
Strategies for Supplying Precursors to Enhance the Production of Secondary Metabolites in Solid-State Fermentation

 ,
,  , and
, and
Abstract
:1. Introduction
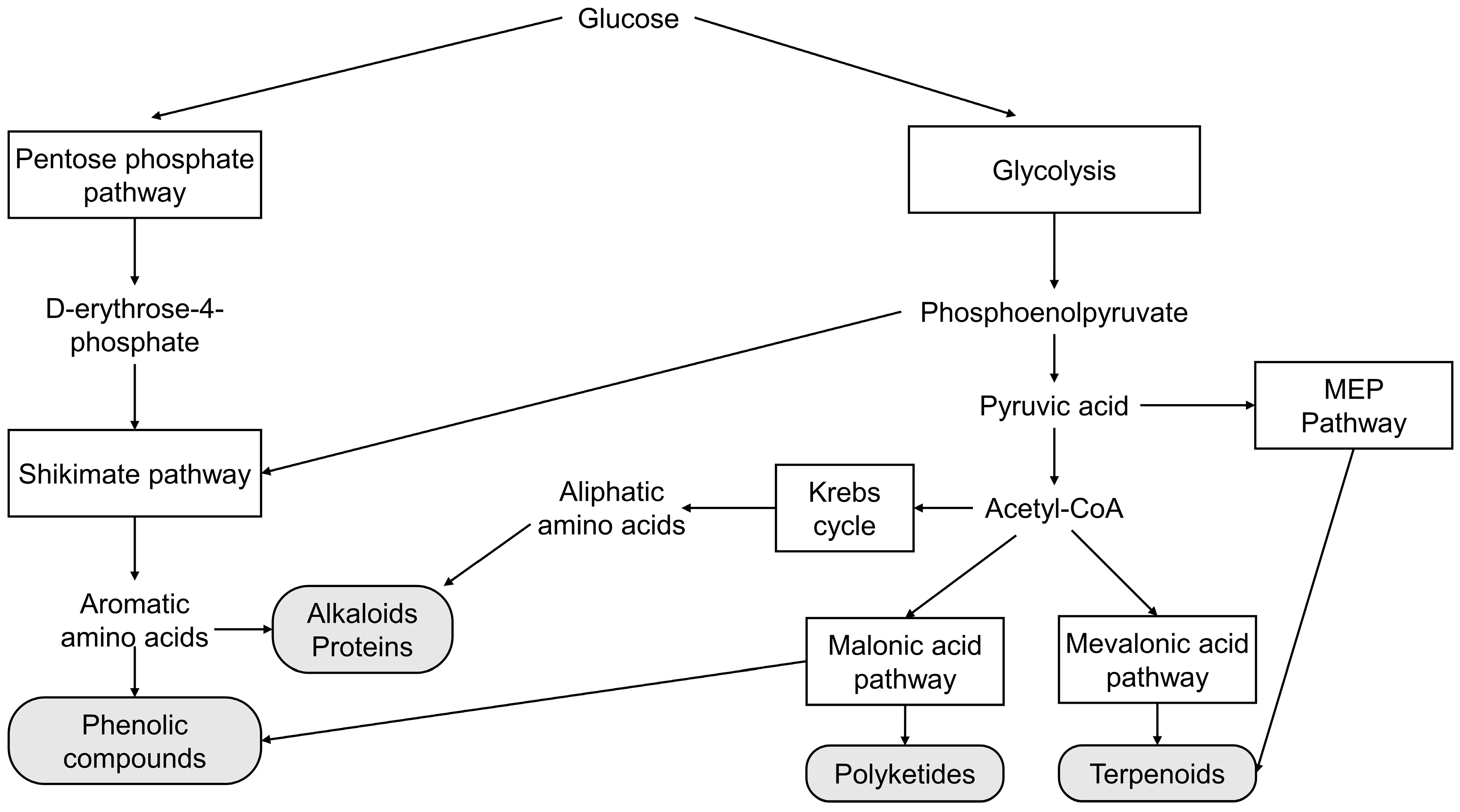
2. Main Biosynthetic Pathways of Secondary Metabolism
2.1. Shikimate Pathway
2.2. Malonic Acid Pathway
2.3. Mevalonic Acid Pathway (MVA)
2.4. Methylerythritol-Phosphate (MEP) Pathway
3. Using Precursors to Enhance Secondary Metabolite Production
4. Enhancing Secondary Metabolite Production in SSF
4.1. Stimulating Internal Precursor Generation
4.2. Using Precursors That Are Already Content in the Solid Material
4.3. Adding External Precursors to SSF
5. Discussion
6. Concluding Remarks and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, V.; Ahluwalia, V.; Saran, S.; Kumar, J.; Patel, A.K.; Singhania, R.R. Recent Developments on Solid-State Fermentation for Production of Microbial Secondary Metabolites: Challenges and Solutions. Bioresour. Technol. 2021, 323, 124566. [Google Scholar] [CrossRef] [PubMed]
- Fouillaud, M.; Dufossé, L. Microbial Secondary Metabolism and Biotechnology. Microorganisms 2022, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Sahu, P.K.; Agarwal, V.; Singh, N. Exploiting Endophytic Microbes as Micro-Factories for Plant Secondary Metabolite Production. Appl. Microbiol. Biotechnol. 2021, 105, 6579–6596. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring Structural Diversity of Microbe Secondary Metabolites Using OSMAC Strategy: A Literature Review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Durán, L.V.; Michel, M.R.; Pichardo, A.; Aguilar-Zárate, P. Microbial Bioreactors for Secondary Metabolite Production. In Microbial Bioreactors for Industrial Molecules; Wiley: Hoboken, NJ, USA, 2023; pp. 275–296. [Google Scholar] [CrossRef]
- Cerda-Cejudo, N.D.; Buenrostro-Figueroa, J.J.; Sepúlveda-Torre, L.; Torres-León, C.; Chávez-González, M.L.; Ascacio-Valdés, J.A.; Aguilar, C.N. Solid-State Fermentation for the Recovery of Phenolic Compounds from Agro-Wastes. Resources 2023, 12, 36. [Google Scholar] [CrossRef]
- Soccol, C.R.; Costa, E.S.F.d.; Letti, L.A.J.; Karp, S.G.; Woiciechowski, A.L.; Vandenberghe, L.P.d.S. Recent Developments and Innovations in Solid State Fermentation. Biotechnol. Res. Innov. 2017, 1, 52–71. [Google Scholar] [CrossRef]
- Barrios-González, J. Secondary Metabolites Production: Physiological Advantages in Solid-State Fermentation. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 257–283. [Google Scholar]
- Jazmín Edith, M.-H.; Octavio, L.; Edna Madai, M.-H.; Esperanza, H.; Nicolás Óscar, S.-C. Coupling Energy-Production Processes: The Use of Residues from Bioethanol Production to Improve the Anaerobic Digestion of Corn Stover. Biomass Bioenergy 2019, 128, 105322. [Google Scholar] [CrossRef]
