
Comparative Transcriptome Analysis of Non-Organogenic and Organogenic Tissues of Gaillardia pulchella Revealing Genes Regulating De Novo Shoot Organogenesis
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Culture Establishment
2.2. Total RNA Extraction and cDNA Library Construction
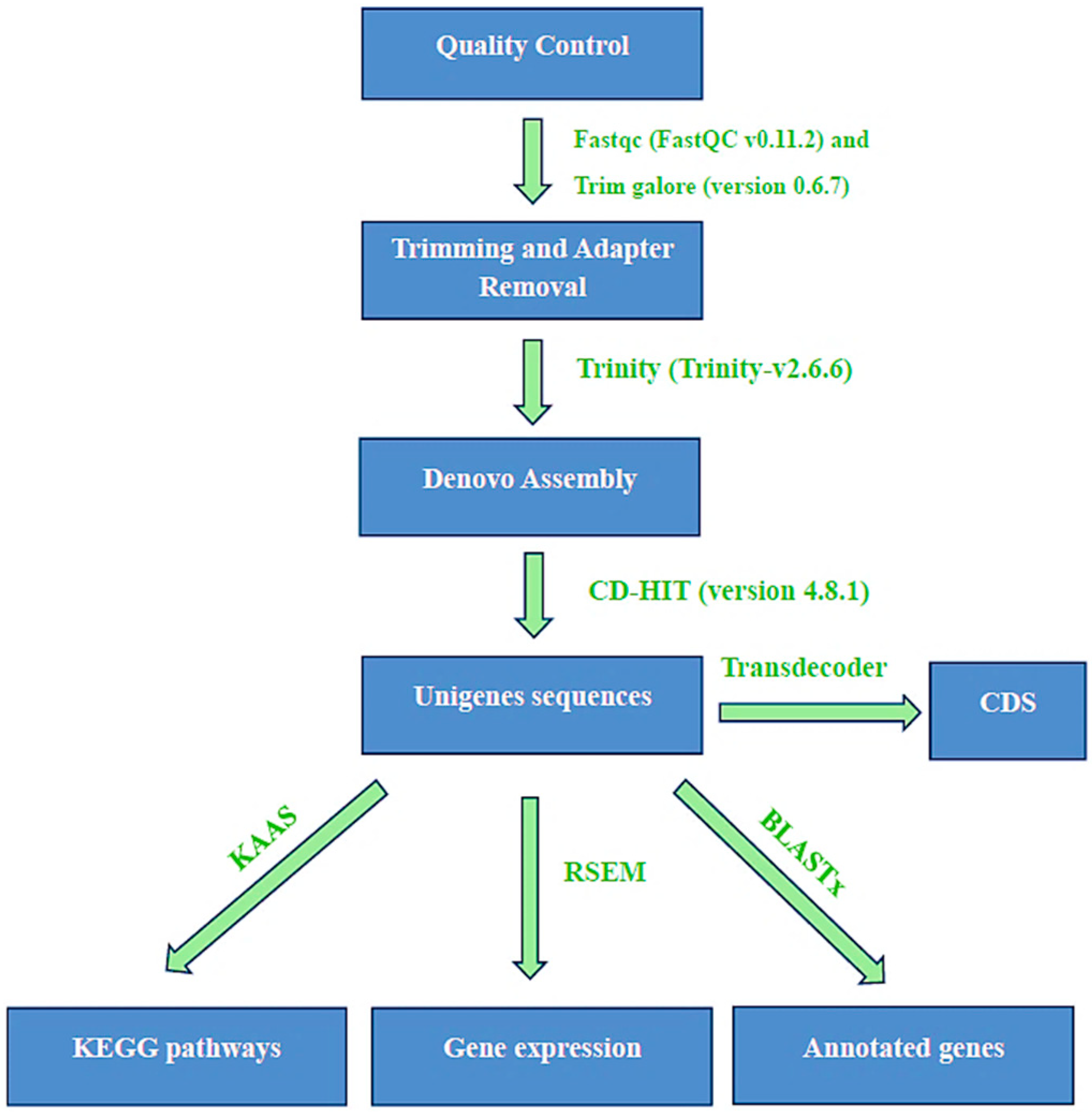
2.3. Transcriptome De Novo Assembly and Functional Annotation
2.4. Differential Gene Expression (DEG) Analysis
2.5. Statistical Analyses
3. Results
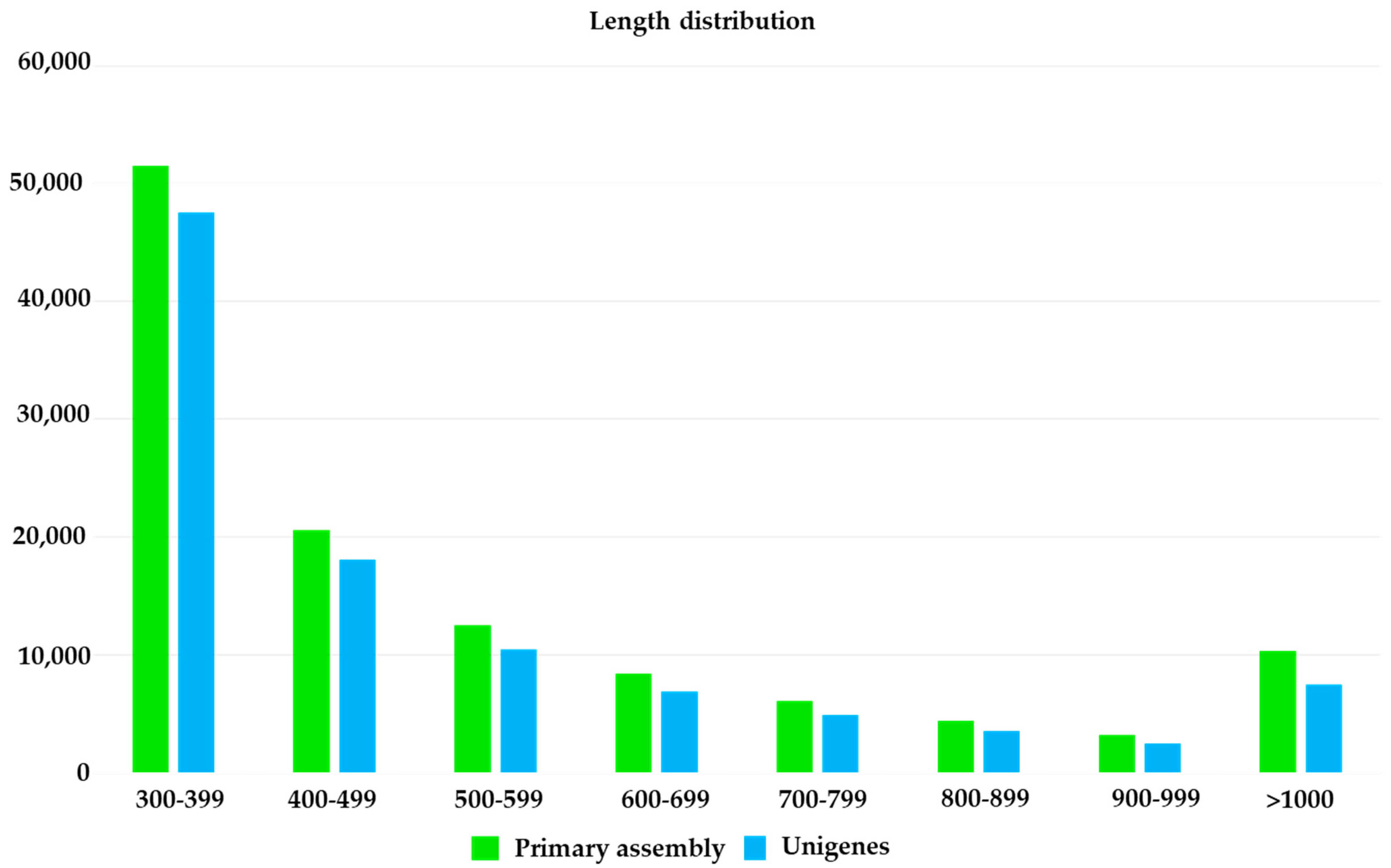
3.1. G. pulchella Transcriptomes and Some Unique Features
3.2. Coding Sequence (CDS) Prediction
3.3. Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathways Classification
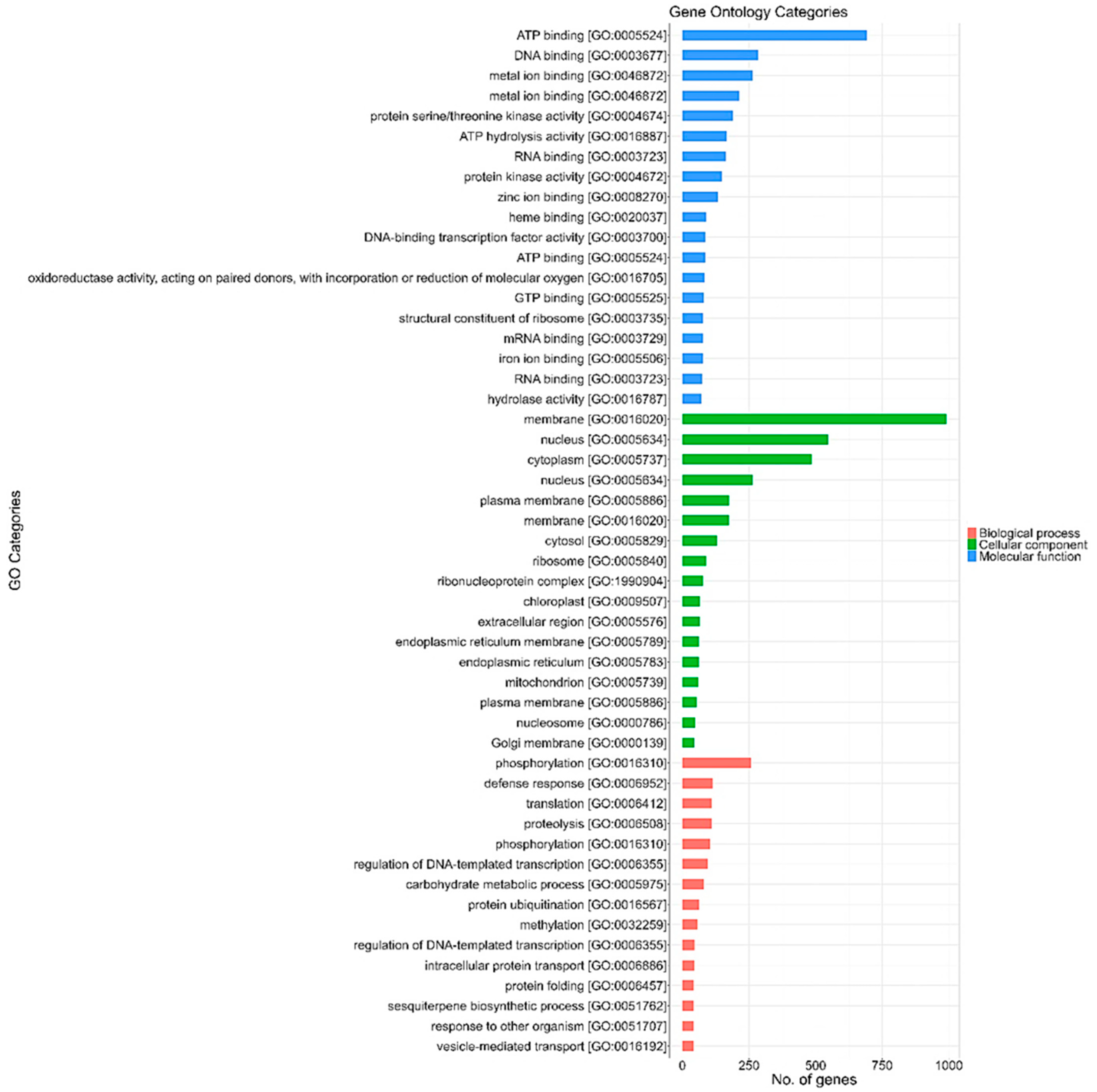
3.4. Functional Annotation and Gene Ontology (GO) Sequence Distribution
3.5. Differential Gene Expression Analysis
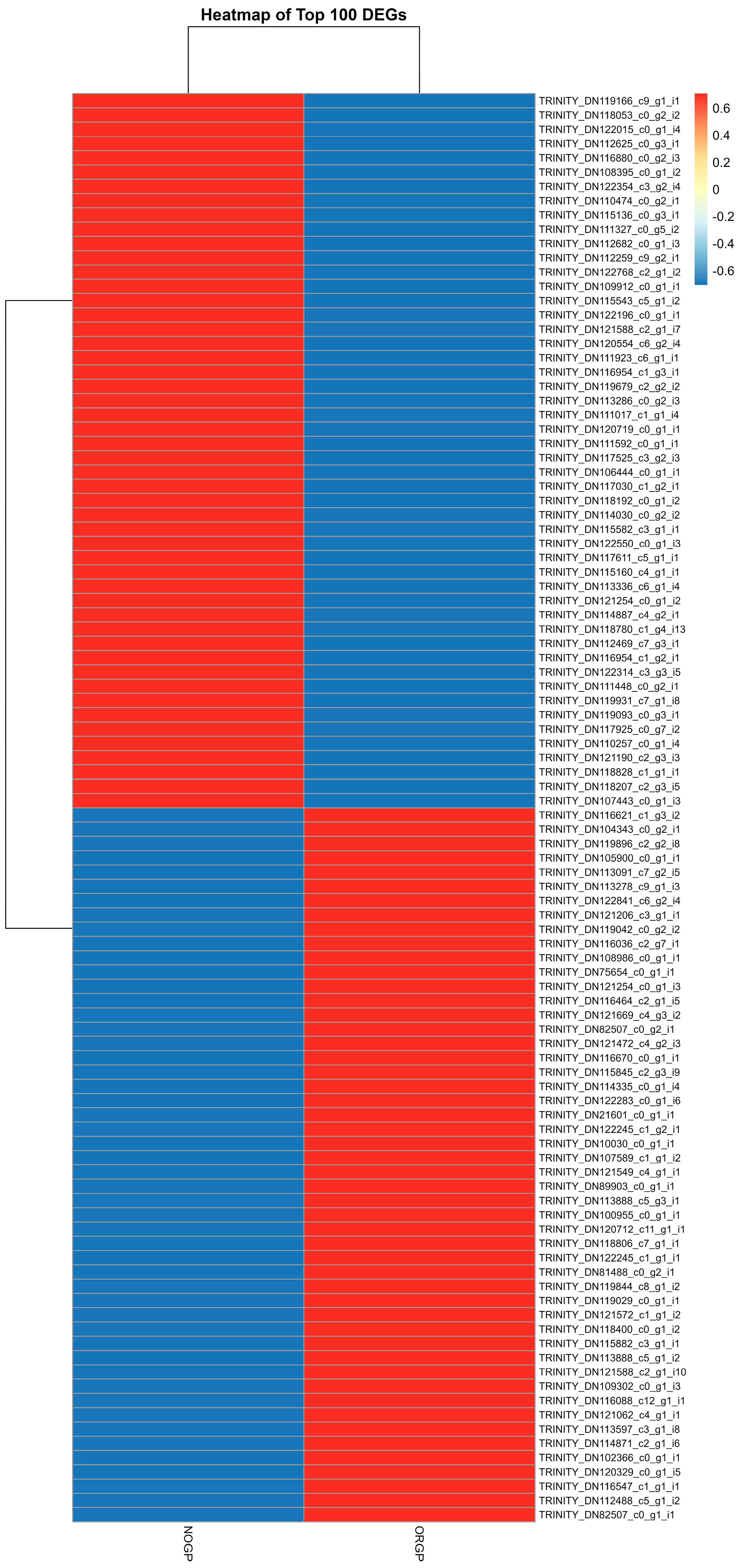
3.6. Heatmap of Differentially Expressed Genes
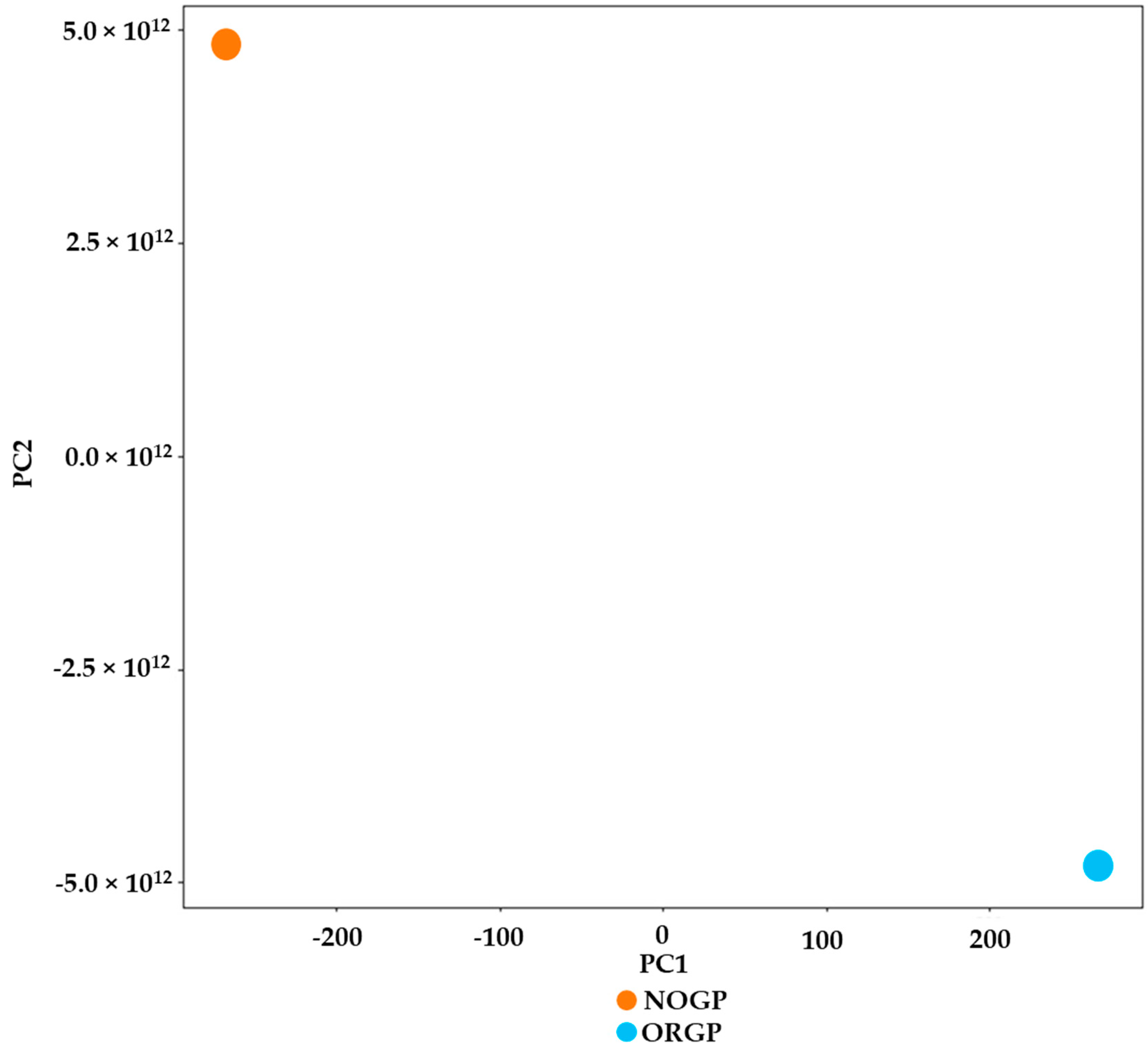
3.7. Principal Component Analysis (PCA) Plot
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gawade, N.; Bhalekar, S.G.; Bhosale, P.; Katwate, S.M.; Wadekar, V. Studies on different genotypes of Gaillardia (Gaillardia pulchella L.) for quantitative and qualitative performance. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1030–1039. [Google Scholar] [CrossRef]
- Nagy, K.N.; Kardos, L.V.; Orbán, Z.; Bakacsy, L. The allelochemical potential of an invasive ornamental plant, the Indian blanket flower (Gaillardia pulchella Foug.). Plant Species Biol. 2023, 39, 102–108. [Google Scholar] [CrossRef]
- Kadam, M.; Malshe, K.; Salvi, B.; Chavan, S. Effect of plant growth regulators on flowering and flower yield in Gaillardia (Gaiilardia pulchella) cv. Local Double. Int. J. Chem. Stud. 2020, 8, 927–930. [Google Scholar] [CrossRef]
- El-Khateeb, M.; Ashour, H.; Eid, R.; Mahfouze, H.; Abd Elaziz, N.; Radwan Ragab, M.S. Induction of genetic variability with gamma radiation and detection of DNA polymorphisms among radio mutants using sequence-related amplified polymorphism markers in Gaillardia pulchella Foug. plants. Egypt. Pharmaceut. J. 2023, 22, 272. [Google Scholar] [CrossRef]
- Yao, X.T.; Ling, P.X.; Jiang, S.; Lai, P.X.; Zhu, C.G. Analysis of the essential oil from Gaillardia pulchella Foug. and its antioxidant activity. J. Oleo Sci. 2013, 62, 329–333. [Google Scholar] [CrossRef]
- Moharram, F.A.; Dib, R.A.E.M.E.; Marzouk, M.S.; El-Shenawy, S.M.; Ibrahim, H.A. New apigenin glycoside, polyphenolic constituents, anti-Inflammatory and hepatoprotective activities of Gaillardia grandiflora and Gaillardia pulchella aerial Parts. Pharmacogn. Mag. 2017, 13, 244. [Google Scholar] [CrossRef]
- Bosco, A.; Molina, L.; Kernéis, S.M.; Hatzopoulos, G.; Favez, T.; Gonczy, P.; Tantapakul, C.; Maneerat, W.; Yeremy, B.; Williams, D.E.; et al. Pulchelloid A, a sesquiterpene lactone from the Canadian prairie plant Gaillardia aristata inhibits mitosis in human cells. Mol. Biol. Rep. 2021, 48, 5459–5471. [Google Scholar] [CrossRef]
