
Advances in the Production of Theanine by Plants and Microorganisms

 and
and
Abstract
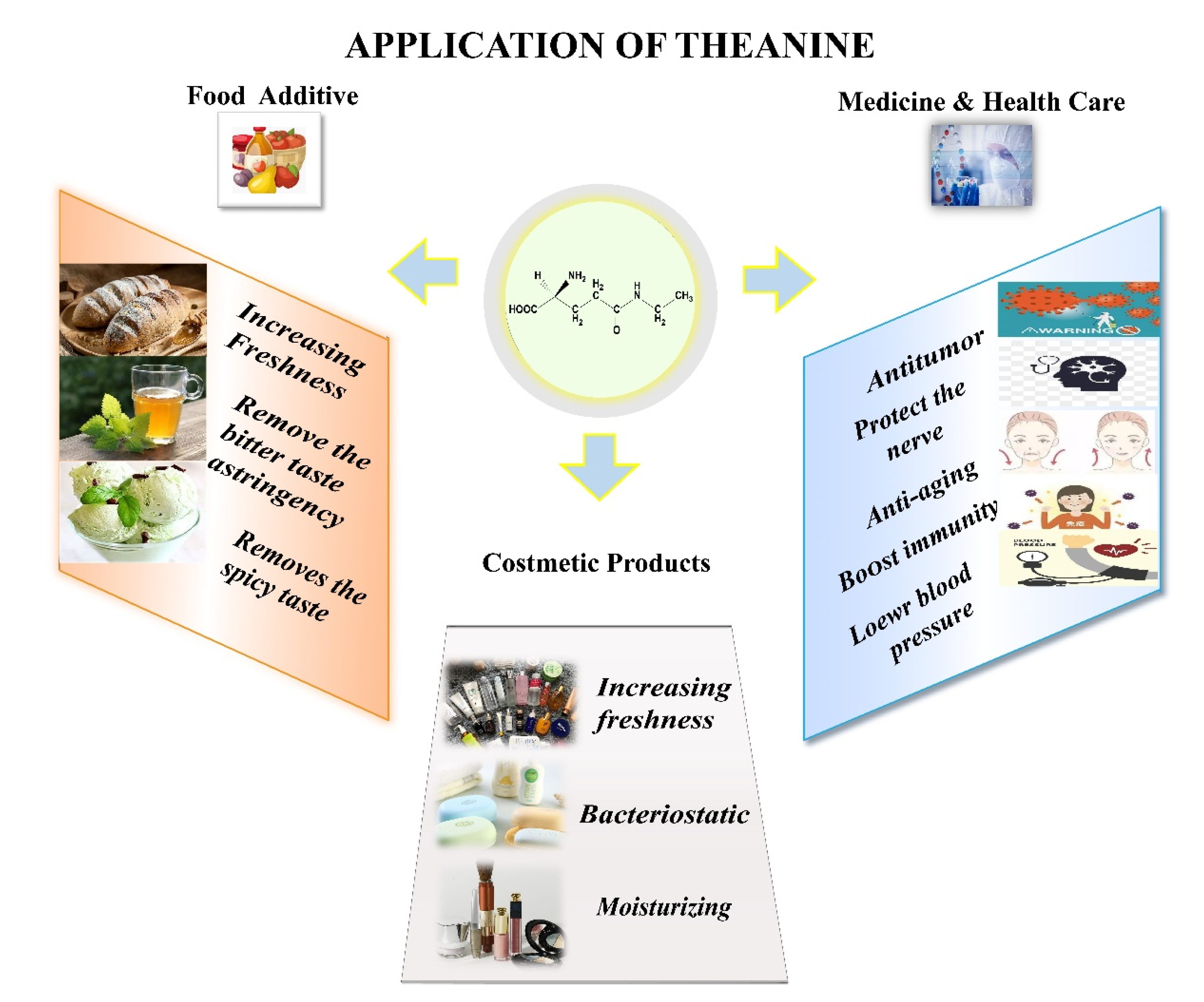
1. Introduction
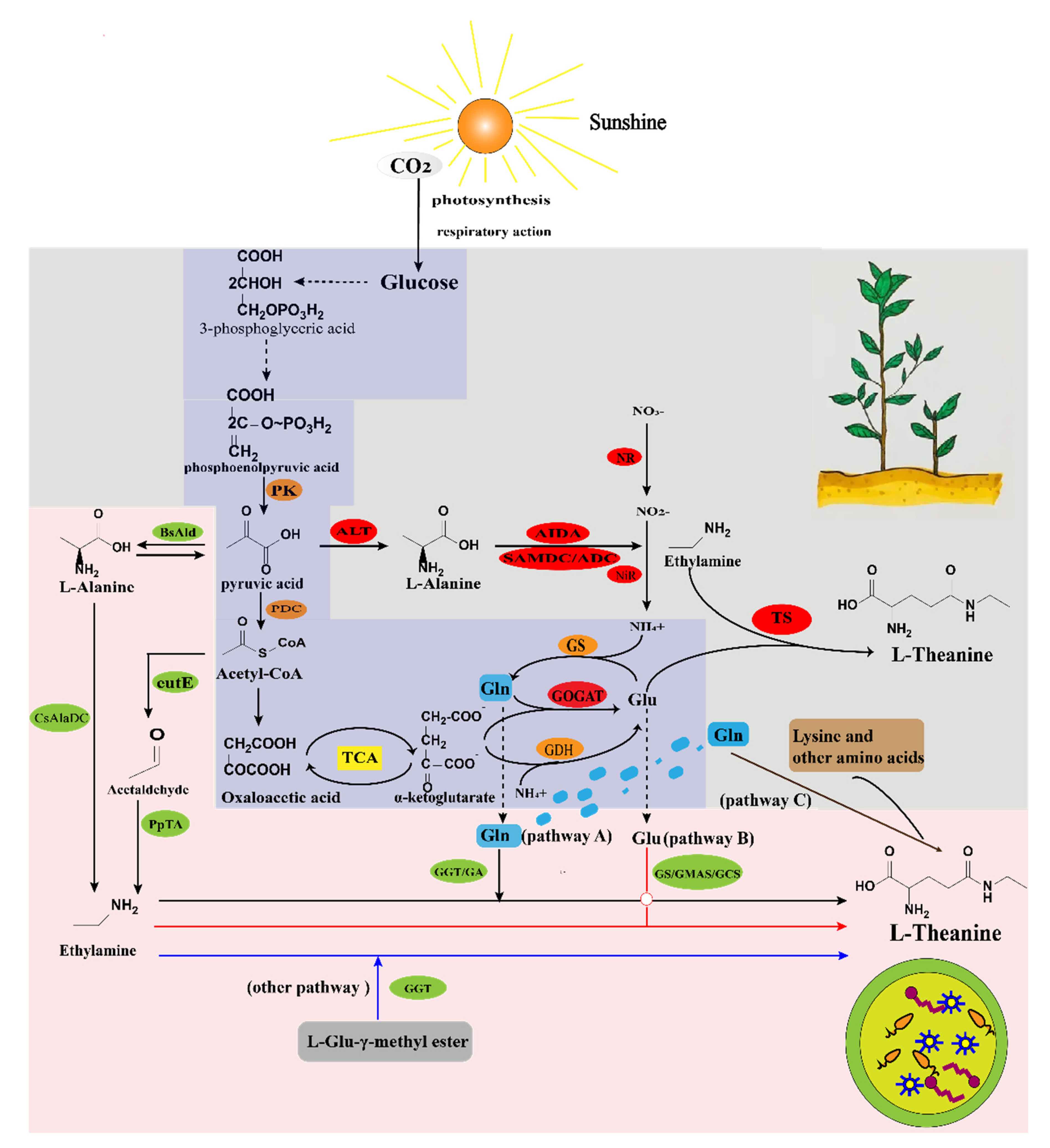
2. The Biosynthetic Pathway of L-Theanine in Plants
2.1. The Biosynthetic Pathway of L-Theanine in Tea Tree
2.2. The Biosynthetic Pathway of L-Theanine in Other Plants
3. The Synthesis Pathway of L-Theanine in Microorganisms
3.1. Glutamate-Mediated Pathway of L-Theanine in Microorganisms
3.1.1. Enzyme-Catalyzed Method
3.1.2. Whole-Cell Catalytic Method
3.1.3. Ethanolamine Flow plus Microbial Fermentation Method
3.1.4. One-Step Fermentation Method
3.2. Glutamine-Mediated Pathway of L-Theanine in Microorganisms
3.3. Other Substrate-Mediated Pathways of L-Theanine in Microorganisms
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsushida, T.; Takeo, T. Occurrence of Theanine in Camellia Japonica and Camellia sasanqua Seedlings. Agric. Biol. Chem. 2014, 48, 2861–2862. [Google Scholar] [CrossRef]
- Williams, J.L.; Everett, J.M.; D’Cunha, N.M.; Sergi, D.; Georgousopoulou, E.N.; Keegan, R.J.; McKune, A.J.; Mellor, D.D.; Anstice, N.; Naumovski, N. The Effects of Green Tea Amino Acid L-Theanine Consumption on the Ability to Manage Stress and Anxiety Levels: A Systematic Review. Plant Foods Hum. Nutr. 2020, 75, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, K.; Akaihata, H.; Hata, J.; Tanji, R.; Honda-Takinami, R.; Onagi, A.; Hoshi, S.; Koguchi, T.; Sato, Y.; Kataoka, M.; et al. L-Theanine Protects Bladder Function by Suppressing Chronic Sympathetic Hyperactivity in Spontaneously Hypertensive Rat. Metabolites 2021, 11, 778. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, Y.; Zhang, H.; Ji, D.; Wu, F.; Tian, H.; Liu, K.; Zhang, Y.; Wu, B.; Zhang, G. Theanine from tea and its semi-synthetic derivative TBrC suppress human cervical cancer growth and migration by inhibiting EGFR/Met-Akt/NF-κB signaling. Eur. J. Pharmacol. 2016, 791, 297–307. [Google Scholar] [CrossRef]
- Wong, P.-P.; Bodrug, N.; Hodivala-Dilke, K.M. Exploring novel methods for modulating tumor blood vessels in cancer treatment. Curr. Biol. 2016, 26, R1161–R1166. [Google Scholar] [CrossRef]
- Lin, S.; Chen, Z.; Chen, T.; Deng, W.; Wan, X.; Zhang, Z. Theanine metabolism and transport in tea plants (Camellia sinensis L.): Advances and perspectives. Crit. Rev. Biotechnol. 2023, 43, 327–341. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, Z.; Yuan, H.; He, N. From Tea Leaves to Factories: A Review of Research Progress in L-Theanine Biosynthesis and Production. J. Agric. Food Chem. 2021, 69, 1187–1196. [Google Scholar] [CrossRef]
- Lin, N.; Liu, X.; Zhu, W.; Cheng, X.; Wang, X.; Wan, X.; Liu, L. Ambient ultraviolet B signal modulates tea flavor characteristics via shifting a metabolic flux in flavonoid biosynthesis. J. Agric. Food Chem. 2021, 69, 3401–3414. [Google Scholar] [CrossRef]