- Sainz-Mellado, D.C.; Méndez-Hernández, J.E.; López-Miranda, J.; Páez-Lerma, J.B.; Aguilar, C.N.; Soto-Cruz, N.O. Gradually Supply of Isoamyl Alcohol Increases the Isoamyl Acetate Production in Solid-State Fermentation. Lett. Appl. Microbiol. 2023, 76, ovac061. [Google Scholar] [CrossRef]
- Barrios-González, J. Solid-State Fermentation: Physiology of Solid Medium, Its Molecular Basis and Applications. Process Biochem. 2012, 47, 175–185. [Google Scholar] [CrossRef]
- Chaudhary, V.; Katyal, P.; Puniya, A.K.; Panwar, H.; Arora, M.; Kaur, J.; Rokana, N.; Wakchaure, N.; Raposo, A.; Raheem, D.; et al. Pilot-Scale Process to Produce Bio-Pigment from Monascus Purpureus Using Broken Rice as Substrate for Solid-State Fermentation. Eur. Food Res. Technol. 2023, 249, 1845–1855. [Google Scholar] [CrossRef]
- Srianta, I.; Zubaidah, E.; Estiasih, T.; Yamada, M.; Harijono. Comparison of Monascus purpureus Growth, Pigment Production and Composition on Different Cereal Substrates with Solid State Fermentation. Biocatal. Agric. Biotechnol. 2016, 7, 181–186. [Google Scholar] [CrossRef]
- Domínguez, M.; Mejía, A.; Revah, S.; Barrios-González, J. Optimization of Bagasse, Nutrients and Initial Moisture Ratios on the Yield of Penicillin in Solid-State Fermentation. World J. Microbiol. Biotechnol. 2001, 17, 751–756. [Google Scholar] [CrossRef]
- Domínguez, M.; Mejía, A.; Barrios-González, J. Respiration Studies of Penicillin Solid-State Fermentation. J. Biosci. Bioeng. 2000, 89, 409–413. [Google Scholar] [CrossRef] [PubMed]
- López-Calleja, A.C.; Cuadra, T.; Barrios-González, J.; Fierro, F.; Fernández, F.J. Solid-State and Submerged Fermentations Show Different Gene Expression Profiles in Cephalosporin C Production by Acremonium chrysogenum. Microb. Physiol. 2012, 22, 126–134. [Google Scholar] [CrossRef]
- Cuadra, T.; Fernández, F.J.; Tomasini, A.; Barrios-González, J. Influence of PH Regulation and Nutrient Content on Cephalosporin C Production in Solid-State Fermentation by Acremonium Chrysogenum C10. Lett. Appl. Microbiol. 2007, 46, 216–220. [Google Scholar] [CrossRef]
- El-Housseiny, G.S.; Ibrahim, A.A.; Yassien, M.A.; Aboshanab, K.M. Production and Statistical Optimization of Paromomycin by Streptomyces rimosus NRRL 2455 in Solid State Fermentation. BMC Microbiol. 2021, 21, 34. [Google Scholar] [CrossRef]
- Machado, I.; Teixeira, J.A.; Rodríguez-Couto, S. Semi-Solid-State Fermentation: A Promising Alternative for Neomycin Production by the Actinomycete Streptomyces fradiae. J. Biotechnol. 2013, 165, 195–200. [Google Scholar] [CrossRef]
- Vastrad, B.M.; Neelagund, S.E.; Iiger, S.R.; Godbole, A.M.; Kulkarni, V. Improved Rifamycin B Production by Nocardia mediterranei MTCC 14 under Solid-State Fermentation through Process Optimization. Biochem. Res. Int. 2014, 2014, 1–13. [Google Scholar] [CrossRef]
- Lucas, E.M.; Machado, Y.; Ferreira, A.A.; Dolabella, L.M.; Takahashi, J.A. Improved Production of Pharmacologically-Active Sclerotiorin by Penicillium Sclerotiorum. 2010, Volume 9. Available online: http://www.tjpr.org (accessed on 15 June 2023).