- Bansal, Y.; Mujib, A.; Siddiqui, Z.H.; Mamgain, J.; Syeed, R.; Ejaz, B. Ploidy status, nuclear DNA content and start codon targeted (SCOT) genetic homogeneity assessment in Digitalis purpurea L., regenerated in vitro. Genes 2022, 13, 2335. [Google Scholar] [CrossRef]
- Norouzi, O.; Hesami, M.; Pepe, M.; Dutta, A.; Jones, A.M.P. In vitro plant tissue culture as the fifth generation of bioenergy. Sci. Rep. 2022, 12, 5038. [Google Scholar] [CrossRef]
- Cantabella, D.; Dolcet-Sanjuan, R.; Teixidó, N. Using plant growth-promoting microorganisms (PGPMs) to improve plant development under in vitro culture conditions. Planta 2022, 255, 117. [Google Scholar] [CrossRef]
- Shin, J.; Bae, S.; Seo, P.J. De novo shoot organogenesis during plant regeneration. Bot. Exp. Bot. 2020, 71, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Hnatuszko-Konka, K.; Gerszberg, A.; Weremczuk-Jeżyna, I.; Grzegorczyk-Karolak, I. Cytokinin signaling and de novo shoot organogenesis. Genes 2021, 12, 265. [Google Scholar] [CrossRef]
- Nadiya, F.; Anjali, N.; Thomas, J.; Gangaprasad, A.; Sabu, K.K. Genome-wide differential expression profiling in wild and cultivar genotypes of cardamom reveals regulation of key pathways in plant growth and development. Agric. Gene 2018, 8, 18–27. [Google Scholar]
- Chen, S.; Xu, X.; Ma, Z.; Liu, J.; Zhang, B. Organ-specific transcriptome analysis identifies candidate genes involved in the stem specialization of bermudagrass (Cynodon dactylon L.). Front. Genet. 2021, 12, 678673. [Google Scholar] [CrossRef] [PubMed]
- Torres-Silva, G.; Correia, L.N.F.; Batista, D.S.; Koehler, A.D.; Resende, S.V.; Romanel, E.; Cassol, D.; Almeida, A.M.R.; Strickler, S.R.; Specht, C.D.; et al. Transcriptome analysis of Melocactus glaucescens (Cactaceae) reveals metabolic changes during in vitro shoot organogenesis induction. Front. Plant Sci. 2021, 12, 697556. [Google Scholar] [CrossRef]
- Schellenbaum, P.; Jacques, A.; Maillot, P.; Bertsch, C.; Mazet, F.; Farine, S.; Walter, B. Characterization of VvSERK1, VvSERK2, VvSERK3 and VvL1L Genes and their expression during somatic embryogenesis of grapevine (Vitis vinifera L.). Plant Cell Rep. 2008, 27, 1799–1809. [Google Scholar] [CrossRef] [PubMed]
- Salaün, C.; Lepiniec, L.; Dubreucq, B. Genetic and molecular control of somatic embryogenesis. Plants 2021, 10, 1467. [Google Scholar] [CrossRef]
- Huang, X.; Chen, J.; Bao, Y.; Liu, L.; Jiang, H.; An, X.; Dai, L.; Wang, B.; Peng, D. Transcript profiling reveals auxin and cytokinin signaling pathways and transcription regulation during In vitro organogenesis of ramie (Boehmeria nivea L. Gaud). PLoS ONE 2014, 9, e113768. [Google Scholar] [CrossRef] [PubMed]
