
Bioconversion Study of Olive Tree Biomass Hemicellulosic Hydrolysates by Candida guilliermondii at Different Scales for Ethanol and Xylitol Production

 , ,
, ,  , and
, and 
Abstract

1. Introduction
2. Materials and Methods
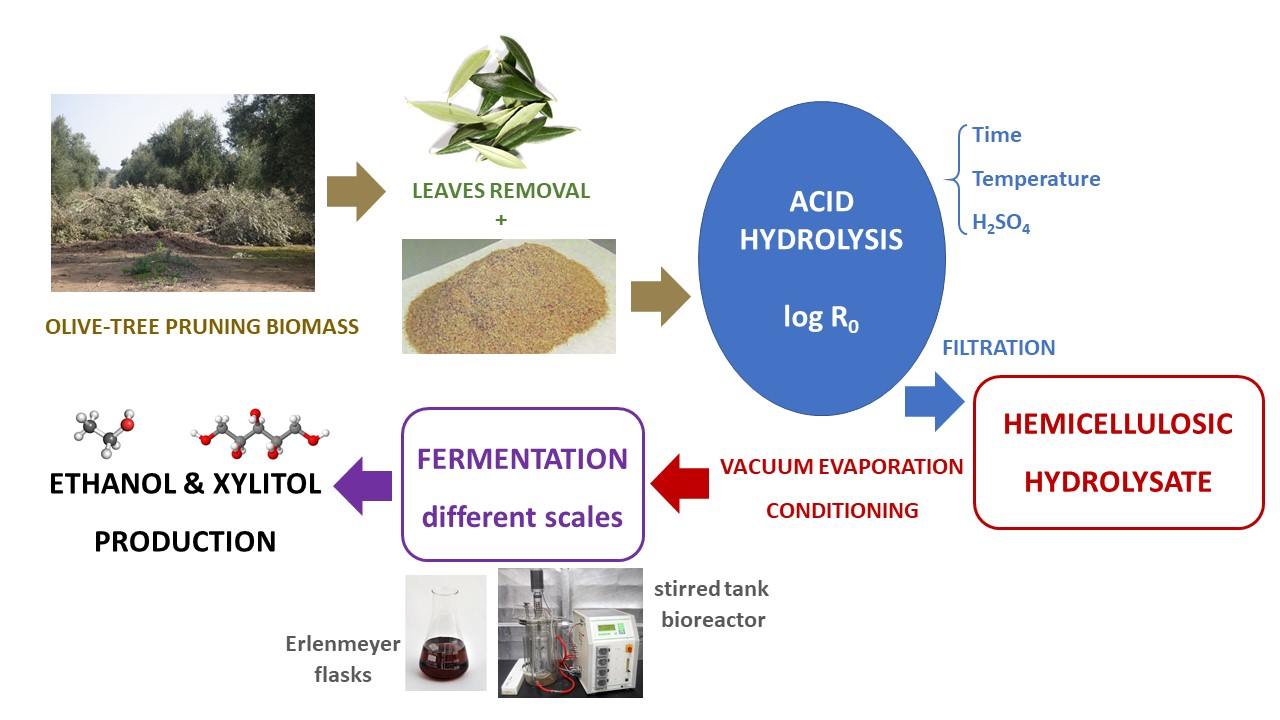
2.1. Raw Material
2.2. General Analytical Methods
2.3. Acid Hydrolysis
2.4. Microorganism and Inoculum Cultivation
2.5. Fermentation Conditions
2.6. Fermentative Parameters Calculation
3. Results and Discussion
3.1. Raw Material Composition
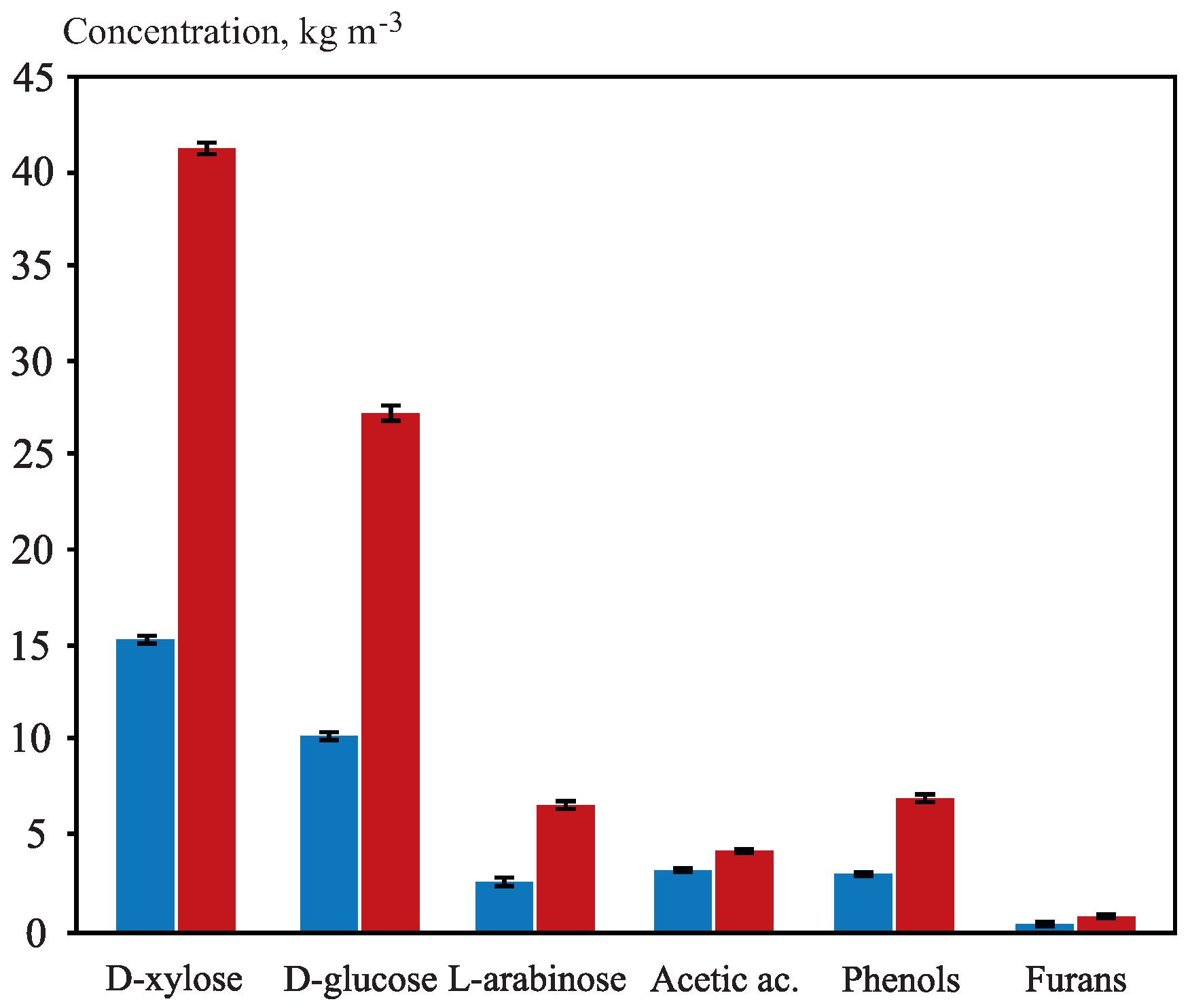
3.2. OTPB Hemicellulosic Hydrolysate
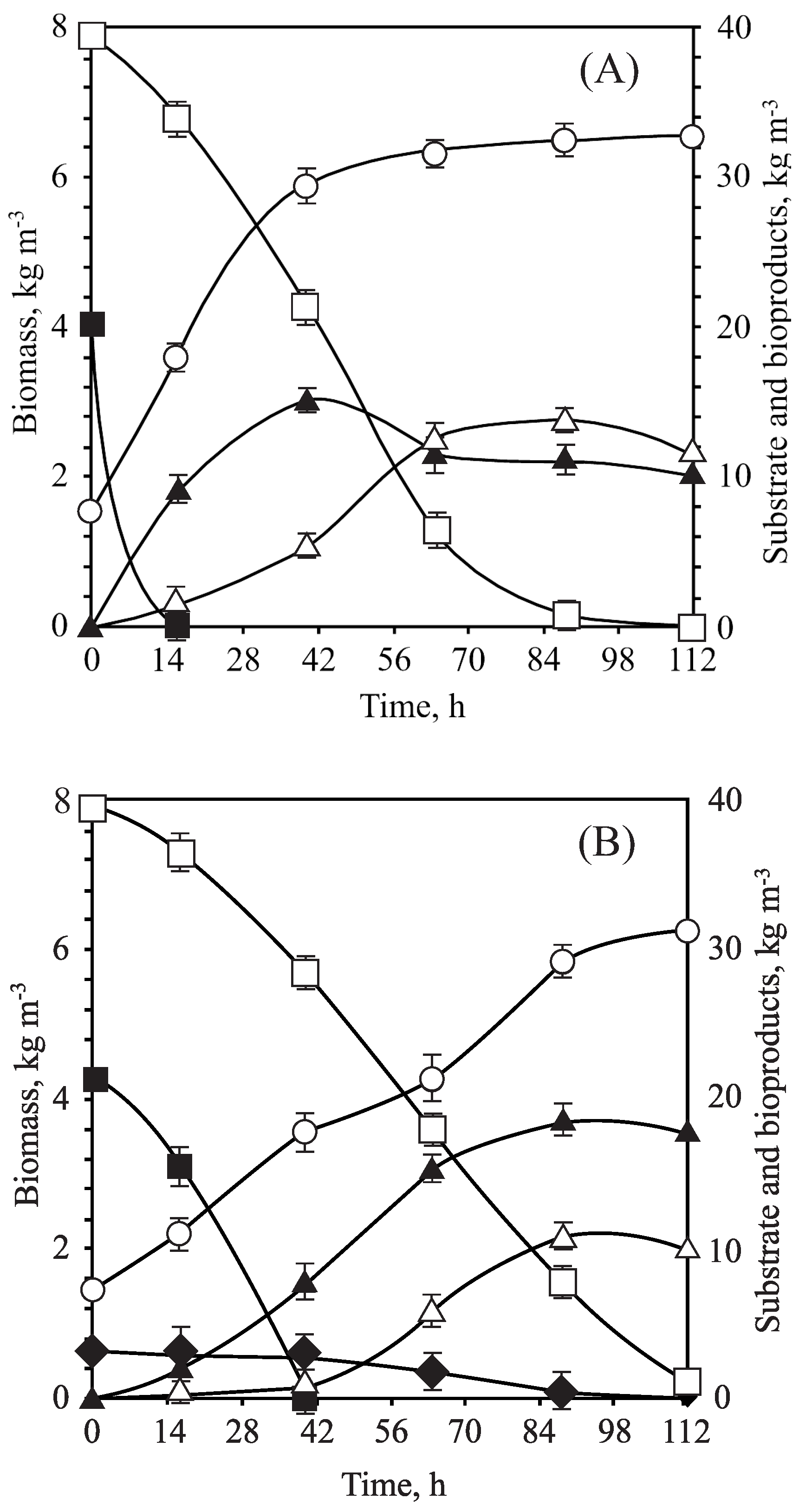
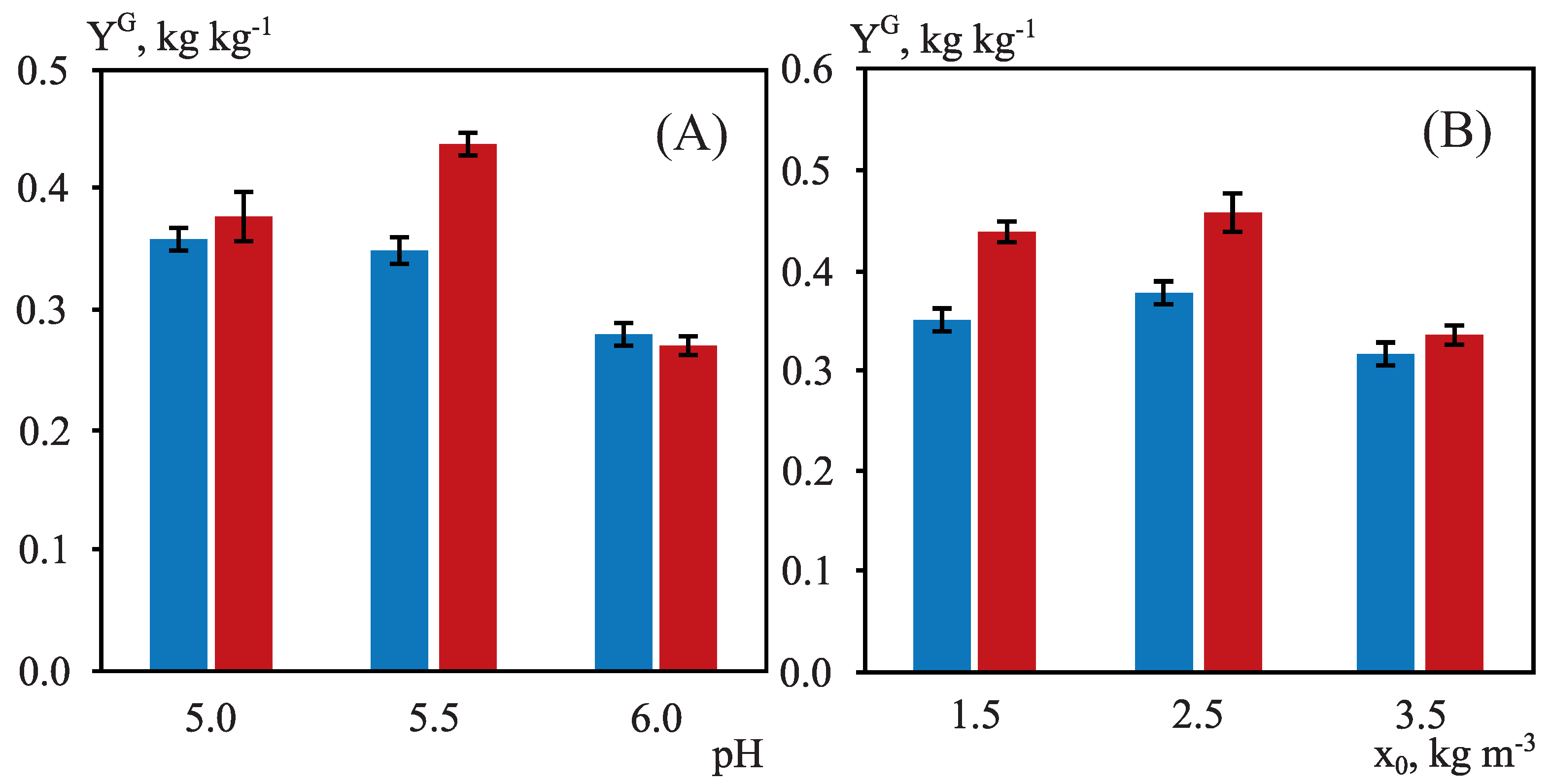
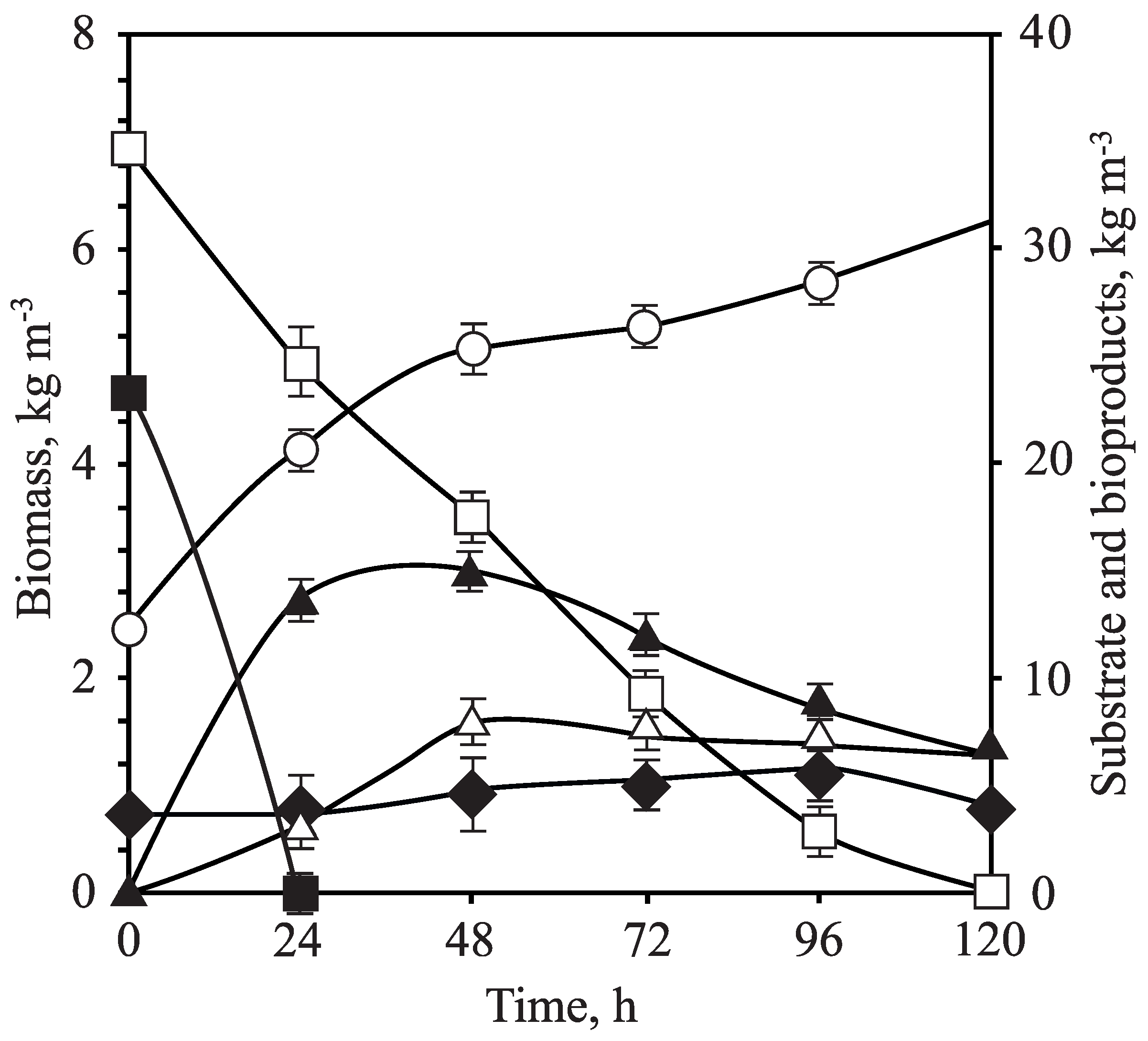
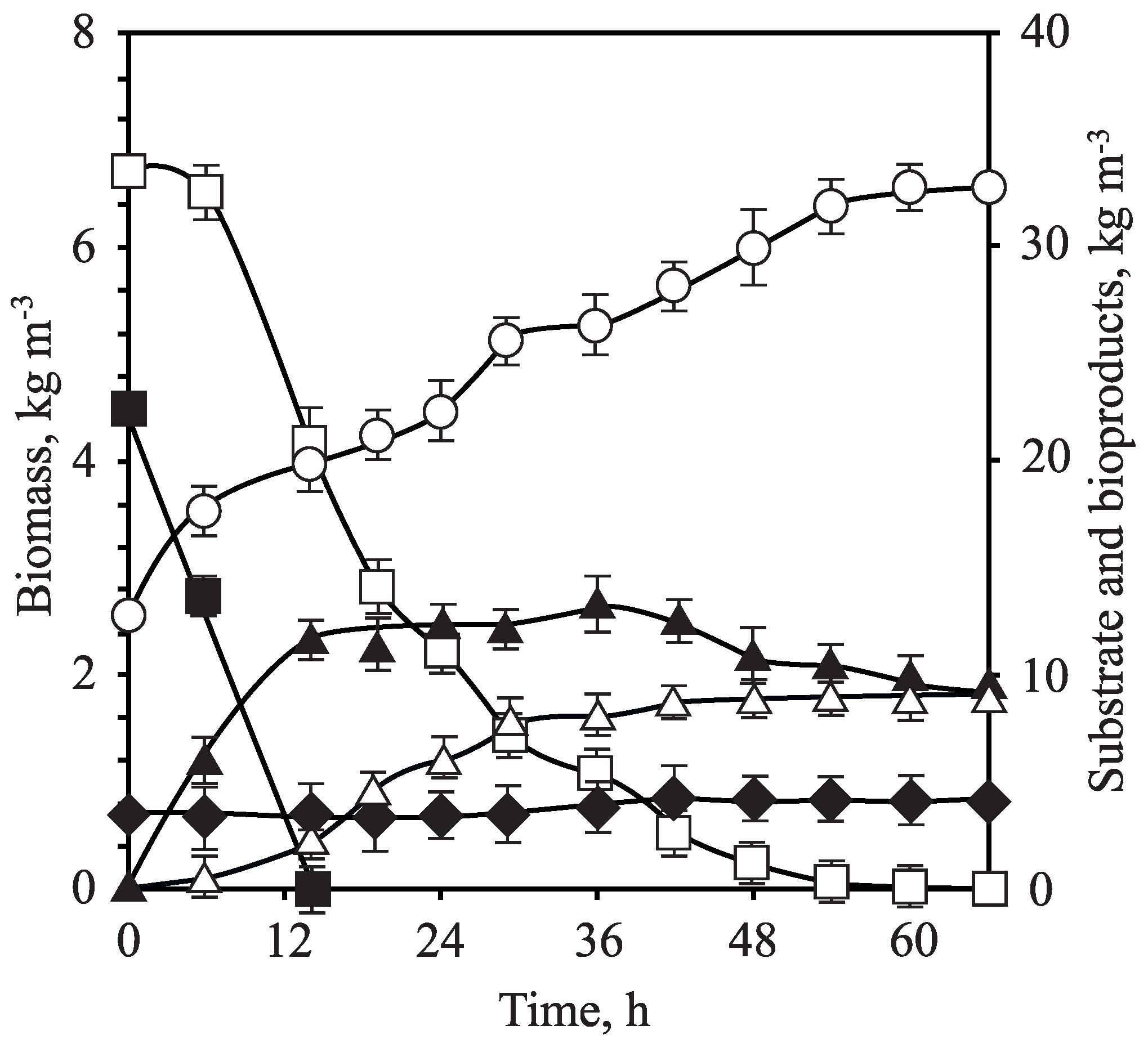
3.3. Fermentation of OTPB Concentrated Hemicellulosic Hydrolysate at Different Scales
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Absorbance at 600 nm | |
| Absorbance at 760 nm | |
| ASTM | American Society for Testing and Materials |
| b | Biomass volumetric productivity |
| HMF | 5-hydroxymethylfurfural |
| HPLC | High-performance liquid chromatography |
| Oxygen transfer volumetric coefficient | |
| log R | Combined severity factor |
| NREL | National Renewable Energy Laboratory |
| OTPB | Olive tree-pruning biomass |
| OTPB-H | OTPB concentrated hydrolysate without addition of nutrients |