- Saykhedkar, S.S.; Singhal, R.S. Solid-State Fermentation for Production of Griseofulvin on Rice Bran Using Penicillium griseofulvum. Biotechnol. Prog. 2004, 20, 1280–1284. [Google Scholar] [CrossRef]
- Zeng, X.; Miao, W.; Zeng, H.; Zhao, K.; Zhou, Y.; Zhang, J.; Zhao, Q.; Tursun, D.; Xu, D.; Li, F. Production of Natamycin by Streptomyces gilvosporeus Z28 through Solid-State Fermentation Using Agro-Industrial Residues. Bioresour. Technol. 2019, 273, 377–385. [Google Scholar] [CrossRef]
- Baños, J.G.; Tomasini, A.; Szakács, G.; Barrios-González, J. High Lovastatin Production by Aspergillus terreus in Solid-State Fermentation on Polyurethane Foam: An Artificial Inert Support. J. Biosci. Bioeng. 2009, 108, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Ávila, N.; Tarragó-Castellanos, M.R.; Barrios-González, J. Environmental Cues That Induce the Physiology of Solid Medium: A Study on Lovastatin Production by Aspergillus terreus. J. Appl. Microbiol. 2017, 122, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, A.; Uribe-Carvajal, S.; Cabrera-Orefice, A.; Barrios-González, J. Key Role of Alternative Oxidase in Lovastatin Solid-State Fermentation. Appl. Microbiol. Biotechnol. 2017, 101, 7347–7356. [Google Scholar] [CrossRef] [PubMed]
- Shaligram, N.S.; Singh, S.K.; Singhal, R.S.; Pandey, A.; Szakacs, G. Compactin Production Studies Using Penicillium brevicompactum Under Solid-State Fermentation Conditions. Appl. Biochem. Biotechnol. 2009, 159, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.-B.; Xing, H.-B.; Jiang, B.-J.; Chen, L.; Xu, G.-R.; Jiang, Y.; Zhang, D.-Y. Using Millet as Substrate for Efficient Production of Monacolin K by Solid-State Fermentation of Monascus ruber. J. Biosci. Bioeng. 2018, 125, 333–338. [Google Scholar] [CrossRef]
- Liu, X.; Sun, A.; Li, Q.; Du, Y.; Zhao, T. A Systematic Study of the Production of Monacolin K by Solid State Fermentation of Monascus ruber. AMB Express 2022, 12, 29. [Google Scholar] [CrossRef]
- Zouari, R.; Ellouze-Chaabouni, S.; Ghribi-Aydi, D. Optimization of Bacillus subtilis SPB1 Biosurfactant Production Under Solid-State Fermentation Using By-Products of a Traditional Olive Mill Factory. Achiev. Life Sci. 2014, 8, 162–169. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, F.; Wei, Z.; Ran, W.; Shen, Q. The Usage of Rice Straw as a Major Substrate for the Production of Surfactin by Bacillus amyloliquefaciens XZ-173 in Solid-State Fermentation. J. Env. Environ. Manag. 2013, 127, 96–102. [Google Scholar] [CrossRef]
- Piedrahíta-Aguirre, C.A.; Bastos, R.G.; Carvalho, A.L.; Monte Alegre, R. The Influence of Process Parameters in Production of Lipopeptide Iturin A Using Aerated Packed Bed Bioreactors in Solid-State Fermentation. Bioprocess. Biosyst. Eng. 2014, 37, 1569–1576. [Google Scholar] [CrossRef]
- Gong, Z.; He, Q.; Che, C.; Liu, J.; Yang, G. Optimization and Scale-up of the Production of Rhamnolipid by Pseudomonas aeruginosa in Solid-State Fermentation Using High-Density Polyurethane Foam as an Inert Support. Bioprocess. Biosyst. Eng. 2020, 43, 385–392. [Google Scholar] [CrossRef]
- Dabaghi, S.; Ataei, S.A.; Taheri, A. Production of Rhamnolipid Biosurfactants in Solid-State Fermentation: Process Optimization and Characterization Studies. BMC Biotechnol. 2023, 23, 2. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Peñalver, P.; Castillejos, M.; Koh, A.; Gross, R.; Sánchez, A.; Font, X.; Gea, T. Production and Characterization of Sophorolipids from Stearic Acid by Solid-State Fermentation, a Cleaner Alternative to Chemical Surfactants. J. Clean. Prod. 2018, 172, 2735–2747. [Google Scholar] [CrossRef]
- Rodríguez, A.; Gea, T.; Font, X. Sophorolipids Production from Oil Cake by Solid-State Fermentation. Inventory for Economic and Environmental Assessment. Front. Chem. Eng. 2021, 3, 2752. [Google Scholar] [CrossRef]
- Mehmood, T.; Saleem, F.; Javed, S.; Nawaz, S.; Sultan, A.; Safdar, A.; Ullah, A.; Waseem, R.; Saeed, S.; Abbas, M.; et al. Biotransformation of Agricultural By-Products into Biovanillin through Solid-State Fermentation (SSF) and Optimization of Different Parameters Using Response Surface Methodology (RSM). Fermentation 2022, 8, 206. [Google Scholar] [CrossRef]