- Puvvala, S.S.; Muddanuru, T.; Thangella, P.A.V.; Kumar, O.A.; Chakravartty, N.; Vettath, V.K.; Katta, A.V.S.K.M.; Lekkala, S.P.; Kuriakose, B.; Gupta, S.; et al. Deciphering the transcriptomic insight during organogenesis in castor (Ricinus communis L.), jatropha (Jatropha curcas L.) and sunflower (Helianthus annuus L.). 3 Biotech 2019, 9, 434. [Google Scholar] [CrossRef]
- Tu, M.; Wang, W.; Yao, N.; Cai, C.; Liu, Y.; Lin, C.; Zuo, Z.; Zhu, Q. The transcriptional dynamics during de novo shoot organogenesis of Ma bamboo (Dendrocalamus latiflorus Munro): Implication of the contributions of the abiotic stress response in this process. Plant J. 2021, 107, 1513–1532. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: Cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef] [PubMed]
- Bansal, M.; Mujib, A.; Bansal, Y.; Dewir, Y.H.; Mendler-Drienyovszki, N. An efficient In vitro shoot organogenesis and comparative GC-MS metabolite profiling of Gaillardia pulchella Foug. Horticulturae 2024, 10, 728. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Cervantes-Pérez, S.A.; Espinal-Centeno, A.; Oropeza-Aburto, A.; Caballero-Pérez, J.; Falcon, F.; Aragón-Raygoza, A.; Sánchez-Segura, L.; Herrera-Estrella, L.; Cruz-Hernández, A.; Cruz-Ramírez, A. Transcriptional profiling of the CAM plant Agave salmiana reveals conservation of a genetic program for regeneration. Dev. Biol. 2018, 442, 28–39. [Google Scholar] [CrossRef]
- Abe, M.; Kobayashi, Y.; Yamamoto, S.; Daimon, Y.; Yamaguchi, A.; Ikeda, Y.; Ichinoki, H.; Notaguchi, M.; Goto, K.; Araki, T. FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 2005, 309, 1052–1056. [Google Scholar] [CrossRef]
- Cohen, O.; Borovsky, Y.; David-Schwartz, R.; Paran, I. CaJOINTLESS is a MADS-Box gene involved in suppression of vegetative growth in all shoot meristems in pepper. J. Exp. Bot. 2012, 63, 4947–4957. [Google Scholar] [CrossRef]
- Liu, T.; Zhu, S.; Tang, Q.; Tang, S. Identification of 32 Full-Length NAC Transcription factors in ramie (Boehmeria nivea L. Gaud) and characterization of the expression pattern of these genes. Mol. Genet. Genom. 2014, 289, 675–684. [Google Scholar] [CrossRef]
- Schuster, C.; Gaillochet, C.; Medzihradszky, A.; Busch, W.; Daum, G.; Krebs, M.; Kehle, A.; Lohmann, J.U. A Regulatory framework for shoot stem cell control integrating metabolic, transcriptional, and phytohormone signals. Dev. Cell 2014, 28, 438–449. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Favero, D.S.; Sakamoto, Y.; Iwase, A.; Coleman, D.; Rymen, B.; Sugimoto, K. Molecular mechanisms of plant regeneration. Annu. Rev. Plant Biol. 2019, 70, 377–406. [Google Scholar] [CrossRef]
- Zhang, T.-Q.; Lian, H.; Zhou, C.-M.; Xu, L.; Jiao, Y.; Wang, J.-W. A two-step model for de novo activation of WUSCHEL during plant shoot regeneration. Plant Cell 2017, 29, 1073–1087. [Google Scholar] [CrossRef] [PubMed]