| OTPB-LH | OTPB concentrated hydrolysate with addition of nutrients proposed by [36] at 50% |
| OTPB-RH | OTPB concentrated hydrolysate with addition of nutrients proposed by [37] |
| Ethanol volumetric productivity | |
| Specific rate of substrate consumption | |
| Xylitol volumetric productivity | |
| SM-L | Substrate constituted by the addition of nutrients proposed by [36] at 50% |
| SM-R | Substrate constituted by the addition of nutrients proposed by [37] |
| Total phenolic compounds concentration | |
| Biomass global yield | |
| Ethanol global yield | |
| Xylitol global yield | |
| Aeration |
References
- Domingues, J.; Pelletier, C.; Brunelle, T. Cost of ligno-cellulosic biomass production for bioenergy: A review in 45 countries. Biomass Bioenergy 2022, 165, 106583. [Google Scholar] [CrossRef]
- Hoang, A.T.; Nizetic, S.; Ong, H.C.; Chong, C.T.; Atabani, A.; Pham, V. Acid-based lignocellulosic biomass biorefinery for bioenergy production: Advantages, application constraints, and perspectives. J. Environ. Manag. 2021, 296, 113194. [Google Scholar] [CrossRef] [PubMed]
- Mateo, S.; Puentes, J.; Sánchez, S.; Moya, A. Oligosaccharides and monomeric carbohydrates production from olive tree pruning biomass. Carbohydr. Polym. 2013, 93, 416–423. [Google Scholar] [CrossRef]