- Mehmood, T.; Ahmed, S.; Waseem, R.; Saeed, S.; Ahmed, W.; Irfan, M.; Ullah, A. Valorization of Fruit Peels into Biovanillin and Statistical Optimization of Process Using Enterobacter hormaechei through Solid-State Fermentation. Fermentation 2022, 8, 40. [Google Scholar] [CrossRef]
- Mehmood, T.; Saeed, S.; Hussain, N.; Waseem, R. Biotransformation of Wheat Straw into Biovanillin by Solid-State Fermentation and Optimization of Conditions Parameters through Response Surface Methodology. Biomass Convers. Biorefin 2022, 19, 10. [Google Scholar] [CrossRef]
- Saeed, S.; Aslam, S.; Mehmood, T.; Naseer, R.; Nawaz, S.; Mujahid, H.; Firyal, S.; Anjum, A.A.; Sultan, A. Production of Gallic Acid Under Solid-State Fermentation by Utilizing Waste from Food Processing Industries. Waste Biomass Valorization 2021, 12, 155–163. [Google Scholar] [CrossRef]
- Wu, C.-Y.; Liang, C.-H.; Ou, C.-H.; Liang, Z.-C. Zinc Ion Addition to Grain Media Enhanced Hispidin Production during Solid-State Fermentation of Phellinus linteus. J. Taiwan. Inst. Chem. Eng. 2021, 121, 101–107. [Google Scholar] [CrossRef]
- Patel, G.; Patil, M.D.; Soni, S.; Chisti, Y.; Banerjee, U.C. Production of Mycophenolic Acid by Penicillium brevicompactum Using Solid State Fermentation. Appl. Biochem. Biotechnol. 2017, 182, 97–109. [Google Scholar] [CrossRef]
- Survase, S.A. A Novel Medium for the Enhanced Production of Cyclosporin A ByTolypocladium inflatum MTCC 557 Using Solid State Fermentation. J. Microbiol. Biotechnol. 2009, 19, 462–467. [Google Scholar] [CrossRef]
- Gelmi, C.; Pérez-Correa, R.; Agosin, E. Modelling Gibberella fujikuroi Growth and GA3 Production in Solid-State Fermentation. Process Biochem. 2002, 37, 1033–1040. [Google Scholar] [CrossRef]
- Doi, Y.; Wakana, D.; Takeda, H.; Tanaka, E.; Hosoe, T. Production of Ergot Alkaloids by the Japanese Isolate Claviceps purpurea Var. Agropyri on Rice Medium. Adv. Microbiol. 2022, 12, 254–269. [Google Scholar] [CrossRef]
- Dufossé, L.; Galaup, P.; Yaron, A.; Arad, S.M.; Blanc, P.; Chidambara Murthy, K.N.; Ravishankar, G.A. Microorganisms and Microalgae as Sources of Pigments for Food Use: A Scientific Oddity or an Industrial Reality? Trends Food Sci. Technol. 2005, 16, 389–406. [Google Scholar] [CrossRef]
- Arumugam, G.K.; Selvaraj, V.; Gopal, D.; Ramalingam, K. Solid-State Fermentation of Agricultural Residues for the Production of Antibiotics. In Biotransformation of Waste Biomass into High Value Biochemicals; Springer: New York, NY, USA, 2014; pp. 139–162. [Google Scholar] [CrossRef]
- Barrios-González, J.; Pérez-Sánchez, A.; Bibián, M.E. New Knowledge about the Biosynthesis of Lovastatin and Its Production by Fermentation of Aspergillus terreus. Appl. Microbiol. Biotechnol. 2020, 104, 8979–8998. [Google Scholar] [CrossRef]
- Suryanarayan, S. Current Industrial Practice in Solid State Fermentations for Secondary Metabolite Production: The Biocon India Experience. Biochem. Eng. J. 2003, 13, 189–195. [Google Scholar] [CrossRef]
- Banat, I.M.; Carboué, Q.; Saucedo-Castañeda, G.; de Jesús Cázares-Marinero, J. Biosurfactants: The Green Generation of Speciality Chemicals and Potential Production Using Solid-State Fermentation (SSF) Technology. Bioresour. Technol. 2021, 320, 124222. [Google Scholar] [CrossRef] [PubMed]
- Pascoalino, L.A.; Pires, T.C.S.P.; Taofiq, O.; Ferreira, I.C.F.R.; Barros, L.; Reis, F.S. Biochemistry of Secondary Metabolism of Fungi. In Natural Secondary Metabolites; Springer International Publishing: Cham, Switzerland, 2023; pp. 437–474. [Google Scholar] [CrossRef]
- Saltveit, M.E. Synthesis and Metabolism of Phenolic Compounds. In Fruit and Vegetable Phytochemicals; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 115–124. [Google Scholar] [CrossRef]
- Zurbriggen, A.; Kirst, H.; Melis, A. Isoprene Production Via the Mevalonic Acid Pathway in Escherichia coli (Bacteria). Bioenergy Res. 2012, 5, 814–828. [Google Scholar] [CrossRef]
- Francenia Santos-Sánchez, N.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C. Shikimic Acid Pathway in Biosynthesis of Phenolic Compounds. In Plant Physiological Aspects of Phenolic Compounds; IntechOpen: Rijeka, Croatia, 2019. [Google Scholar] [CrossRef]