- Subramaniyam, S.; Mathiyalagan, R.; Gyo, I.J.; Bum-Soo, L.; Sungyoung, L.; Chun, Y.D. Transcriptome Profiling and In silico Analysis of Gynostemma pentaphyllum using a next generation sequencer. Plant Cell Rep. 2011, 30, 2075–2083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.; Han, S.; Li, X.; Qi, L. Transcriptome profiling and in silico analysis of somatic embryos in Japanese larch (Larix leptolepis). Plant Cell Rep. 2012, 31, 1637–1657. [Google Scholar] [CrossRef] [PubMed]
- Norikane, A.; Da Silva, J.A.T.; Tanaka, M. Growth of in vitro Oncidesa plantlets cultured under cold cathode fluorescent lamps with super-elevated CO2 enrichment. AoB Plants 2013, 5, plt044. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.-L.; Guo, H.-R.; Xie, L.; Zeng, R.-Z.; Zhang, X.-Q.; Zhang, Z.-S. Transcriptomic and hormonal analyses reveal that YUC-mediated auxin biogenesis is involved in shoot regeneration from rhizome in Cymbidium. Front. Plant Sci. 2017, 8, 1866. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Non-Organogenic Callus | Organogenic Callus |
|---|---|---|
| Raw reads | 26.2 | 23.6 |
| Clean reads | 21.4 | 18.4 |
| Total trinity transcripts | 117,149 | 101,444 |
| Total assembled bases | 64,653,174 | 53,724,847 |
| Average contig | 551.89 | 529.6 |
| Contig N50 | 540 | 542 |
| Trinity Id | NOGP | ORGP | Protein Names | Gene Names |
|---|---|---|---|---|
| TRINITY_DN119166_c9_g1_i1 | 6.919624094 | −1.132747354 | dynamin GTPase (EC 3.6.5.5) | E3N88_28147 |
| TRINITY_DN118053_c0_g2_i2 | 6.475540755 | −1.132747354 | Putative developmentally-regulated GTP-binding protein 1 (Small GTP-binding protein) | DRG1 HannXRQ_Chr14g0446081 HanXRQr2_Chr14g0648391 |
| TRINITY_DN122015_c0_g1_i4 | 6.031761186 | −1.132747354 | DNA/RNA-binding protein Kin17 WH-like domain-containing protein | LSAT_3X22041 |
| TRINITY_DN112625_c0_g3_i1 | 5.849473051 | −1.132747354 | AAA+ ATPase domain-containing protein | E3N88_06826 |
| TRINITY_DN116880_c0_g2_i3 | 5.832697338 | −1.132747354 | histidine kinase (EC 2.7.13.3) | WOL HannXRQ_Chr08g0224601 HanXRQr2_Chr08g0338301 |
| TRINITY_DN108395_c0_g1_i2 | 5.833034771 | −1.132747354 | Putative zinc finger (Ubiquitin-hydrolase) domain-containing protein (Transcription factor C2H2 family) | BRIZ1 HannXRQ_Chr05g0138581 HanXRQr2_Chr05g0206571 |
| TRINITY_DN122354_c3_g2_i4 | 5.816236368 | −1.132747354 | Peptidase A1 domain-containing protein | E3N88_37546 |
| TRINITY_DN110474_c0_g2_i1 | 7.151584978 | −1.132747354 | BURP domain-containing protein | E3N88_05164 |
| TRINITY_DN115136_c0_g3_i1 | 7.15967866 | −1.132747354 | Putative U5 small nuclear ribonucleoprotein helicase, putative (RNA helicase (EC 3.6.4.13)) | HannXRQ_Chr17g0533721 HanXRQr2_Chr17g0778891 |
| TRINITY_DN111327_c0_g5_i2 | 6.494756219 | −1.132747354 | Methyltransferase | LSAT_5X175200 |
| TRINITY_DN112682_c0_g1_i3 | 5.698025802 | −1.132747354 | Putative homeodomain-like, DEK | HannXRQ_Chr05g0159891 |
