea L.). Horticulturae, 10(12), 1340. https://doi.org/10.3390/horticulturae10121340