- Sheng, Q.; Yi, L.; Zhong, B.; Wu, X.; Liu, L.; Zhang, B. Shikimic Acid Biosynthesis in Microorganisms: Current Status and Future Direction. Biotechnol. Adv. 2023, 62, 108073. [Google Scholar] [CrossRef]
- Emiliani, G.; Fondi, M.; Fani, R.; Gribaldo, S. A Horizontal Gene Transfer at the Origin of Phenylpropanoid Metabolism: A Key Adaptation of Plants to Land. Biol. Direct 2009, 4, 7. [Google Scholar] [CrossRef]
- Wu, S.; Chen, W.; Lu, S.; Zhang, H.; Yin, L. Metabolic Engineering of Shikimic Acid Biosynthesis Pathway for the Production of Shikimic Acid and Its Branched Products in Microorganisms: Advances and Prospects. Molecules 2022, 27, 4779. [Google Scholar] [CrossRef]
- Ghosh, S.; Chisti, Y.; Banerjee, U.C. Production of Shikimic Acid. Biotechnol. Adv. 2012, 30, 1425–1431. [Google Scholar] [CrossRef] [PubMed]
- Dhaniaputri, R.; Suwono, H.; Amin, M.; Lukiati, B. Introduction to Plant Metabolism, Secondary Metabolites Biosynthetic Pathway, and In-Silico Molecular Docking for Determination of Plant Medicinal Compounds: An Overview. In Proceedings of the 7th International Conference on Biological Science (ICBS 2021), Online, 14-15 October 2021. [Google Scholar] [CrossRef]
- Matulja, D.; Vranješević, F.; Kolympadi Markovic, M.; Pavelić, S.K.; Marković, D. Anticancer Activities of Marine-Derived Phenolic Compounds and Their Derivatives. Molecules 2022, 27, 1449. [Google Scholar] [CrossRef]
- Diner, B.A.; Fan, J.; Scotcher, M.C.; Wells, D.H.; Whited, G.M. Synthesis of Heterologous Mevalonic Acid Pathway Enzymes in Clostridium ljungdahlii for the Conversion of Fructose and of Syngas to Mevalonate and Isoprene. Appl. Env. Environ. Microbiol. 2018, 84, e01723-17. [Google Scholar] [CrossRef] [PubMed]
- Boronat, A.; Rodríguez-Concepción, M. Terpenoid Biosynthesis in Prokaryotes. In Biotechnology of Isoprenoids; Springer: Cham, Switzerland, 2014; pp. 3–18. [Google Scholar] [CrossRef]
- Karlic, H.; Varga, F. Mevalonate Pathway. In Encyclopedia of Cancer; Boffetta, P., Hainaut, P., Eds.; Academic Press: Oxford, UK, 2019; pp. 445–447. [Google Scholar]
- Zeng, L.; Dehesh, K. The Eukaryotic MEP-Pathway Genes Are Evolutionarily Conserved and Originated from Chlaymidia and Cyanobacteria. BMC Genom. 2021, 22, 137. [Google Scholar] [CrossRef]
- Zhao, L.; Chang, W.; Xiao, Y.; Liu, H.; Liu, P. Methylerythritol Phosphate Pathway of Isoprenoid Biosynthesis. Annu. Rev. Biochem. 2013, 82, 497–530. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Müller, R. Possibility of Bacterial Recruitment of Plant Genes Associated with the Biosynthesis of Secondary Metabolites. Plant Physiol. 2003, 132, 1153–1161. [Google Scholar] [CrossRef]
- Hu, M.; Chen, S.; Ni, Y.; Wei, W.; Mao, W.; Ge, M.; Qian, X. CRISPR/Cas9-Mediated Genome Editing in Vancomycin-Producing Strain Amycolatopsis keratiniphila. Front. Bioeng. Biotechnol. 2023, 11, 1141176. [Google Scholar] [CrossRef]
- Qian, H.; Wei, W.; Chen, X.-A.; Mo, X.-T.; Ge, M.; Zhao, Q.-W.; Li, Y.-Q. Strategy for Producing the High-Quality Glycopeptide Antibiotic A82846B in Amycolatopsis Orientalis Based on the CRISPR-Cas12a System. ACS Synth. Biol. 2021, 10, 3009–3016. [Google Scholar] [CrossRef]
- Vásquez-Bonilla, J.N.; Barranco-Florido, J.E.; Ponce-Alquicira, E.; Rincón-Guevara, M.A.; Loera, O. Improvement of Beauvericin Production by Fusarium Oxysporum AB2 under Solid-State Fermentation Using an Optimised Liquid Medium and Co-Cultures. Mycotoxin Res. 2022, 38, 175–183. [Google Scholar] [CrossRef]
- Tiwari, P.; Dufossé, L. Focus and Insights into the Synthetic Biology-Mediated Chassis of Economically Important Fungi for the Production of High-Value Metabolites. Microorganisms 2023, 11, 1141. [Google Scholar] [CrossRef]
- Pickens, L.B.; Tang, Y.; Chooi, Y.-H. Metabolic Engineering for the Production of Natural Products. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 211–236. [Google Scholar] [CrossRef]
- Srivastava, N.; Srivastava, M.; Ramteke, P.W.; Mishra, P.K. Solid-State Fermentation Strategy for Microbial Metabolites Production: An Overview. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 345–354. [Google Scholar] [CrossRef]
- Félix, M.K.C.; Leite, E.R.T.; Dutra, L.R.; Ribeiro, M.A.; Moura, W.S.; Ferreira, T.P.S.; Santos, G.R.; Chapla, V.M.; Vieira, W.A.S.; Câmara, M.P.S.; et al. Secondary Metabolites Produced by Solid-State Fermentation of a Neotropical Aspergillus flavus Strain Confer Anti-Mosquito Activity with Long Shelf-Life. Ind. Crops Prod. 2022, 180, 114743. [Google Scholar] [CrossRef]