| TRINITY_DN112259_c9_g2_i1 | 5.648847889 | −1.132747354 | DUF4378 domain-containing protein | E3N88_24855 |
| TRINITY_DN122768_c2_g1_i2 | 6.475540755 | −1.132747354 | ABC-type xenobiotic transporter (EC 7.6.2.2) | ATMRP14 HannXRQ_Chr14g0462711 HanXRQr2_Chr14g0670341 |
| TRINITY_DN109912_c0_g1_i1 | 5.716066628 | −1.132747354 | Putative chaperone protein DnaJ (Terminal organelle assembly protein TopJ) | DNAJ HannXRQ_Chr04g0117371 HanXRQr2_Chr04g0180601 |
| TRINITY_DN115543_c5_g1_i2 | 5.690969228 | −1.132747354 | nonspecific serine/threonine protein kinase (EC 2.7.11.1) | E3N88_26722 |
| TRINITY_DN122196_c0_g1_i1 | 7.122065892 | −1.132747354 | PHD-type domain-containing protein | E3N88_06438 |
| TRINITY_DN121588_c2_g1_i7 | 7.526502169 | −1.132747354 | NK | NK |
| TRINITY_DN120554_c6_g2_i4 | 6.759140731 | −1.132747354 | NK | NK |
| TRINITY_DN111923_c6_g1_i1 | 6.397838398 | −1.132747354 | nonspecific serine/threonine protein kinase (EC 2.7.11.1) | E3N88_37668 |
| TRINITY_DN116954_c1_g3_i1 | 6.397838398 | −1.132747354 | Nucleoprotein TPR/MLP1 (Putative nuclear pore anchor) | NUA HannXRQ_Chr14g0459251 HanXRQr2_Chr12g0534391 |
| TRINITY_DN119679_c2_g2_i2 | 6.956948069 | −1.132747354 | NK | NK |
| TRINITY_DN113286_c0_g2_i3 | 6.878369196 | −1.132747354 | NAB domain-containing protein | HannXRQ_Chr03g0072561 HanXRQr2_Chr03g0100501 |
| TRINITY_DN111017_c1_g1_i4 | 6.029849065 | −1.132747354 | NK | NK |
| TRINITY_DN120719_c0_g1_i1 | 6.036896642 | −1.132747354 | Pentacotripeptide-repeat region of PRORP domain-containing protein | LSAT_6X30921 |
| TRINITY_DN111592_c0_g1_i1 | 5.992273651 | −1.132747354 | TRAF-like protein | CTI12_AA002210 |
| TRINITY_DN117525_c3_g2_i3 | 6.738124811 | −1.132747354 | AAA+ ATPase domain-containing protein | LSAT_8X58161 |
| TRINITY_DN106444_c0_g1_i1 | 5.812476677 | −1.132747354 | GOLD domain-containing protein | HannXRQ_Chr11g0329321 HanXRQr2_Chr11g0496721 |
| TRINITY_DN117030_c1_g2_i1 | 5.702095466 | −1.132747354 | nonspecific serine/threonine protein kinase (EC 2.7.11.1) | CTI12_AA553500 |
| TRINITY_DN118192_c0_g1_i2 | 5.714785453 | −1.132747354 | NK | NK |
| TRINITY_DN114030_c0_g2_i2 | 5.653249562 | −1.132747354 | Putative SART-1 family (SNU66/SART1 family protein) | DOT2 HannXRQ_Chr09g0244631 HanXRQr2_Chr09g0371141 |
| TRINITY_DN115582_c3_g1_i1 | 5.653249562 | −1.132747354 | Uncharacterized protein | HannXRQ_Chr05g0147041 |
| TRINITY_DN122550_c0_g1_i3 | 5.658399666 | −1.132747354 | Coatomer subunit beta’ | RHSIM_Rhsim08G0220500 |
| TRINITY_DN117611_c5_g1_i1 | 5.634013343 | −1.132747354 | histone acetyltransferase (EC 2.3.1.48) | HannXRQ_Chr05g0150621 HanXRQr2_Chr05g0221231 |
| TRINITY_DN115160_c4_g1_i1 | 5.553986842 | −1.132747354 | Major facilitator superfamily (MFS) profile domain-containing protein | E3N88_10857 |
| TRINITY_DN113336_c6_g1_i4 | 5.634013343 | −1.132747354 | SWI/SNF complex subunit SWI3D | E3N88_34386 |