- Fu, Y.-Q.; Wang, J.-Q.; Chen, J.-X.; Wang, F.; Yin, J.-F.; Zeng, L.; Shi, J.; Xu, Y.-Q. Effect of baking on the flavor stability of green tea beverages. Food Chem. 2020, 331, 127258. [Google Scholar] [CrossRef]
- Liu, S.H.; Li, J.; Huang, J.A.; Liu, Z.H.; Xiong, L.G. New advances in genetic engineering for L-theanine biosynthesis. Trends Food Sci. Technol. 2021, 114, 540–551. [Google Scholar] [CrossRef]
- Xu, Y.-M.; Qiao, F.-B.; Huang, J.-K. Black tea markets worldwide: Are they integrated? J. Integr. Agric. 2022, 21, 552–565. [Google Scholar]
- Sharma, E.; Joshi, R.; Gulati, A. D-Theanine: An astounding sui generis integrant in tea. Food Chem. 2018, 242, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Soda, K.; Uchiyama, K.; Ogata, K. Metabolism of L-Theanine, D-Theanine and the Related Compounds in Bacteria: Part II. Purification and Properties of the Enzyme Hydrolyzing L-Theanine, D-Theanine and the Related Compounds. Agric. Biol. Chem. 1966, 30, 541–557. [Google Scholar] [CrossRef]
- Desai, M.J.; Gill, M.S.; Hsu, W.H.; Armstrong, D.W. Pharmacokinetics of theanine enantiomers in rats. Chirality 2005, 17, 154–162. [Google Scholar] [CrossRef]
- Saeed, M.; Khan, M.S.; Kamboh, A.A.; Alagawany, M.; Khafaga, A.F.; Noreldin, A.E.; Qumar, M.; Safdar, M.; Hussain, M.; Abd El-Hack, M.E. L-theanine: An astounding sui generis amino acid in poultry nutrition. Poult. Sci. 2020, 99, 5625–5636. [Google Scholar] [CrossRef]
- Zhao, J.; Li, P.; Xia, T.; Wan, X. Exploring plant metabolic genomics: Chemical diversity, metabolic complexity in the biosynthesis and transport of specialized metabolites with the tea plant as a model. Crit. Rev. Biotechnol. 2020, 40, 667–688. [Google Scholar] [CrossRef]
- Takeo, T. L-Alanine as a precursor of ethylamine in Camellia sinensis. Phytochemistry 1974, 13, 1401–1406. [Google Scholar] [CrossRef]
- Cheng, S.; Fu, X.; Wang, X.; Liao, Y.; Zeng, L.; Dong, F.; Yang, Z. Studies on the Biochemical Formation Pathway of the Amino Acid L-Theanine in Tea (Camellia sinensis) and Other Plants. J. Agric. Food Chem. 2017, 65, 7210–7216. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liao, Y.; Cheng, S.; Deng, R.; Yang, Z. Stable Isotope-Labeled Precursor Tracing Reveals that L-Alanine is Converted to L-Theanine via L-Glutamate not Ethylamine in Tea Plants In Vivo. J. Agric. Food Chem. 2021, 69, 15354–15361. [Google Scholar] [CrossRef]
- Bai, P.; Wang, L.; Wei, K.; Ruan, L.; Wu, L.; He, M.; Ni, D.; Cheng, H. Biochemical characterization of specific Alanine Decarboxylase (AlaDC) and its ancestral enzyme Serine Decarboxylase (SDC) in tea plants (Camellia sinensis). BMC Biotechnol. 2021, 21, 17. [Google Scholar] [CrossRef]