- Shen, Z.; Zheng, P.; Li, R.; Sun, X.; Chen, P.; Wu, D. High Production of Jasmonic Acid by Lasiodiplodia iranensis Using Solid-State Fermentation: Optimization and Understanding. Biotechnol. J. 2022, 17, 2100550. [Google Scholar] [CrossRef] [PubMed]
- Kaaniche, F.; Hamed, A.; Elleuch, L.; Chakchouk-Mtibaa, A.; Smaoui, S.; Karray-Rebai, I.; Koubaa, I.; Arcile, G.; Allouche, N.; Mellouli, L. Purification and Characterization of Seven Bioactive Compounds from the Newly Isolated Streptomyces cavourensis TN638 Strain via Solid-State Fermentation. Microb. Pathog. 2020, 142, 104106. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.; Dong, J. Multi-Level Optimization and Strategies in Microbial Biotransformation of Nature Products. Molecules 2023, 28, 2619. [Google Scholar] [CrossRef]
- Li, S.; Li, Z.; Pang, S.; Xiang, W.; Wang, W. Coordinating Precursor Supply for Pharmaceutical Polyketide Production in Streptomyces. Curr. Opin. Biotechnol. 2021, 69, 26–34. [Google Scholar] [CrossRef]
- Rollini, M.; Manzoni, M. Influence of Medium Design on Lovastatin and Mevastatin Production By Aspergillus terreus Strains. Ann. Microbiol. 2006, 56, 47–51. [Google Scholar] [CrossRef]
- Ábrego-García, A.; Poggi-Varaldo, H.M.; Ponce-Noyola, M.T.; Calva-Calva, G.; Galíndez-Mayer, C.J.J.; Medina-Mendoza, G.G.; Rinderknecht-Seijas, N.F. Bioprocessing of Two Crop Residues for Animal Feeding into a High-Yield Lovastatin Feed Supplement. Animals 2022, 12, 2697. [Google Scholar] [CrossRef]
- Rentería-Martínez, O.; Páez-Lerma, J.B.; Rojas-Contreras, J.A.; López-Miranda, J.; Martell-Nevárez, M.A.; Soto-Cruz, N.O. Enhancing Isoamyl Acetate Biosynthesis by Pichia fermentans. Rev. Mex. Ing. Quim. 2021, 20, 621–633. [Google Scholar] [CrossRef]
- Mantle, P.; Roberts, A.; Beaumont, C. Notoamide R: A Prominent Diketopiperazine Fermentation Metabolite amongst Others of Aspergillus ochraceus in the Absence of Ochratoxins. Molecules 2023, 28, 3518. [Google Scholar] [CrossRef]
- Li, C.; Hazzard, C.; Florova, G.; Reynolds, K.A. High Titer Production of Tetracenomycins by Heterologous Expression of the Pathway in a Streptomyces cinnamonensis Industrial Monensin Producer Strain. Metab. Eng. 2009, 11, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Leonard, E.; Ajikumar, P.K.; Thayer, K.; Xiao, W.-H.; Mo, J.D.; Tidor, B.; Stephanopoulos, G.; Prather, K.L.J. Combining Metabolic and Protein Engineering of a Terpenoid Biosynthetic Pathway for Overproduction and Selectivity Control. Proc. Natl. Acad. Sci. USA 2010, 107, 13654–13659. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, B.A.; Khosla, C. Biosynthesis of Polyketides in Heterologous Hosts. Microbiol. Mol. Biol. Rev. 2001, 65, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; You, Y.; Chen, Z.; Li, J.; Liu, G.; Wen, Y. Avermectin B1a Production in Streptomyces avermitilis Is Enhanced by Engineering AveC and Precursor Supply Genes. Appl. Microbiol. Biotechnol. 2022, 106, 2191–2205. [Google Scholar] [CrossRef]
- Hao, F.; Wu, Q.; Xu, Y. Precursor Supply Strategy for Tetramethylpyrazine Production by Bacillus subtilis on Solid-State Fermentation of Wheat Bran. Appl. Biochem. Biotechnol. 2013, 169, 1346–1352. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Si, G.; Rao, Z.; Li, J.; Zhang, X.; Mei, J.; Wang, J.; Ye, M.; Zhou, P. High Yield of Tetramethylpyrazine in Functional Fuqu Using Bacillus amyloliquefaciens. Food Biosci. 2019, 31, 100435. [Google Scholar] [CrossRef]
- Gao, H.; Lu, C.; Wang, H.; Wang, L.; Yang, Y.; Jiang, T.; Li, S.; Xu, D.; Wu, L. Production Exopolysaccharide from Kosakonia cowanii LT-1 through Solid-State Fermentation and Its Application as a Plant Growth Promoter. Int. J. Biol. Macromol. 2020, 150, 955–964. [Google Scholar] [CrossRef]
