
Impact of Waste as a Substrate on Biomass Formation, and Optimization of Spent Microbial Biomass Re-Use by Sustainable Metabolic Engineering


Abstract
1. Introduction
2. Methods
2.1. Estimating Amounts of SMB from Waste and Non-Waste Substrates
2.2. Estimating Amino Acid Composition of SMB
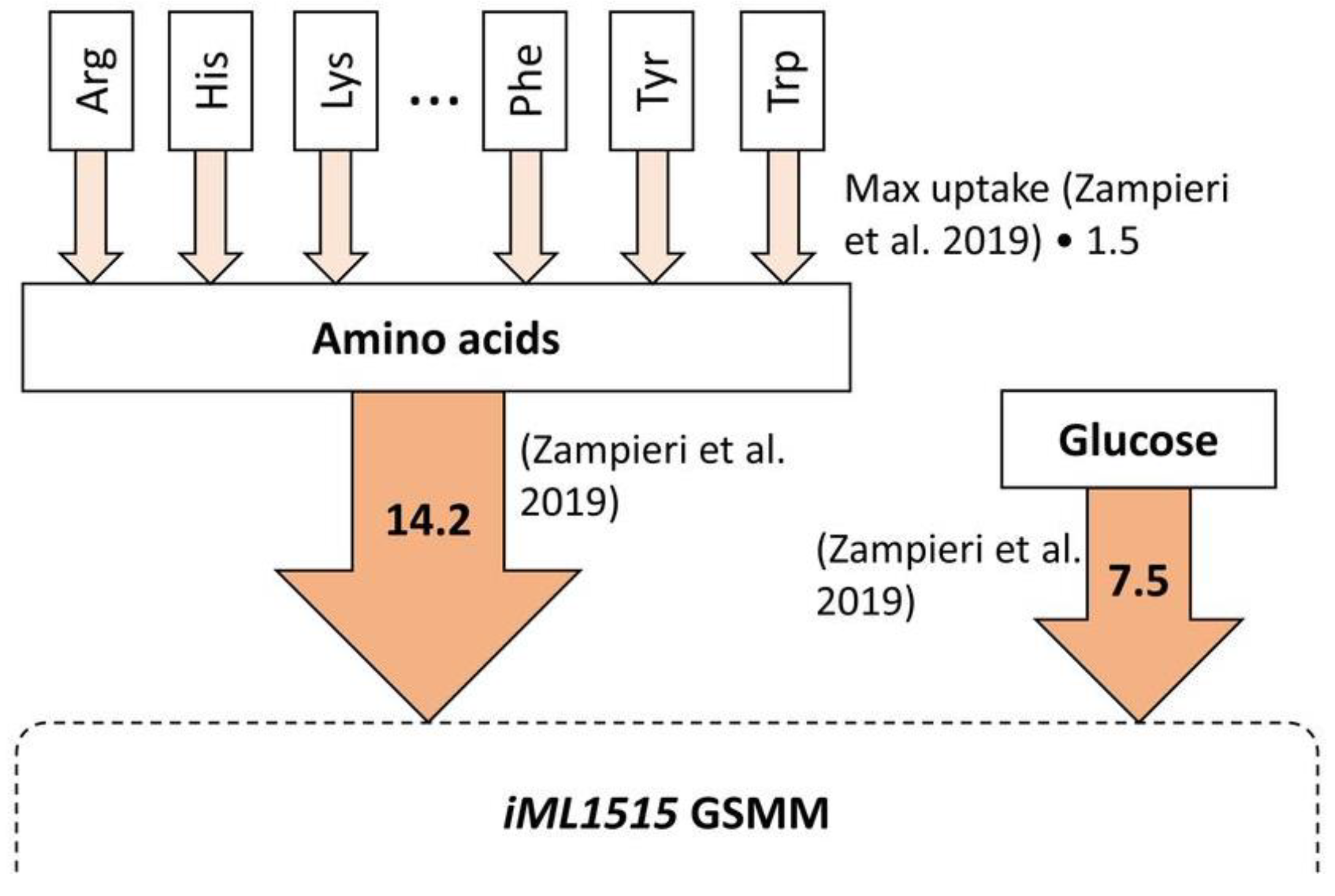