- Bai, P.; Wei, K.; Wang, L.; Zhang, F.; Ruan, L.; Li, H.; Wu, L.; Cheng, H. Identification of a Novel Gene Encoding the Specialized Alanine Decarboxylase in Tea (Camellia sinensis) Plants. Molecules 2019, 24, 540. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.; Li, H.; Chen, Q.; Zhu, J.; Wu, X.; Luo, S.; Zhao, Y.; Wang, L.; Jiang, S.; Li, X.; et al. Secretion of Bacillus amyloliquefaciens gamma-Glutamyltranspeptidase from Bacillus subtilis and Its Application in Enzymatic Synthesis of L-Theanine. J. Agric. Food Chem. 2019, 67, 14129–14136. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Xu, C.; Sun, Y.; Niu, C.; Leng, X.; Hao, B.; Ma, J.; Liu, Z.; Xu, Z.; Yang, C.; et al. Genome-wide identification of glutamate synthase gene family and expression patterns analysis in response to carbon and nitrogen treatment in Populus. Gene 2023, 851, 146996. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Elmetwally, F.; Elghanam, R.; Kamal, N.; Hellal, K.; Hamezah, H.S.; Zhao, C.; Mediani, A. Metabolomics in tea products; a compile of applications for enhancing agricultural traits and quality control analysis of Camellia sinensis. Food Chem. 2023, 404, 134628. [Google Scholar] [CrossRef]
- Deng, W.-W.; Ogita, S.; Ashihara, H. Biosynthesis of theanine (γ-ethylamino-L-glutamic acid) in seedlings of Camellia sinensis. Phytochem. Lett. 2008, 1, 115–119. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Z.P.; Jia, X.H.; Wu, Y.F.; Luo, L.; Yin, Z.M. Mn2+ enhances theanine-forming activity of recombinant glutamine synthetase from Bacillus subtilis in Escherichia coli. World J. Microbiol. Biotechnol. 2007, 24, 1267–1272. [Google Scholar] [CrossRef]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Xia, E.; Lu, Y.; Tai, Y.; She, G.; et al. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, E4151–E4158. [Google Scholar] [CrossRef]
- Yamamoto, S.; Wakayama, M.; Tachiki, T. Cloning and expression of Pseudomonas taetrolens Y-30 gene encoding glutamine synthetase: An enzyme available for theanine production by coupled fermentation with energy transfer. Biosci. Biotechnol. Biochem. 2006, 70, 500–507. [Google Scholar] [CrossRef]
- Ashihara, H. Occurrence, biosynthesis and metabolism of theanine (γ-glutamyl-L-ethylamide) in plants: A comprehensive review. Nat. Prod. Commun. 2015, 10, 803–810. [Google Scholar] [CrossRef]
- Xu, H.; Gao, J.; Portieles, R.; Du, L.; Gao, X.; Borras-Hidalgo, O. Endophytic bacterium Bacillus aryabhattai induces novel transcriptomic changes to stimulate plant growth. PLoS ONE 2022, 17, e0272500. [Google Scholar] [CrossRef]
- Li, M.G.; Villemur, R.; Hussey, P.J.; Silflow, C.D.; Gantt, J.S.; Snustad, D.P. Differential expression of six glutamine synthetase genes in Zea mays. Plant Mol. Biol. 1993, 23, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Scarpeci, T.E.; Marro, M.L.; Bortolotti, S.; Boggio, S.B.; Valle, E.M. Plant nutritional status modulates glutamine synthetase levels in ripe tomatoes (Solanum lycopersicum cv. Micro-Tom). J. Plant Physiol. 2007, 164, 137–145. [Google Scholar] [CrossRef]