- Lindsay, M.A.; Granucci, N.; Greenwood, D.R.; Villas-Boas, S.G. Identification of New Natural Sources of Flavour and Aroma Metabolites from Solid-State Fermentation of Agro-Industrial By-Products. Metabolites 2022, 12, 157. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Alli, I.; Ereifej, K.; Alhamad, M.; Al-Tawaha, A.R.; Rababah, T. Optimisation, Characterisation and Quantification of Phenolic Compounds in Olive Cake. Food Chem. 2010, 123, 117–122. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, D.H.; Kang, H.J.; Shin, M.; Yang, S.-Y.; Yang, J.; Jung, Y.H. Enhanced Production of γ-Aminobutyric Acid (GABA) Using Lactobacillus plantarum EJ2014 with Simple Medium Composition. LWT 2021, 137, 110443. [Google Scholar] [CrossRef]
- Sun, Y.; Mehmood, A.; Battino, M.; Xiao, J.; Chen, X. Enrichment of Gamma-Aminobutyric Acid in Foods: From Conventional Methods to Innovative Technologies. Food Res. Int. 2022, 162, 111801. [Google Scholar] [CrossRef]
- Phukon, L.C.; Singh, S.P.; Pandey, A.; Rai, A.K. Microbial Bioprocesses for Production of Nutraceuticals and Functional Foods. In Current Developments in Biotechnology and Bioengineering; Rai, A.K., Singh, S.P., Pandey, A., Larroche, C., Soccol, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 1–29. [Google Scholar] [CrossRef]
- Hou, D.; Tang, J.; Feng, Q.; Niu, Z.; Shen, Q.; Wang, L.; Zhou, S. Gamma-Aminobutyric Acid (GABA): A Comprehensive Review of Dietary Sources, Enrichment Technologies, Processing Effects, Health Benefits, and Its Applications. Crit. Rev. Food Sci. Nutr. 2023, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Gao, F.; Zhang, X.; Wang, O.; Wu, W.; Zhu, S.; Zhang, D.; Zhou, F.; Ji, B. Evaluation of γ- Aminobutyric Acid, Phytate and Antioxidant Activity of Tempeh-like Fermented Oats (Avena sativa L.) Prepared with Different Filamentous Fungi. J. Food Sci. Technol. 2014, 51, 2544–2551. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.; Regmi, O.; Hasan, M.; Panda, B.P. Response Surface Modeling for the Enrichment of Gamma-Aminobutyric Acid with a Minimum Content of Citrinin in Monascus-Fermented Rice. eFood 2020, 1, 181–187. [Google Scholar] [CrossRef]
- Chen, C.Y.; Zhang, R.; Zhang, L.J.; Hu, Z.Y.; Wang, S.P.; Mei, X.; Mi, W.; Zhang, J.Y. Biotransformation and Bioaccessibility of Active Ingredients from Radix Astragali by Poria cocos during Solid-State Fermentation and in Vitro Digestion and Antioxidant Activity Evaluation. Sci. Rep. 2023, 13, 6888. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hou, X.; Xu, R.; Liu, C.; Tu, M. Research Review on the Pharmacological Effects of Astragaloside IV. Fundam. Clin. Pharmacol. 2017, 36, 17–36. [Google Scholar] [CrossRef]
- Dikshit, R.; Tallapragada, P. Screening and Optimization of γ-Aminobutyric Acid Production from Monascus sanguineus under Solid-State Fermentation. Front. Life Sci. 2015, 8, 172–181. [Google Scholar] [CrossRef]
- Zhang, L.; Yue, Y.; Wang, X.; Dai, W.; Piao, C.; Yu, H. Optimization of Fermentation for γ-Aminobutyric Acid (GABA) Production by Yeast Kluyveromyces marxianus C21 in Okara (Soybean Residue). Bioprocess. Biosyst. Eng. 2022, 45, 1111–1123. [Google Scholar] [CrossRef]
- Martínez-Avila, O.; Muñoz-Torrero, P.; Sánchez, A.; Font, X.; Barrena, R. Valorization of Agro-Industrial Wastes by Producing 2-Phenylethanol via Solid-State Fermentation: Influence of Substrate Selection on the Process. Waste Manag. 2021, 121, 403–411. [Google Scholar] [CrossRef]
- Roy, P.; Gahlawat, V.K.; Saravanan, C.; Singh, B.P. Enhancing Bioflavor Production by Solid-State Fermentation Using Kluyveromyces marxianus and l-Phenylalanine. J. Basic. Microbiol. 2023, 63, 75–91. [Google Scholar] [CrossRef]
- Peng, L.; Ai-lati, A.; Liu, S.; Ji, Z.; Mao, J.; Che, X. Effects of Chinese Medicines on Monacolin K Production and Related Genes Transcription of Monascus ruber in Red Mold Rice Fermentation. Food Sci. Nutr. 2020, 8, 2134–2142. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Liu, Y.; Huan, C.C.; Xu, L.; Ji, G.; Yan, Z. Comparison of Poly-γ-Glutamic Acid Production between Sterilized and Non-Sterilized Solid-State Fermentation Using Agricultural Waste as Substrates. J. Clean. Prod. 2020, 255, 120248. [Google Scholar] [CrossRef]
- Oreb, M. Construction of Artificial Membrane Transport Metabolons—An Emerging Strategy in Metabolic Engineering. FEMS Microbiol. Lett. 2020, 367, fnaa027. [Google Scholar] [CrossRef]