| TRINITY_DN121254_c0_g1_i2 | 7.282803045 | −1.132747354 | Putative eukaryotic molybdopterin oxidoreductase, Immunoglobulin E-set | HannXRQ_Chr07g0198211 |
| TRINITY_DN114887_c4_g2_i1 | 6.518132631 | −1.132747354 | 1,3-beta-glucan synthase (EC 2.4.1.34) (1,3-beta-glucan synthase) | ATGSL08 HannXRQ_Chr09g0272811 HanXRQr2_Chr09g0411381 |
| TRINITY_DN118780_c1_g4_i13 | 6.46142319 | −1.132747354 | NK | NK |
| TRINITY_DN112469_c7_g3_i1 | 7.370146001 | −1.132747354 | NK | NK |
| TRINITY_DN116954_c1_g2_i1 | 6.094324922 | −1.132747354 | NK | NK |
| TRINITY_DN122314_c3_g3_i5 | 5.977087247 | −1.132747354 | NK | NK |
| TRINITY_DN111448_c0_g2_i1 | 5.961739283 | −1.132747354 | Acyl-CoA dehydrogenase family member 11 | Ccrd_002843 |
| TRINITY_DN119931_c7_g1_i8 | 5.8978544 | −1.132747354 | Nepenthesin (EC 3.4.23.12) (Putative eukaryotic aspartyl protease family protein) | HannXRQ_Chr05g0136621 HanXRQr2_Chr00c091g0833781 |
| TRINITY_DN119093_c0_g3_i1 | 5.866055935 | −1.132747354 | LisH domain-containing protein | E3N88_33932 |
| TRINITY_DN117925_c0_g7_i2 | 5.698211036 | −1.132747354 | Amino acid transporter transmembrane domain-containing protein | E3N88_30362 |
| TRINITY_DN110257_c0_g1_i4 | 7.199937594 | −1.132747354 | Uncharacterized protein | E3N88_30651 |
| TRINITY_DN121190_c2_g3_i3 | 5.656303694 | −1.132747354 | Hexosyltransferase (EC 2.4.1.-) | E3N88_10478 |
| TRINITY_DN118828_c1_g1_i1 | 5.594753913 | −1.132747354 | Smr domain-containing protein | E3N88_11151 |
| TRINITY_DN118207_c2_g3_i5 | 6.349007448 | −1.132747354 | Heat shock protein 70 family (Putative heat shock protein 70 (Hsp 70) family protein) | HannXRQ_Chr17g0541391 HanXRQr2_Chr17g0789871 |
| TRINITY_DN107443_c0_g1_i3 | 7.097839407 | −1.132747354 | EH domain, EF-hand domain pair protein (Putative EF-hand domain pair) | HannXRQ_Chr14g0442891 HanXRQr2_Chr14g0644141 |
| TRINITY_DN116621_c1_g3_i2 | −1.585155177 | 6.518304338 | Ribosomal protein L2 C-terminal domain-containing protein | COCSUDRAFT_9852 COCSUDRAFT_9853 |
| TRINITY_DN104343_c0_g2_i1 | −1.585155177 | 6.414147106 | CAP superfamily protein (Putative pathogenesis-related protein 1B) | PR1B HannXRQ_Chr04g0096331 HanXRQr2_Chr06g0269641 |
| TRINITY_DN119896_c2_g2_i8 | −1.585155177 | 6.156164849 | Uncharacterized protein | E3N88_15356 |
| TRINITY_DN105900_c0_g1_i1 | −1.585155177 | 6.861606083 | NK | NK |
| TRINITY_DN113091_c7_g2_i5 | −1.585155177 | 6.861606083 | Putative NAC domain containing protein 83 (Transcription factor NAM family) | ANAC083 HannXRQ_Chr12g0362371 HanXRQr2_Chr12g0530861 |
| TRINITY_DN113278_c9_g1_i3 | −1.585155177 | 7.491207275 | MAPK kinase substrate protein | CTI12_AA167510 |
| TRINITY_DN122841_c6_g2_i4 | −1.585155177 | 7.952061034 | NADH-ubiquinone oxidoreductase chain 1 | NK |
| TRINITY_DN121206_c3_g1_i1 | −1.585155177 | 7.011910889 | NK | NK |
| TRINITY_DN119042_c0_g2_i2 | −1.585155177 | 6.546028821 | Putative UDP-glycosyltransferase 76G1 (UDP-glucuronosyl/UDP-glucosyltransferase) | U76G1 HannXRQ_Chr14g0458621 HanXRQr2_Chr14g0665451 |