- Wyman, C.E. Ethanol from lignocellulosic biomass: Technology, economics, and opportunities. Bioresour. Technol. 1994, 50, 3–15. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, Y.Y.; Torget, R.W. Cellulose hydrolysis under extremely low sulfuric acid and high-temperature conditions. In 22nd Symposium on Biotechnology for Fuels and Chemicals; Humana Press: New York, NY, USA, 2001; pp. 331–340. [Google Scholar]
- Mussatto, S.; Roberto, I. Kinetic behavior of Candida guilliermondii yeast during xylitol production from highly concentrated hydrolysate. Process Biochem. 2004, 39, 1433–1439. [Google Scholar] [CrossRef]
- Villarreal, M.; Prata, A.; Felipe, M.; Silva, J.A.E. Detoxification procedures of eucalyptus hemicellulose hydrolysate for xylitol production by Candida guilliermondii. Enzym. Microb. Technol. 2006, 40, 17–24. [Google Scholar] [CrossRef]
- Fonseca, B.G.; Mateo, S.; Moya, A.J.; Roberto, I.C. Biotreatment optimization of rice straw hydrolyzates for ethanolic fermentation with Scheffersomyces stipitis. Biomass Bioenergy 2018, 112, 19–28. [Google Scholar] [CrossRef]
- Mateo, S.; Roberto, I.C.; Sánchez, S.; Moya, A.J. Detoxification of hemicellulosic hydrolyzate from olive tree pruning residue. Ind. Crop. Prod. 2013, 49, 196–203. [Google Scholar] [CrossRef]