- Ge, X.; Vasco-Correa, J.; Li, Y. Solid-State Fermentation Bioreactors and Fundamentals. In Current Developments in Biotechnology and Bioengineering; Larroche, C., Sanromán, M.Á., Du, G., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 381–402. [Google Scholar] [CrossRef]
- Chen, H. Principles of Solid-State Fermentation Engineering and Its Scale-Up. In Modern Solid State Fermentation; Chen, H., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 75–139. [Google Scholar] [CrossRef]
- Tang, Q.; Chen, X.; Huang, J.; Zhang, S.; Qin, H.; Dong, Y.; Wang, C.; Wang, X.; Wu, C.; Jin, Y.; et al. Mechanism of Enhancing Pyrazines in Daqu via Inoculating Bacillus licheniformis with Strains Specificity. Foods 2023, 12, 304. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Metabolite and Its Potential Application | Microorganism | Solid Support | Yield (mg/g SS) | Reference |
|---|---|---|---|---|
| Pigments | ||||
| Monascin, ankaflavin, rubropunctatin, monascorubrin, rubropunctamin, monascorubramine, etc. | Monascus sp. | Rice | 49.65 * | [12,13] |
| Antibiotics | ||||
| Penicillin | Penicillium chrysogenum | Sugarcane bagasse | 7–8 | [14,15] |
| Cephalosporin C | Acremonium chrysogenum | Sugarcane bagasse | 3.2 | [16,17] |
| Paromomycin | Streptomyces rimosus | Corn bran | 2.2 | [18] |
| Neomycin | Streptomyces fradiae | Nylon sponge | n.d. | [19] |
| Rifamycin B | Nocardia mediterranei | Sunflower oil cake | 9.87 | [20] |
| Antifungal | ||||
| Sclerotiorin | Penicillium sclerotiorum | Rice | n.d. | [21] |
| Griseofulvin | Penicillium griseofulvum | Rice bran | 9–10 | [22] |
| Natamycin | Streptomyces gilvosporeus | Wheat bran, rapeseed cake, and rice hull | 9.62 | [23] |
| Statins | ||||
| Lovastatin | Aspergillus terreus | Glucose and lactose | 19.95–25 | [24,25,26] |
| Compactin (mevastatin) | Penicillium brevicompactum | Soybean meal | 1.406 | [27] |
| Monacolin K | Monascus ruber | Millet | 19.81 | [28] |
| Rice and bran | 14.53 | [29] | ||
| Biosurfactants | ||||
| Surfactin | Bacillus subtilis | Olive cake flour | 30.67 | [30] |
| Bacillus amyloliquefaciens | Soybean flour | 15.03 | [31] | |
| Iturin | Bacillus subtilis | Defatted soybean meal, wheat bran and ricehusk | 5.58 | [32] |
| Rhamnolipids | Pseudomonas aeruginosa | Polyurethane foam | 39.8 ** | [33] |
| Soybean meal | 19.68 | [34] | ||
| Sophorolipids | Starmerella bombicola | Polyurethane foam | 211 *** | [35] |
| Wheat straw | 195 | [36] | ||
| Phenolics | ||||
| Vainillin | Enterobacter hormaechei | Sugarcane bagasse | 4.76 | [37] |
| Enterobacter hormaechei | Pomegranate peels | 0.462 | [38] | |
| Streptomyces sannanensis | Wheat straw | 2.74 | [39] | |
| Gallic acid | Aspergillus niger | Black plum seed | 14.5 | [40] |
| Hispidin | Phellinus linteus | Brown rice and pearl barley | 0.375 | [41] |
| Immunosuppressants | ||||
| Mycophenolic acid | Penicillium brevicompactum | Parmal rice | 4.5 | [42] |
| Cyclosporin A | Tolypocladium inflatum | Wheat bran flour and coconut oil cake | 6.48 | [43] |
| Phytohormones | ||||
| Gibberellic acid | Gibberella fujikuroi | Amberlite IRA-900 | n.d. | [44] |
| Alkaloids | ||||
| Ergotamine | Claviceps purpurea | Rice | ≈0.015 | [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méndez-Hernández, J.E.; Rodríguez-Durán, L.V.; Páez-Lerma, J.B.; Soto-Cruz, N.O. Strategies for Supplying Precursors to Enhance the Production of Secondary Metabolites in Solid-State Fermentation. Fermentation 2023, 9, 804. https://doi.org/10.3390/fermentation9090804
Méndez-Hernández JE, Rodríguez-Durán LV, Páez-Lerma JB, Soto-Cruz NO. Strategies for Supplying Precursors to Enhance the Production of Secondary Metabolites in Solid-State Fermentation. Fermentation. 2023; 9(9):804. https://doi.org/10.3390/fermentation9090804
Chicago/Turabian StyleMéndez-Hernández, Jazmín E., Luis V. Rodríguez-Durán, Jesús B. Páez-Lerma, and Nicolás O. Soto-Cruz. 2023. "Strategies for Supplying Precursors to Enhance the Production of Secondary Metabolites in Solid-State Fermentation" Fermentation 9, no. 9: 804. https://doi.org/10.3390/fermentation9090804
APA StyleMéndez-Hernández, J. E., Rodríguez-Durán, L. V., Páez-Lerma, J. B., & Soto-Cruz, N. O. (2023). Strategies for Supplying Precursors to Enhance the Production of Secondary Metabolites in Solid-State Fermentation. Fermentation, 9(9), 804. https://doi.org/10.3390/fermentation9090804