- Mu, W.; Zhang, T.; Jiang, B. An overview of biological production of L-theanine. Biotechnol. Adv. 2015, 33, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Han, Y.H.; Park, Y.L.; Park, J.Y.; No, S.Y.; Jeong, D.; Park, S.; Park, H.Y.; Kim, W.; Seo, S.O.; et al. Production of L-Theanine Using Escherichia coli Whole-Cell Overexpressing gamma-Glutamylmethylamide Synthetase with Bakers Yeast. J. Microbiol. Biotechnol. 2020, 30, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Miyake, K.; Kakita, S. A novel catalytic ability of gamma-glutamylcysteine synthetase of Escherichia coli and its application in theanine production. Biosci. Biotechnol. Biochem. 2009, 73, 2677–2683. [Google Scholar] [CrossRef]
- Pan, X.; Yu, J.; Du, Q.; Zeng, S.; Liu, J.; Jiao, Q.; Zhang, H. Efficient synthesis of gamma-glutamyl compounds by co-expression of gamma-glutamylmethylamide synthetase and polyphosphate kinase in engineered Escherichia coli. J. Ind. Microbiol. Biotechnol. 2020, 47, 573–583. [Google Scholar] [CrossRef]
- Yu, Z.; Yang, Z. Understanding different regulatory mechanisms of proteinaceous and non-proteinaceous amino acid formation in tea (Camellia sinensis) provides new insights into the safe and effective alteration of tea flavor and function. Crit. Rev. Food Sci. Nutr. 2020, 60, 844–858. [Google Scholar] [CrossRef]
- Tachiki, T.; Suzuki, H.; Wakisaka, S.; Yano, T.; Tochikura, T. Production of γ-glutamylmethylamide and γ-glutamylethylamide by couplingn of baker’s yesst preparations and bacterial glutamine synthetase. J. Gen. Appl. Microbiol. 1986, 32, 545–548. [Google Scholar] [CrossRef]
- Zhu, W.; Li, X.-H.; Wang, L.-Y.; Fang, W.-P.; Cheng, H. Construction of E. coli recombinant engineered strain for theanine biosynthesis with GS gene embedded. J. Tea Sci. 2008, 28, 242–248. [Google Scholar] [CrossRef]
- Yao, J.; Li, J.; Xiong, D.; Qiu, Y.; Shi, G.; Jin, J.M.; Tao, Y.; Tang, S.Y. Development of a highly efficient and specific L-theanine synthase. Appl. Microbiol. Biotechnol. 2020, 104, 3417–3431. [Google Scholar] [CrossRef]
- Yamamoto, S.; Morihara, Y.; Wakayama, M.; Tachiki, T. Theanine production by coupled fermentation with energy transfer using gamma-glutamylmethylamide synthetase of Methylovorus mays No. 9. Biosci. Biotechnol. Biochem. 2008, 72, 1206–1211. [Google Scholar] [CrossRef]
- Ma, H.; Fan, X.; Cai, N.; Zhang, D.; Zhao, G.; Wang, T.; Su, R.; Yuan, M.; Ma, Q.; Zhang, C.; et al. Efficient fermentative production of L-theanine by Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2020, 104, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Long, Y.K.; Cao, J.H.; Wu, J.S.; Yuan, Y. Production of L-theanine using recombinant Escherichia coli. Food Ferment. Ind. 2019, 45, 6–11. [Google Scholar]
- Fan, X.; Zhang, T.; Ji, Y.; Li, J.; Long, K.; Yuan, Y.; Li, Y.; Xu, Q.; Chen, N.; Xie, X. Pathway engineering of Escherichia coli for one-step fermentative production of L-theanine from sugars and ethylamine. Metab. Eng. Commun. 2020, 11, e00151. [Google Scholar] [CrossRef] [PubMed]