- Bari, I.D.; Liuzzi, F.; Villone, A.; Braccio, G. Hydrolysis of concentrated suspensions of steam pretreated Arundo donax. Appl. Energy 2013, 102, 179–189. [Google Scholar] [CrossRef]
- Fidio, N.D.; Liuzzi, F.; Mastrolitti, S.; Albergo, R.; Bari, I.D. Single Cell Oil Production from Undetoxified Arundo donax L. hydrolysate by Cutaneotrichosporon curvatus. J. Microbiol. Biotechnol. 2019, 29, 256–267. [Google Scholar] [CrossRef]
- Fidio, N.D.; Ragaglini, G.; Dragoni, F.; Antonetti, C.; Galletti, A.M.R. Integrated cascade biorefinery processes for the production of single cell oil by Lipomyces starkeyi from Arundo donax L. hydrolysates. Bioresour. Technol. 2021, 325, 124635. [Google Scholar] [CrossRef]
- Castillo-Ruiz, F.J.; Colmenero-Martinez, J.T.; Bayano-Tejero, S.; Gonzalez-Sanchez, E.J.; Lara, F.M.; Blanco-Roldán, G.L. Methodology for olive pruning windrow assessment using 3D time-of-flight camera. Agronomy 2021, 11, 1209. [Google Scholar] [CrossRef]
- Moya, A.J.; Bravo, V.; Mateo, S.; Sánchez, S. Fermentation of acid hydrolysates from olive-tree pruning debris by Pachysolen tannophilus. Bioprocess Biosyst. Eng. 2008, 31, 611–617. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Food and Agriculture Organization of the United Nations. Statistic Division. 2022. Available online: http://www.fao.org/faostat/en/#data/QC/ (accessed on 10 October 2022).