| TRINITY_DN116036_c2_g7_i1 | −1.585155177 | 6.588351835 | Putative ypt/Rab-GAP domain of gyp1p superfamily protein (Rab-GTPase-TBC domain-containing protein) | HannXRQ_Chr07g0206381 HanXRQr2_Chr07g0317121 |
| TRINITY_DN108986_c0_g1_i1 | −1.585155177 | 6.595445282 | Pectinesterase (EC 3.1.1.11) | RHS12 HannXRQ_Chr07g0189571 HanXRQr2_Chr07g0289991 |
| TRINITY_DN75654_c0_g1_i1 | −1.585155177 | 6.743769593 | NK | NK |
| TRINITY_DN121254_c0_g1_i3 | −1.585155177 | 7.571709666 | Putative eukaryotic molybdopterin oxidoreductase, Immunoglobulin E-set | HannXRQ_Chr07g0198211 |
| TRINITY_DN116464_c2_g1_i5 | −1.585155177 | 7.952061034 | glutathione transferase (EC 2.5.1.18) | GSTF HannXRQ_Chr04g0118231 HanXRQr2_Chr04g0177751 |
| TRINITY_DN121669_c4_g3_i2 | −1.585155177 | 7.166460665 | Laccase (EC 1.10.3.2) (Benzenediol:oxygen oxidoreductase) (Diphenol oxidase) (Urishiol oxidase) | E3N88_34257 |
| TRINITY_DN82507_c0_g2_i1 | −1.585155177 | 8.117551064 | NK | NK |
| TRINITY_DN121472_c4_g2_i3 | −1.585155177 | 9.372607705 | RRM domain-containing protein | Golob_000306 |
| TRINITY_DN116670_c0_g1_i1 | −1.585155177 | 7.75294902 | NK | NK |
| TRINITY_DN115845_c2_g3_i9 | −1.585155177 | 8.874700463 | Auxin-responsive protein | E3N88_39490 |
| TRINITY_DN114335_c0_g1_i4 | −1.585155177 | 9.545113204 | Bifunctional inhibitor/plant lipid transfer protein/seed storage helical (Putative bifunctional inhibitor/lipid-transfer protein/seed storage 2S albumin superfamily protein) | HannXRQ_Chr17g0549921 HanXRQr2_Chr17g0801201 |
| TRINITY_DN122283_c0_g1_i6 | −1.585155177 | 9.579040055 | Peroxidase (EC 1.11.1.7) | LSAT_7X16000 |
| TRINITY_DN21601_c0_g1_i1 | −1.585155177 | 9.370084451 | Neuroendocrine convertase 2-like | LOC115749069 |
| TRINITY_DN122245_c1_g2_i1 | −1.585155177 | 14.83400592 | NK | NK |
| TRINITY_DN10030_c0_g1_i1 | −1.585155177 | 9.008082417 | NK | NK |
| TRINITY_DN107589_c1_g1_i2 | −1.585155177 | 9.803154329 | Furin-like protease 1 | LOC115751520 |
| TRINITY_DN121549_c4_g1_i1 | −1.585155177 | 9.602029997 | NK | NK |
| TRINITY_DN89903_c0_g1_i1 | −1.585155177 | 9.611245507 | Peptidase S1 domain-containing protein | OSTQU699_LOCUS8363 |
| TRINITY_DN113888_c5_g3_i1 | −1.585155177 | 10.85487196 | NK | NK |
| TRINITY_DN100955_c0_g1_i1 | −1.585155177 | 9.672190318 | NK | NK |
| TRINITY_DN120712_c11_g1_i1 | −1.585155177 | 11.41197564 | NK | NK |
| TRINITY_DN118806_c7_g1_i1 | −1.585155177 | 10.87392936 | Furin-like protease 2 | LOC115745509 |
| TRINITY_DN122245_c1_g1_i1 | −1.585155177 | 14.32921163 | NK | NK |
| TRINITY_DN81488_c0_g2_i1 | −1.585155177 | 12.43104475 | NK | NK |
| TRINITY_DN119844_c8_g1_i2 | −1.585155177 | 6.150618859 | NK | NK |
| TRINITY_DN119029_c0_g1_i1 | −1.585155177 | 6.179954119 | Long-chain-fatty-acid--CoA ligase (EC 6.2.1.3) | LACS4 HannXRQ_Chr03g0083551 HanXRQr2_Chr03g0129061 |