- Benninghaus, L.; Walter, T.; Mindt, M.; Risse, J.M.; Wendisch, V.F. Metabolic Engineering of Pseudomonas putida for Fermentative Production of L-Theanine. J. Agric. Food Chem. 2021, 69, 9849–9858. [Google Scholar] [CrossRef]
- Hagihara, R.; Ohno, S.; Hayashi, M.; Tabata, K.; Endo, H. Production of L-Theanine by Escherichia coli in the Absence of Supplemental Ethylamine. Appl. Environ. Microbiol. 2021, 87, e00031-21. [Google Scholar] [CrossRef]
- Tachiki, T.; Yamada, T.; Mizuno, K.; Ueda, M.; Shiode, J.; Fukami, H. γ-Glutamyl Transfer Reactions by Glutaminase from Pseudomonas nitroreducens IFO 12694 and Their Application for the Syntheses of Theanine and γ-Glutamylmethylamide. Biosci. Biotechnol. Biochem. 1998, 62, 1279–1283. [Google Scholar] [CrossRef]
- Shuai, Y.Y.; Zhang, T.; Jiang, B.; Hua, Y.; Mu, C.F. An efficient method for the high-yield production of L-theanine using a newly isolated glutaminase-producing organism. Food Biosci. 2019, 28, 164–169. [Google Scholar] [CrossRef]
- Alemzadeh, I.; Sakhaei, M. Enzymatic Synthesis of Theanine in the Presence of L-glutaminase Produced by Trichoderma koningii. Appl. Food Biotechnol. 2017, 4, 113–121. [Google Scholar]
- Suzuki, H.; Izuka, S.; Miyakawa, N.; Kumagai, H. Enzymatic production of theanine, an “umami” component of tea, from glutamine and ethylamine with bacterial γ-glutamyltranspeptidase. Enzym. Microb. Technol. 2002, 31, 884–889. [Google Scholar] [CrossRef]
- Jia, H.X.; Chen, L.; Zhao, W.N.; Yin, M.Z. Production of Ltheanine by bioconversion with recombinant γGlutamyltranspeptidase. Sci. Technol. Food Ind. 2008, 29, 166–169. [Google Scholar]
- Bindal, S.; Gupta, R. L-theanine synthesis using gamma-glutamyl transpeptidase from Bacillus licheniformis ER-15. J. Agric. Food Chem. 2014, 62, 9151–9159. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Han, F.; Zhang, X.; Yu, Q. Ultrasound enhanced biosynthesis of L-theanine from L-glutamine and ethylamine by recombinant gamma-glutamyltranspeptidase. Bioresour. Technol. 2020, 307, 123251. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Min, C.; Zhu, F.; Xin, Y.; Zhang, S.; Luo, L.; Yin, Z. Production of bioactive gamma-glutamyl transpeptidase in Escherichia coli using SUMO fusion partner and application of the recombinant enzyme to L-theanine synthesis. Curr. Microbiol. 2011, 62, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bai, W.; Zeng, X.; Cui, C. Gamma glutamyl peptides: The food source, enzymatic synthesis, kokumi-active and the potential functional properties—A review. Trends Food Sci. Technol. 2019, 91, 339–346. [Google Scholar] [CrossRef]
- He, F.; Yang, T.; Xu, M.; Zhang, X.; Rao, Z.; Tang, L. Efficient synthesis of L-theanine by recombinant strain Corynebacterium glutamicum SYPA5-5. Wei Sheng Wu Xue Bao Acta Microbiol. Sin. 2016, 56, 1595–1605. [Google Scholar]
- Huang, F.; Tang, Y.P.; Huang, L.; Cai, J.; Xu, Z.N. High level conversion of L-theanine by recombinant Escherichia coli. Chem. Eng. 2016, 44, 1–4. [Google Scholar]