- Puentes, J.G.; Mateo, S.; Fonseca, B.G.; Roberto, I.C.; Sánchez, S.; Moya, A.J. Monomeric carbohydrates production from olive tree pruning biomass: Modeling of dilute acid hydrolysis. Bioresour. Technol. 2013, 149, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.; Mussatto, S.; Roberto, I. Study of xylitol production by Candida guilliermondii on a bench bioreactor. J. Food Eng. 2006, 75, 115–119. [Google Scholar] [CrossRef]
- Martínez, E.A.; Silva, S.S.; Felipe, M.G.A. Effect of the oxygen transfer coefficient on xylitol production from sugarcane bagasse hydrolysate by continuous stirred-tank reactor fermentation. In 21st Symposium on Biotechnology for Fuels and Chemicals; Humana Press: New York, NY, USA, 2000; pp. 633–641. [Google Scholar]
- Forsberg, C.; Dale, B.; Jones, D.; Hossain, T.; Morais, A.; Wendt, L. Replacing liquid fossil fuels and hydrocarbon chemical feedstocks with liquid biofuels from large-scale nuclear biorefineries. Appl. Energy 2021, 298, 117225. [Google Scholar] [CrossRef]
- Naik, S.; Goud, V.V.; Rout, P.K.; Dalai, A.K. Production of first and second generation biofuels: A comprehensive review. Renew. Sustain. Energy Rev. 2010, 14, 578–597. [Google Scholar] [CrossRef]
- Roberto, I.C.; Felipe, M.; Lacis, L.; Silva, S.; de Mancilha, I.M. Utilization of sugar cane bagasse hemicellulosic hydrolyzate by Candida guilliermondii for xylitol production. Bioresour. Technol. 1991, 36, 271–275. [Google Scholar] [CrossRef]
- Silva, C.; Roberto, I. Optimization of xylitol production by Candida guilliermondii FTI 20037 using response surface methodology. Process Biochem. 2001, 36, 1119–1124. [Google Scholar] [CrossRef]
- Cortez, D.; Roberto, I. Optimization of D-xylose to xylitol biotransformation by Candida guilliermondii cells permeabilized with Triton X-100. Biocatal. Biotransform. 2014, 32, 34–38. [Google Scholar] [CrossRef]
- Mateo, S.; Puentes, J.; Roberto, I.; Sánchez, S.; Moya, J. Optimization of acid hydrolysis of olive tree pruning residue. Fermentation with Candida guilliermondii. Biomass Bioenergy 2014, 69, 39–46. [Google Scholar] [CrossRef]
- Roberto, I.; Felipe, M.; de Mancilha, I.; Vitolo, M.; Sato, S.; da Silva, S.S. Xylitol production by Candida guilliermondii as an approach for the utilization of agroindustrial residues. Bioresour. Technol. 1995, 51, 255–257. [Google Scholar] [CrossRef]
- Prescott, L.M.; Harley, J.P.; Klein, D.A. Microbiology, 5th ed.; The McGraw-Hill Companies: New York, NY, USA, 2002. [Google Scholar]
- Soleimani, M.; Tabil, L.; Panigrahi, S. Bio-production of a polyalcohol (xylitol) from lignocellulosic resources: A review. In Proceedings of the Canadian Society for Bioengineering, Paper No. 06-106. CSBE-SCGAB Technical Conferences, Edmonton, AB, Canada, 16–19 July 2006; pp. 1–6. [Google Scholar]
- Pereira, R.; Mussatto, S.; Roberto, I. Inhibitory action of toxic compounds present in lignocellulosic hydrolysates on xylose to xylitol bioconversion by Candida guilliermondii. J. Ind. Microbiol. Biotechnol. 2010, 38, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Winkelhausen, E.; Amartey, S.; Kuzmanova, S. Xylitol production from D-Xylose at different oxygen transfer coefficients in a batch bioreactor. Eng. Life Sci. 2004, 4, 150–154. [Google Scholar] [CrossRef]
- Espeso, J.; Isaza, A.; Lee, J.Y.; Sörensen, P.M.; Jurado, P.; Avena-Bustillos, R.J.; Olaizola, M.; Arboleya, J. Olive Leaf Waste Management. Front. Sustain. Food Syst. 2021, 5, 660582. [Google Scholar] [CrossRef]
- Irick, T.J.; West, K.; Brownell, H.H.; Schwald, W.; Saddler, J.N. Comparison of colorimetric and HPLC techniques for quantitating the carbohydrate components of steam-treated wood. Appl. Biochem. Biotechnol. 1988, 17, 137–149. [Google Scholar] [CrossRef]
- Rocha, G.J.M. Deslignificaçao de Bagaço de Cana de Açúcar Assistida por Oxigênio. Ph.D. Thesis, University of São Paulo, São Paulo, Brazil, 2000. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. In Oxidants and Antioxidants Part A; Elsevier: Amsterdam, The Netherlands, 1999; pp. 152–178. [Google Scholar] [CrossRef]
- Silva, J.P.A.; Mussatto, S.I.; Roberto, I.C.; Teixeira, J.A. Fermentation medium and oxygen transfer conditions that maximize the xylose conversion to ethanol by Pichia stipitis. Renew. Energy 2012, 37, 259–265. [Google Scholar] [CrossRef]
- Stanbury, P.F.; Whitaker, A.; Hall, S.J. Principles of Fermentation Technology, 2nd ed.; Elsevier Science Ltd.: Amsterdam, The Netherlands, 1995; pp. 247–252. [Google Scholar]
- Lindegren, C.C.; Nagai, S.; Nagai, H. Induction of respiratory deficiency in yeast by manganese, copper, cobalt and nickel. Nature 2010, 182, 446–448. [Google Scholar] [CrossRef]
- Roberto, I.C.; Mancilha, I.M.; de Souza, C.A.; Felipe, M.G.A.; Sato, S.; de Castro, H.F. Evaluation of rice straw hemicellulose hydrolysate in the production of xylitol by Candida guilliermondii. Biotechnol. Lett. 1994, 16, 1211–1216. [Google Scholar] [CrossRef]
- Sánchez, S.; Bravo, V.; Castro, E.; Moya, A.J.; Camacho, F. The fermentation of mixtures of D-glucose and D-xylose by Candida shehatae, Pichia stipitis or Pachysolen tannophilus to produce ethanol. J. Chem. Technol. Biotechnol. 2002, 77, 641–648. [Google Scholar] [CrossRef]
- Rocha, G.J.M.; Nascimento, V.M.; Gonçalves, A.R.; Nunes-Silva, V.F.; Martín, C. Influence of mixed sugarcane bagasse samples evaluated by elemental and physical–chemical composition. Ind. Crop. Prod. 2015, 64, 52–58. [Google Scholar] [CrossRef]
- Matei, J.C.; Soares, M.; Bonato, A.C.H.; de Freitas, M.P.A.; Helm, C.V.; Maroldi, W.V.; Magalhães, W.L.; Haminiuk, C.; Maciel, G. Enzymatic delignification of sugar cane bagasse and rice husks and its effect in saccharification. Renew. Energy 2020, 157, 987–997. [Google Scholar] [CrossRef]