2.3. Sustainable Metabolic Engineering (SME) Task Set-Up
2.4. Environmental, Economic, and Social Impact of Biomass Extract for SME
3. Results
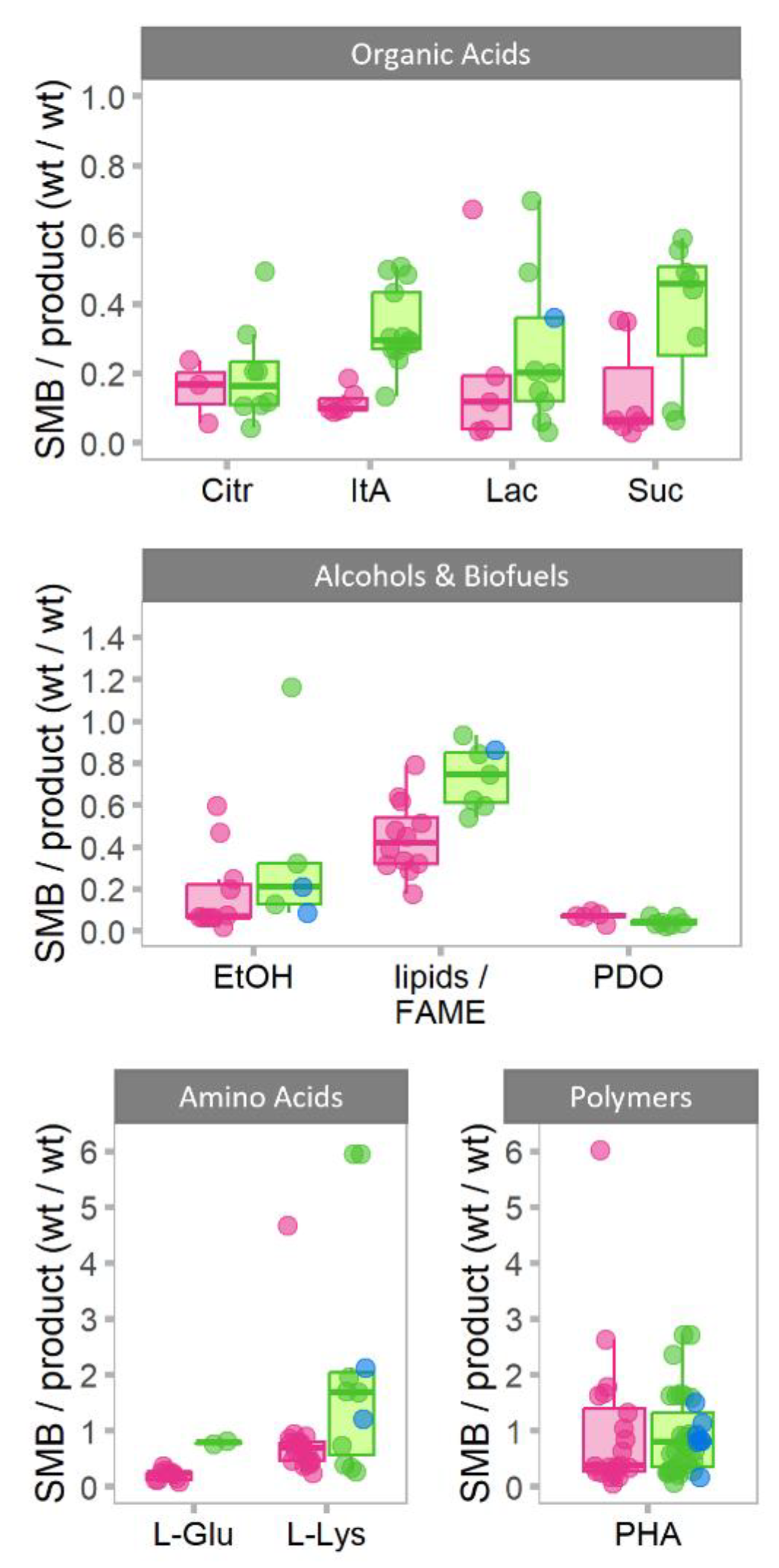
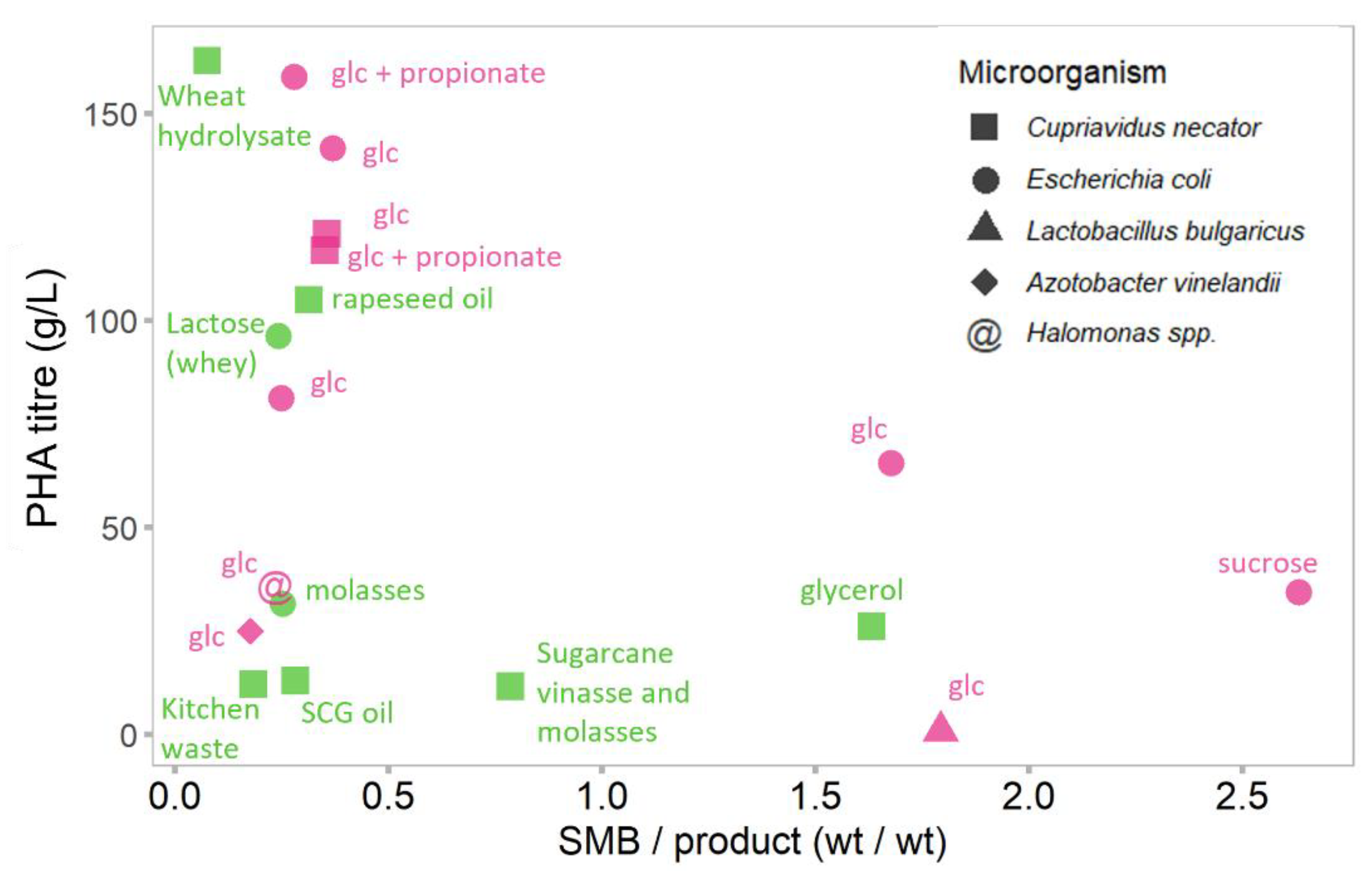
3.1. The Comparison of SMB Formation Using Purified and Waste Substrates
The Case of SMB Formation in PHA