- Sun, J.; Chang, M.; Li, H.; Zhang, Z.; Chen, Q.; Chen, Y.; Yao, Y.; Pan, A.; Shi, C.; Wang, C.; et al. Endophytic Bacteria as Contributors to Theanine Production in Camellia sinensis. J. Agric. Food Chem. 2019, 67, 10685–10693. [Google Scholar] [CrossRef]
- Zhang, F.; Zheng, Q.Z.; Jiao, Q.C.; Liu, J.Z.; Zhao, G.H. Enzymatic synthesis of theanine from glutamic acid gamma-methyl ester and ethylamine by immobilized Escherichia coli cells with gamma-glutamyltranspeptidase activity. Amino Acids 2010, 39, 1177–1182. [Google Scholar] [CrossRef]
- Tachiki, T.; Suzuki, H.; Wakisaka, S.; Yano, T.; Tochikura, T. Glutamine production in high concentrations with energy transfer employing glutamine synthetase from micrococcus glutamicus. J. Gen. Appl. Microbiol. 1983, 29, 355–363. [Google Scholar] [CrossRef]
- Yamamoto, S.; Uchimura, K.; Wakayama, M.; Tachiki, T. Purification and characterization of glutamine synthetase of Pseudomonas taetrolens Y-30: An enzyme usable for production of theanine by coupling with the alcoholic fermentation system of baker’s yeast. Biosci. Biotechnol. Biochem. 2004, 68, 1888–1897. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Wakayama, M.; Tachiki, T. Theanine Production by Coupled Fermentation with Energy Transfer Employing Pseudomonas taetrolens Y-30 Glutamine Synthetase and Baker’s Yeast Cells. Biosci. Biotechnol. Biochem. 2005, 69, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.C.; Yang, C.; Zhao, Z.H.; Xu, J.J.; Li, J.; Li, P. Application of Cell-Free Protein Synthesis System for the Biosynthesis of L-Theanine. ACS Synth. Biol. 2021, 10, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Hibi, T.; Nii, H.; Nakatsu, T.; Kimura, A.; Kato, H.; Hiratake, J.; Oda, J.i. Crystal structure of γ-glutamylcysteine synthetase: Insights into the mechanism of catalysis by a key enzyme for glutathione homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 15052–15057. [Google Scholar] [CrossRef]
- Kimura, T.; Sugahara, I.; Hanai, K.; Tonomura, Y. Purification and Characterization of gamma-Glutamylmethylamide Synthetase from Methylophaga sp. AA-30. Biosci. Biotechnol. Biochem. 1992, 56, 708–711. [Google Scholar] [CrossRef]
- Yamamoto, S.; Wakayama, M.; Tachiki, T. Characterization of theanine-forming enzyme from Methylovorus mays no. 9 in respect to utilization of theanine production. Biosci. Biotechnol. Biochem. 2007, 71, 545–552. [Google Scholar] [CrossRef]
- Galanie, S.; Entwistle, D.; Lalonde, J. Engineering biosynthetic enzymes for industrial natural product synthesis. Nat. Prod. Rep. 2020, 37, 1122–1143. [Google Scholar] [CrossRef]
- Niu, H.; Li, R.; Liang, Q.; Qi, Q.; Li, Q.; Gu, P. Metabolic engineering for improving L-tryptophan production in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2019, 46, 55–65. [Google Scholar] [CrossRef]
- Kondo, T.; Yumura, S. Strategies for enhancing gene expression in Escherichia coli. Appl. Microbiol. Biotechnol. 2020, 104, 3825–3834. [Google Scholar] [CrossRef]