- Niju, S.; Swathika, M. Delignification of sugarcane bagasse using pretreatment strategies for bioethanol production. Biocatal. Agric. Biotechnol. 2019, 20, 101263. [Google Scholar] [CrossRef]
- Fonseca, B.G.; Puentes, J.G.; Mateo, S.; Sánchez, S.; Moya, A.J.; Roberto, I.C. Detoxification of rice straw and olive tree pruning hemicellulosic hydrolysates employing Saccharomyces cerevisiae and its effect on the ethanol production by Pichia stipitis. J. Agric. Food Chem. 2013, 61, 40. [Google Scholar] [CrossRef]
- Dey, N.; Vickram, S.; Thanigaivel, S.; Subbaiya, R.; Kim, W.; Karmegam, N.; Govarthanan, M. Nanomaterials for transforming barrier properties of lignocellulosic biomass towards potential applications—A review. Fuel 2022, 316, 123444. [Google Scholar] [CrossRef]
- Rowell, R.M. Chapter 2. Emerging technologies for material and chemicals from biomass. In Opportunities for Lignocellulosic Materials and Composites; American Chemical Society: Washington, DC, USA, 1992; pp. 26–31. [Google Scholar]
- Fu, Y.; Gao, H.; Yu, H.; Yang, Q.; Peng, H.; Liu, P.; Li, Y.; Hu, Z.; Zhang, R.; Li, J.; et al. Specific lignin and cellulose depolymerization of sugarcane bagasse for maximum bioethanol production under optimal chemical fertilizer pretreatment with hemicellulose retention and liquid recycling. Renew. Energy 2022, 200, 1371–1381. [Google Scholar] [CrossRef]
- Pan, S.; Zabed, H.; Wei, Y.; Qi, X. Technoeconomic and environmental perspectives of biofuel production from sugarcane bagasse: Current status, challenges and future outlook. Ind. Crop. Prod. 2022, 188, 115684. [Google Scholar] [CrossRef]
- Howard, R.L.; Abotsi, E.; van Rensburg, E.J.; Howard, S. Lignocellulose biotechnology: Issues of bioconversion and enzyme production. Afr. J. Biotechnol. 2003, 2, 602–619. [Google Scholar] [CrossRef]
- Cherubini, F.; Ulgiati, S. Crop residues as raw materials for biorefinery systems—A LCA case study. Appl. Energy 2010, 87, 47–57. [Google Scholar] [CrossRef]
- Jin, Y.; Shi, Z.; Xu, G.; Yang, H.; Yang, J. A stepwise pretreatment of sugarcane bagasse by alkaline and hydroxymethyl reagent for bioethanol production. Ind. Crop. Prod. 2020, 145, 112136. [Google Scholar] [CrossRef]
- Rencoret, J.; Gutiérrez, A.; Nieto, L.; Jiménez-Barbero, J.; Faulds, C.B.; Kim, H.; Ralph, J.; Martínez, Á.T.; del Río, J. Lignin composition and structure in young versus adult Eucalyptus globulus plants. Plant Physiol. 2010, 155, 667–682. [Google Scholar] [CrossRef] [PubMed]
- Miranda, I.; Gominho, J.; Mirra, I.; Pereira, H. Fractioning and chemical characterization of barks of Betula pendula and Eucalyptus globulus. Ind. Crop. Prod. 2013, 41, 299–305. [Google Scholar] [CrossRef]
- García-Martín, J.F.; Cuevas, M.; Bravo, V.; Sánchez, S. Ethanol production from olive prunings by autohydrolysis and fermentation with Candida tropicalis. Renew. Energy 2010, 35, 1602–1608. [Google Scholar] [CrossRef]
- García-Martín, J.F.; Cuevas, M.; Feng, C.; Álvarez-Mateos, P.; Torres-García, M.; Sánchez, S. Energetic valorisation of olive biomass: Olive-tree pruning, olive stones and pomaces. Processes 2020, 8, 511. [Google Scholar] [CrossRef]
- Manzanares, P.; Ruiz, E.; Ballesteros, M.; Negro, M.; Gallego, F.; López-Linares, J.; Castro, E. Residual biomass potential in olive tree cultivation and olive oil industry in Spain: Valorization proposal in a biorefinery context. Span. J. Agric. Res. 2017, 15, e0206. [Google Scholar] [CrossRef]
- Romero-García, J.M.; López-Linares, J.C.; Contreras, M.; Romero, I.; Castro, E. Exploitation of olive tree pruning biomass through hydrothermal pretreatments. Ind. Crop. Prod. 2022, 176, 114425. [Google Scholar] [CrossRef]
- Mamaní, A.; Maturano, Y.; Mestre, V.; Montoro, L.; Gassa, L.; Deiana, C.; Sardella, F. Valorization of olive tree pruning. Application for energy storage and biofuel production. Ind. Crop. Prod. 2021, 173, 114082. [Google Scholar] [CrossRef]
- Garcia-Maraver, A.; Salvachúa, D.; Martínez, M.; Diaz, L.; Zamorano, M. Analysis of the relation between the cellulose, hemicellulose and lignin content and the thermal behavior of residual biomass from olive trees. Waste Manag. 2013, 33, 2245–2249. [Google Scholar] [CrossRef]
- Mateo, S.; Mateo, P.; Barbanera, M.; Buratti, C.; Moya, A.J. Acid hydrolysis of olive tree leaves: Preliminary study towards biochemical conversion. Processes 2020, 8, 886. [Google Scholar] [CrossRef]
- Fonseca, B.G.; Mateo, S.; Roberto, I.C.; Sánchez, S.; Moya, A.J. Bioconversion in batch bioreactor of olive-tree pruning biomass optimizing treatments for ethanol production. Biochem. Eng. J. 2020, 164, 107793. [Google Scholar] [CrossRef]
- Oliva, J.M.; Negro, M.J.; Álvarez, C.; Manzanares, P.; Moreno, A.D. Fermentation strategies for the efficient use of olive tree pruning biomass from a flexible biorefinery approach. Fuel 2020, 277, 118171. [Google Scholar] [CrossRef]
- Barbanera, M.; Buratti, C.; Cotana, F.; Foschini, D.; Lascaro, E. Effect of steam explosion pretreatment on sugar production by enzymatic hydrolysis of olive tree pruning. Energy Procedia 2015, 81, 146–154. [Google Scholar] [CrossRef]
- Peinado, S.; Mateo, S.; Sánchez, S.; Moya, A.J. Effectiveness of sodium borohydride treatment on acid hydrolyzates from olive-tree pruning biomass for bioethanol production. BioEnergy Res. 2019, 12, 302–311. [Google Scholar] [CrossRef]
- Mateo, S.; Puentes, J.; Roberto, I.; Moya, A.; Sánchez, S. A new approximation on olive pruning by-product characterization. In Proceedings of the ANQUE International Congress of Chemical Engineering (ICCE), Sevilla, Spain, 24–27 June 2012. [Google Scholar]