| TRINITY_DN121572_c1_g1_i2 | −1.585155177 | 6.258195419 | Protein TAR1 | E3N88_44475 |
| TRINITY_DN118400_c0_g1_i2 | −1.585155177 | 6.370601382 | Transmembrane protein 214-A | LSAT_4X13621 |
| TRINITY_DN115882_c3_g1_i1 | −1.585155177 | 8.527248539 | NK | NK |
| TRINITY_DN113888_c5_g1_i2 | −1.585155177 | 8.52367751 | NK | NK |
| TRINITY_DN121588_c2_g1_i10 | −1.585155177 | 8.782205775 | Uncharacterized protein | E3N88_28192 |
| TRINITY_DN109302_c0_g1_i3 | −1.585155177 | 7.35509268 | K Homology domain-containing protein | E3N88_08574 |
| TRINITY_DN116088_c12_g1_i1 | −1.585155177 | 6.006803999 | Succinate dehydrogenase [ubiquinone] iron-sulfur subunit, mitochondrial (EC 1.3.5.1) | LSAT_4X184781 |
| TRINITY_DN121062_c4_g1_i1 | −1.585155177 | 6.112139706 | Uncharacterized protein | E3N88_31121 |
| TRINITY_DN113597_c3_g1_i8 | −1.585155177 | 6.90122619 | Uncharacterized protein | LSAT_7X71780 |
| TRINITY_DN114871_c2_g1_i6 | −1.585155177 | 7.263000975 | Glycine rich protein | Ccrd_010946 |
| TRINITY_DN102366_c0_g1_i1 | −1.585155177 | 6.960009787 | isocitrate lyase (EC 4.1.3.1) | E3N88_04782 |
| TRINITY_DN120329_c0_g1_i5 | −1.585155177 | 6.268132083 | Uncharacterized protein | GLYMA_13G011700 |
| TRINITY_DN116547_c1_g1_i1 | −1.585155177 | 6.398634107 | Diphosphomevalonate decarboxylase (EC 4.1.1.33) | HannXRQ_Chr11g0342191 HanXRQr2_Chr14g0660761 |
| TRINITY_DN112488_c5_g1_i2 | −1.585155177 | 7.842412771 | Tuber agglutinin | hta-c |
| TRINITY_DN82507_c0_g1_i1 | −1.585155177 | 7.638742116 | NK | NK |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bansal, Y.; Mujib, A.; Bansal, M.; Mohsin, M.; Nafees, A.; Dewir, Y.H. Comparative Transcriptome Analysis of Non-Organogenic and Organogenic Tissues of Gaillardia pulchella Revealing Genes Regulating De Novo Shoot Organogenesis. Horticulturae 2024, 10, 1138. https://doi.org/10.3390/horticulturae10111138
Bansal Y, Mujib A, Bansal M, Mohsin M, Nafees A, Dewir YH. Comparative Transcriptome Analysis of Non-Organogenic and Organogenic Tissues of Gaillardia pulchella Revealing Genes Regulating De Novo Shoot Organogenesis. Horticulturae. 2024; 10(11):1138. https://doi.org/10.3390/horticulturae10111138
Chicago/Turabian StyleBansal, Yashika, A. Mujib, Mahima Bansal, Mohammad Mohsin, Afeefa Nafees, and Yaser Hassan Dewir. 2024. "Comparative Transcriptome Analysis of Non-Organogenic and Organogenic Tissues of Gaillardia pulchella Revealing Genes Regulating De Novo Shoot Organogenesis" Horticulturae 10, no. 11: 1138. https://doi.org/10.3390/horticulturae10111138
APA StyleBansal, Y., Mujib, A., Bansal, M., Mohsin, M., Nafees, A., & Dewir, Y. H. (2024). Comparative Transcriptome Analysis of Non-Organogenic and Organogenic Tissues of Gaillardia pulchella Revealing Genes Regulating De Novo Shoot Organogenesis. Horticulturae, 10(11), 1138. https://doi.org/10.3390/horticulturae10111138