- Sharma, M.K.; Melosso, M.; Chandra, S. Potential spectral lines of ethanimine (CH3CHNH)-an important possible precursor of amino acids. New Astron. 2023, 98, 101928. [Google Scholar] [CrossRef]
- Yamamoto, S.; Wakayama, M.; Tachiki, T. Cloning and expression of Methylovorus mays No. 9 gene encoding gamma-glutamylmethylamide synthetase: An enzyme usable in theanine formation by coupling with the alcoholic fermentation system of baker’s yeast. Biosci. Biotechnol. Biochem. 2008, 72, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.W.; Wu, Z.J.; Li, H.; Wang, Y.X.; Zhuang, J. L-Theanine content and related gene expression: Novel insights into theanine biosynthesis and hydrolysis among different tea plant (Camellia sinensis L.) tissues and cultivars. Front. Plant Sci. 2017, 8, 498. [Google Scholar] [CrossRef] [PubMed]
- Tabata, K.; Shoto, O. Method of Making Theanine; Kyowa Hakko Bio Co., Ltd.: Tokyo, Japan, 2021. [Google Scholar]
- Yang, T.; Irene, K.; Liu, H.; Liu, S.; Zhang, X.; Xu, M.; Rao, Z. Enhanced extracellular gamma glutamyl transpeptidase production by overexpressing of PrsA lipoproteins and improving its mRNA stability in Bacillus subtilis and application in biosynthesis of L-theanine. J. Biotechnol. 2019, 302, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Zhang, W.G.; Wang, Z.J.; Zhan, Y.P.; Xu, L.S.; Liu, J.Z.; Liu, Q.; Jiao, Q. Enzymatic synthesis of theanine with Escherichia coli γ-glutamyltranspeptidase from a series of γ-glutamyl anilide substrate analogues. Biotechnol. Bioprocess Eng. 2013, 18, 358–364. [Google Scholar] [CrossRef]
- Chen, X.Y.; Su, L.P.; Wu, D.; Wu, J. Application of recombinant Bacillus subtilis γ-glutamyltranspeptidase to the production of L-theanine. Process Biochem. 2014, 49, 1429–1439. [Google Scholar] [CrossRef]
- Wang, H.Q.; Yao, Z.; Zhou, Z.; Sun, Y.; Wei, P.; Ouyang, P.K. Enzymatic synthesis of theanine with L-glutamine-Zn(II) complexes. Biotechnol. Bioprocess Eng. 2013, 17, 1135–1139. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Host | Engineered Strategy | L-Theanine Titer (g/L) | L-Theanine Yield (g/g) | Substrate | Reference | |
|---|---|---|---|---|---|---|
| Pathway (A) | Micrococcus glutamicus | Coupling of cell-free extracts of baker’s yeast with partially purified glutamine synthetase | 0.01 | − | Sodium glutamate | [38] |
| Pathway (A) | E. coli | Expression of GS gene | 29.6 | 0.85 | Sodium glutamate | [28] |
| Pathway (A) | E. coli | Expression of GS gene | 15.3 | 0.45 | Sodium glutamate | [26] |
| Pathway (A) | E. coli | Whole-cell catalytic reaction | 6.2 | 0.178 | Sodium glutamate | [39] |
| Pathway (A) | E. coli | Targeted transformation modification of γ-GCS mutants | 30.4 | 0.871 | Sodium glutamate | [40] |
| Pathway (A) | E. coli | Recombinant GMAS coupled to an ATP regeneration system | 110 | 1.00 | Sodium glutamate | [41] |
| Pathway (A) | E. coli | Co-expressions of PPK and GMAS | 34.67 | 0.66 | Sodium glutamate | [36] |