- Servian-Rivas, L.D.; Ruiz-Pachón, E.; Rodríguez, M.; González-Miquel, M.; González, E.; Díaz, I. Techno-economic and environmental impact assessment of an olive tree pruning waste multiproduct biorefinery. Food Bioprod. Process. 2022, 134, 95–108. [Google Scholar] [CrossRef]
- Canilha, L.; Carvalho, W.; Felipe, M.G.A.; Silva, J.B.A. Xylitol production from wheat straw hemicellulosic hydrolysate: Hydrolysate detoxification and carbon source used for inoculum preparation. Braz. J. Microbiol. 2008, 39, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.J.; Peinado, S.; Mateo, S.; Fonseca, B.G.; Sánchez, S. Improving bioethanol production from olive pruning biomass by deacetylation step prior acid hydrolysis and fermentation processes. Bioresour. Technol. 2016, 220, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.D.; Felipe, M.; Mancilha, I.; Luchese, R.; Silva, S.S. Inhibitory effect of acetic acid on bioconversion of xylose in xylitol by Candida guilliermondii in sugarcane bagasse hydrolysate. Braz. J. Microbiol. 2004, 35, 248–254. [Google Scholar] [CrossRef]
- Rodrigues, R.; Felipe, M.; Almeida e Silva, J.; Vitolo, M.; Gómez, P. The influence of pH, temperature and hydrolyzate concentration on the removal of volatile and nonvolatile compounds from sugarcane bagasse hemicellulosic hydrolyzate treated with activated charcoal before or after vacuum evaporation. Braz. J. Chem. Eng. 2001, 18, 299–311. [Google Scholar] [CrossRef]
- Felipe, M.; Alves, L.; Silva, S.; Roberto, I.; Mancilha, I.; e Silva, J.A. Fermentation of eucalyptus hemicellulosic hydrolysate to xylitol by Candida guilliermondii. Bioresour. Technol. 1996, 56, 281–283. [Google Scholar] [CrossRef]
- Castro, J.F.; Parra, C.; Yáñez-S, M.; Rojas, J.; Mendonça, R.T.; Baeza, J.; Freer, J. Optimal pretreatment of Eucalyptus globulus by hydrothermolysis and alkaline extraction for microbial production of ethanol and xylitol. Ind. Eng. Chem. Res. 2013, 52, 5713–5720. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Dragone, G.; Roberto, I.C. Influence of the toxic compounds present in brewer’s spent grain hemicellulosic hydrolysate on xylose-to-xylitol bioconversion by Candida guilliermondii. Process. Biochem. 2005, 40, 3801–3806. [Google Scholar] [CrossRef]
- Cortez, D.V.; Roberto, I.C. Individual and interaction effects of vanillin and syringaldehyde on the xylitol formation by Candida guilliermondii. Bioresour. Technol. 2010, 101, 1858–1865. [Google Scholar] [CrossRef]
- Wikandari, R.; Sanjaya, A.P.; Millati, R.; Karimi, K.; Taherzadeh, M.J. Fermentation inhibitors in ethanol and biogas processes and strategies to counteract their effects. In Biofuels: Alternative Feedstocks and Conversion Processes for the Production of Liquid and Gaseous Biofuels; Elsevier: Amsterdam, The Netherlands, 2019; pp. 461–499. [Google Scholar] [CrossRef]
- Cuevas, M.; Sánchez, S.; Bravo, V.; Cruz, N.; García, J.F. Fermentation of enzymatic hydrolysates from olive stones by Pachysolen tannophilus. J. Chem. Technol. Biotechnol. 2009, 84, 461–467. [Google Scholar] [CrossRef]
- Cuevas, M.; García, J.F.; Hodaifa, G.; Sánchez, S. Oligosaccharides and sugars production from olive stones by autohydrolysis and enzymatic hydrolysis. Ind. Crop. Prod. 2015, 70, 100–106. [Google Scholar] [CrossRef]
- Luo, C.; Brink, D.; Blanch, H. Identification of potential fermentation inhibitors in conversion of hybrid poplar hydrolyzate to ethanol. Biomass Bioenergy 2002, 22, 125–138. [Google Scholar] [CrossRef]
- Liu, Z.L.; Blaschek, H.P. Biomass conversion inhibitors and in situ detoxification. In Biomass to Biofuels: Strategies for Global Industries; Wiley: Chichester, UK, 2010; pp. 233–259. [Google Scholar]
- Mussatto, S. Biomass pretreatment with acids. In Biomass Fractionation Technologies for a Lignocellulosic Feedstock Based Biorefinery; Elsevier: Amsterdam, The Netherlands, 2016; pp. 169–185. [Google Scholar] [CrossRef]
- Mateo, S.; Peinado, S.; Morillas-Gutiérrez, F.; Rubia, M.D.L.; Moya, A.J. Nanocellulose from Agricultural Wastes: Products and Applications—A Review. Processes 2021, 9, 1594. [Google Scholar] [CrossRef]
- Rao, R.S.; Jyothi, C.P.; Prakasham, R.; Sarma, P.; Rao, L.V. Xylitol production from corn fiber and sugarcane bagasse hydrolysates by Candida tropicalis. Bioresour. Technol. 2006, 97, 1974–1978. [Google Scholar] [CrossRef] [PubMed]
- Mateo, S.; Puentes, J.; Moya, A.; Sánchez, S. Ethanol and xylitol production by fermentation of acid hydrolysate from olive pruning with Candida tropicalis NBRC 0618. Bioresour. Technol. 2015, 190, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, M.; García-Martín, J.; Bravo, V.; Sánchez, S. Ethanol production from olive stones through liquid hot water pre-treatment, enzymatic hydrolysis and fermentation. Influence of enzyme loading and pre-treatment temperature and time. Fermentation 2021, 7, 25. [Google Scholar] [CrossRef]
- Agbogbo, F.K.; Coward-Kelly, G.; Torry-Smith, M.; Wenger, K.; Jeffries, T.W. The effect of initial cell concentration on xylose fermentation by Pichia stipitis. In Applied Biochemistry and Biotecnology; Humana Press: New York, NY, USA, 2007; pp. 653–662. [Google Scholar] [CrossRef]
- Milessi, T.S.; Chandel, A.K.; Branco, R.F.; Silva, S.S. Effect of dissolved oxygen and inoculum concentration on xylose reductase production from Candida guilliermondii using sugarcane bagasse hemicellulosic hydrolysate. Food Nutr. Sci. 2011, 2, 235–240. [Google Scholar] [CrossRef]
- Pessani, N.K.; Atiyeh, H.K.; Wilkins, M.R.; Bellmer, D.D.; Banat, I.M. Simultaneous saccharification and fermentation of kanlow switchgrass by thermotolerant Kluyveromyces marxianus IMB3: The effect of enzyme loading, temperature and higher solid loadings. Bioresour. Technol. 2011, 102, 10618–10624. [Google Scholar] [CrossRef]
- Hickert, L.R.; Cunha-Pereira, F.; Souza-Cruz, P.B.; Rosa, C.A.; Ayub, M.A.Z. Ethanogenic fermentation of co-cultures of Candida shehatae HM 52.2 and Saccharomyces cerevisiae ICV D254 in synthetic medium and rice hull hydrolysate. Bioresour. Technol. 2013, 131, 508–514. [Google Scholar] [CrossRef] [PubMed]
- García, J.F. Aprovechamiento del residuo de poda de olivo. Hidrólisis ácida y Fermentación con Candida tropicalis. Ph.D. Thesis, University of Jaén, Jaén, Spain, 2007. [Google Scholar]
- Mateo, S. Producción de xilitol con Debaryomyces hansenii NRRL Y-7426 y Candida tropicalis NBRC 0618. Aplicación al residuo de poda de olivo. Ph.D. Thesis, University of Jaén, Jaén, Spain, 2011. [Google Scholar]
- Medeiros, L.L.; Silva, F.L.H.; Santos, S.F.M.; Madruga, M.S.; Melo, D.J.N.; Conrado, L.S. Bioconversion of hydrolyzed cashew peduncle bagasse for ethanol and xylitol production. Rev. Bras. Eng. Agrícola Ambient. 2017, 21, 488–492. [Google Scholar] [CrossRef]
- Lohmeier-Vogel, E.M.; Sopher, C.R.; Lee, H. Intracellular acidification as a mechanism for the inhibition by acid hydrolysis-derived inhibitors of xylose fermentation by yeasts. J. Ind. Microbiol. Biotechnol. 1998, 20, 75–81. [Google Scholar] [CrossRef]
- Scordia, D.; Cosentino, S.L.; Lee, J.; Jeffries, T.W. Bioconversion of giant reed (Arundo donax L.) hemicellulose hydrolysate to ethanol by Scheffersomyces stipitis CBS6054. Biomass Bioenergy 2012, 39, 296–305. [Google Scholar] [CrossRef]
- Pant, S.; Ritika; Prakash, A.; Kuila, A. Integrated production of ethanol and xylitol from Brassica juncea using Candida sojae JCM 1644. Bioresour. Technol. 2022, 351, 126903. [Google Scholar] [CrossRef]
- Aguiar, W.; Faria, L.; Couto, M.; Araujo, O.; Pereira, N. Growth model and prediction of oxygen transfer rate for xylitol production from D-xylose by C. guilliermondii. Biochem. Eng. J. 2002, 12, 49–59. [Google Scholar] [CrossRef]
- Schirmer-Michel, Â.C.; Flôres, S.H.; Hertz, P.F.; Ayub, M.A.Z. Effect of oxygen transfer rates on alcohols production by Candida guilliermondii cultivated on soybean hull hydrolysate. J. Chem. Technol. Biotechnol. 2009, 84, 223–228. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | % |
|---|---|
| Cellulose | 33.85 ± 0.76 |
| Hemicellulose | 18.63 ± 0.27 |
| Acid-insoluble lignin | 18.93 ± 0.08 |
| Acid-soluble lignin | 4.19 ± 0.03 |
| Ash | 4.62 ± 0.28 |
| Acetyl groups | 2.07 ± 0.02 |
| Water extracts | 17.39 ± 0.32 |
| Ethanol extracts | 1.79 ± 0.01 |
| Parameter | OTPB-RH | OTPB-LH | SM-R | SM-L |
|---|---|---|---|---|
| , kg kg | 0.05 ± 0.01 | 0.06 ± 0.01 | 0.10 ± 0.01 | 0.09 ± 0.01 |
| , kg kg | 0.33 ± 0.01 | 0.36 ± 0.01 | 0.19 ± 0.01 | 0.38 ± 0.01 |
| , kg m | 15.09 ± 0.01 | 18.42 ± 0.18 | 6.16 ± 0.03 | 15.15 ± 0.05 |
| , kg m h | 0.17 ± 0.01 | 0.24 ± 0.01 | 0.32 ± 0.01 | 0.57 ± 0.01 |
| , kg kg | 0.25 ± 0.01 | 0.39 ± 0.01 | 0.56 ± 0.01 | 0.40 ± 0.01 |
| , kg m | 7.98 ± 0.01 | 10.84 ± 0.17 | 24.99 ± 0.31 | 13.74 ± 0.02 |
| , kg m h | 0.07 ± 0.01 | 0.12 ± 0.01 | 0.54 ± 0.01 | 0.19 ± 0.01 |
| Experiment | OTPB-L1 | OTPB-L2 | OTPB-L3 | OTPB-L4 | OTPB-L5 |
|---|---|---|---|---|---|
| , kg kg | 0.11 ± 0.01 | 0.10 ± 0.01 | 0.08 ± 0.01 | 0.07 ± 0.01 | 0.07 ± 0.01 |
| , kg kg | 0.10 ± 0.01 | 0.10 ± 0.01 | 0.08 ± 0.01 | 0.07 ± 0.01 | 0.07 ± 0.01 |
| , kg kg | 0.11 ± 0.01 | 0.25 ± 0.01 | 0.27 ± 0.01 | 0.33 ± 0.01 | 0.36 ± 0.01 |
| , kg m | 8.68 ± 0.31 | 8.08 ± 0.06 | 10.19 ± 0.33 | 10.17 ± 0.74 | 12.94 ± 0.07 |
| , kg m h | 0.06 ± 0.01 | 0.07 ± 0.01 | 0.06 ± 0.01 | 0.10 ± 0.01 | 0.14 ± 0.01 |
| , kg kg | 0.23 ± 0.01 | 0.26 ± 0.01 | 0.35 ± 0.01 | 0.36 ± 0.02 | 0.38 ± 0.02 |
| , kg m | 7.38 ± 0.08 | 7.88 ± 0.13 | 7.68 ± 0.33 | 8.15 ± 0.05 | 9.15 ± 0.34 |
| , kg m h | 0.04 ± 0.01 | 0.05 ± 0.01 | 0.04 ± 0.01 | 0.06 ± 0.01 | 0.07 ± 0.01 |
| Fermentative Parameters | OTPB-BB | OTPB-EF |
|---|---|---|
| b (kg m h) | 0.06 | |
| (kg kg) | 0.06 | 0.06 ± 0.01 |
| Initial D-glucose concentration (kg m) | 33.44 | 34.88 ± 0.15 |
| Initial D-xylose concentration (kg m) | 22.14 | 23.29 ± 0.08 |
| Maximum ethanol concentration (kg m) | 13.32 | 14.91 ± 0.12 |
| Time to reach the maximum ethanol concentration (h) | 36 | 48 |
| (kg kg) | 0.28 | 0.38 ± 0.01 |
| (kg m h) | 0.84 | 0.57 ± 0.01 |
| Ethanol yield | 54.90 | 74.51 ± 0.04 |
| Maximum xylitol concentration (kg m) | 9.13 | 8.11 ± 0.11 |
| Time to reach the maximum xylitol concentration (h) | 66 | 48 |
| (kg kg) | 0.37 | 0.46 ± 0.02 |
| (kg m h) | 0.26 | 0.17 ± 0.01 |
| Xylitol yield (% of theoretical value) | 40.66 | 50.55 ± 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puentes, J.G.; Mateo, S.; Sánchez, S.; Roberto, I.C.; Moya, A.J. Bioconversion Study of Olive Tree Biomass Hemicellulosic Hydrolysates by Candida guilliermondii at Different Scales for Ethanol and Xylitol Production. Fermentation 2023, 9, 553. https://doi.org/10.3390/fermentation9060553
Puentes JG, Mateo S, Sánchez S, Roberto IC, Moya AJ. Bioconversion Study of Olive Tree Biomass Hemicellulosic Hydrolysates by Candida guilliermondii at Different Scales for Ethanol and Xylitol Production. Fermentation. 2023; 9(6):553. https://doi.org/10.3390/fermentation9060553
Chicago/Turabian StylePuentes, Juan Gabriel, Soledad Mateo, Sebastian Sánchez, Inês C. Roberto, and Alberto J. Moya. 2023. "Bioconversion Study of Olive Tree Biomass Hemicellulosic Hydrolysates by Candida guilliermondii at Different Scales for Ethanol and Xylitol Production" Fermentation 9, no. 6: 553. https://doi.org/10.3390/fermentation9060553
APA StylePuentes, J. G., Mateo, S., Sánchez, S., Roberto, I. C., & Moya, A. J. (2023). Bioconversion Study of Olive Tree Biomass Hemicellulosic Hydrolysates by Candida guilliermondii at Different Scales for Ethanol and Xylitol Production. Fermentation, 9(6), 553. https://doi.org/10.3390/fermentation9060553