| Pathway (A) | C. glutamicum | Batch make-up fermentation strategy | 42.0 | 0.196 | Glucose | [42] |
| Pathway (A) | E. coli | Ethanolamine addition | 30.45 | 0.201 | Glucose | [43] |
| Pathway (A) | E. coli | Engineering of a one-step fermentation pathway from sugar and ethylamine | 70.6 | 0.42 | Glucose | [44] |
| Pathway (A) | Pseudomonas putida | Fermentation process using Pseudomonas aeruginosa metabolic engineering compatible with various alternative carbon sources | 17.2 | − | Glucose with xylose | [45] |
| Pathway (A) | E. coli | Establishing a TA pathway without ethylamine supplementation | 1.53 | − | Glucose | [46] |
| Pathway (B) | P. nitroreducens | Enzyme-catalyzed reaction | 47.034 | 0.458 | Glutamine | [47] |
| Pathway (B) | P. nitroreducens | Enzyme-catalyzed reaction | 85.358 | 0.661 | Glutamine | [48] |
| Pathway (B) | Trichoderma koningii | Enzyme-catalyzed reaction | 7.491 | 0.171 | Glutamine | [49] |
| Pathway (B) | E. coli | Enzyme-catalyzed reaction | 20.904 | 0.715 | Glutamine | [50] |
| Pathway (B) | E. coli | Crude enzyme catalysis | 26.884 | 0.578 | Glutamine | [51] |
| Pathway (B) | E. coli | Separation of pure enzyme catalysis | 12.124 | 0.964 | Glutamine | [52] |
| Pathway (B) | E. coli | Point mutation, pure enzyme catalysis | 18.622 | 0.89 | Glutamine | [53] |
| Pathway (B) | E. coli | Expression of GGT in recombinant E. coli using small ubiquitin-related modifier (SUMO) fusion technology | 41 | 0.800 | Glutamine | [54] |
| Pathway (B) | B. subtlis | Separation of pure enzyme catalysis | 32.66 | 0.74 | Glutamine | [55] |
| Pathway (B) | C. glutamicum | Separation of pure enzyme catalysis | 18.17 | 0.898 | Glutamine | [56] |
| Pathway (B) | E. coli | Whole-cell catalysis | 34.650 | 0.790 | Glutamine | [57] |
| Pathway (C) | Luteibacter | Separation of pure enzyme catalysis | 34.223 × 10−6 | − | Glutamine | [58] |
| Other pathways | E. coli | Whole-cell catalysis | 15.3 | 0.95 | Glutamate-γ-methyl ester | [59] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, S.; Qian, R.; Hu, S.; Fu, Z.; Bai, T.; Wang, W.; Cheng, J.; Zhang, J. Advances in the Production of Theanine by Plants and Microorganisms. Fermentation 2023, 9, 543. https://doi.org/10.3390/fermentation9060543
Xiao S, Qian R, Hu S, Fu Z, Bai T, Wang W, Cheng J, Zhang J. Advances in the Production of Theanine by Plants and Microorganisms. Fermentation. 2023; 9(6):543. https://doi.org/10.3390/fermentation9060543
Chicago/Turabian StyleXiao, Shujian, Rong Qian, Shunyang Hu, Zhongdan Fu, Ting Bai, Wei Wang, Jie Cheng, and Jiamin Zhang. 2023. "Advances in the Production of Theanine by Plants and Microorganisms" Fermentation 9, no. 6: 543. https://doi.org/10.3390/fermentation9060543
APA StyleXiao, S., Qian, R., Hu, S., Fu, Z., Bai, T., Wang, W., Cheng, J., & Zhang, J. (2023). Advances in the Production of Theanine by Plants and Microorganisms. Fermentation, 9(6), 543. https://doi.org/10.3390/fermentation9060543