Production from Wastes and Purified Substrates
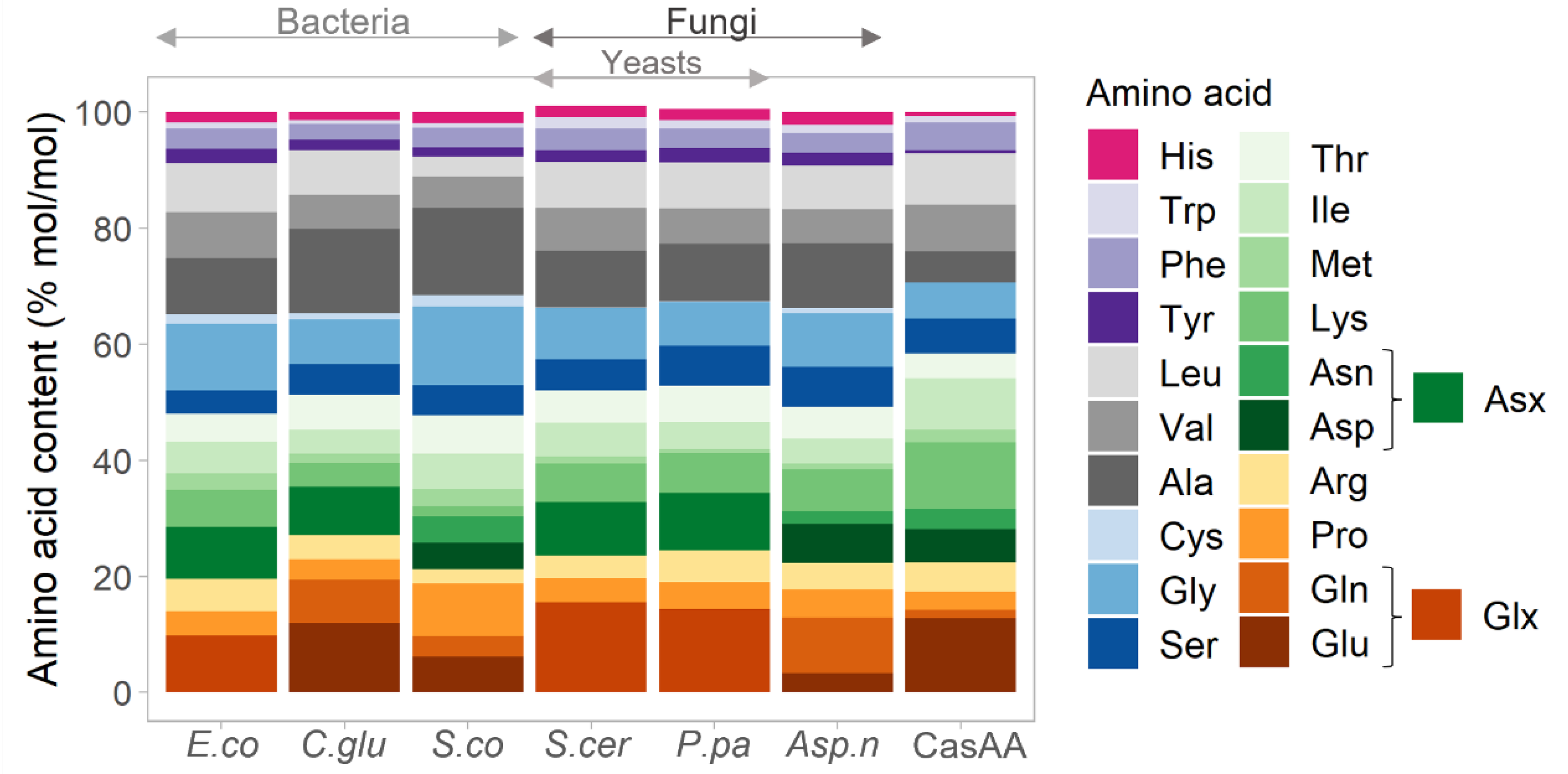
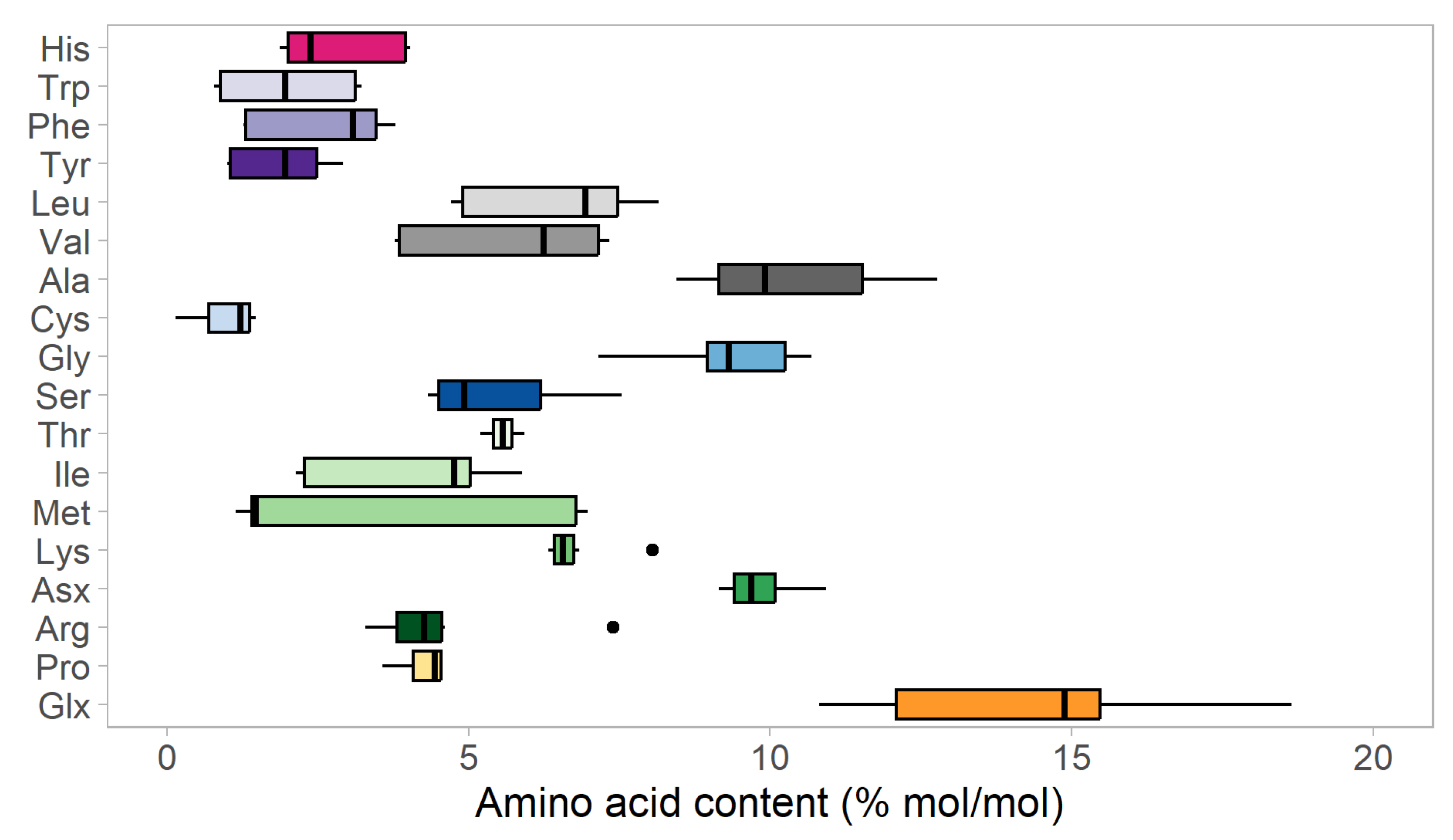
3.2. Amino Acid Composition of SMB
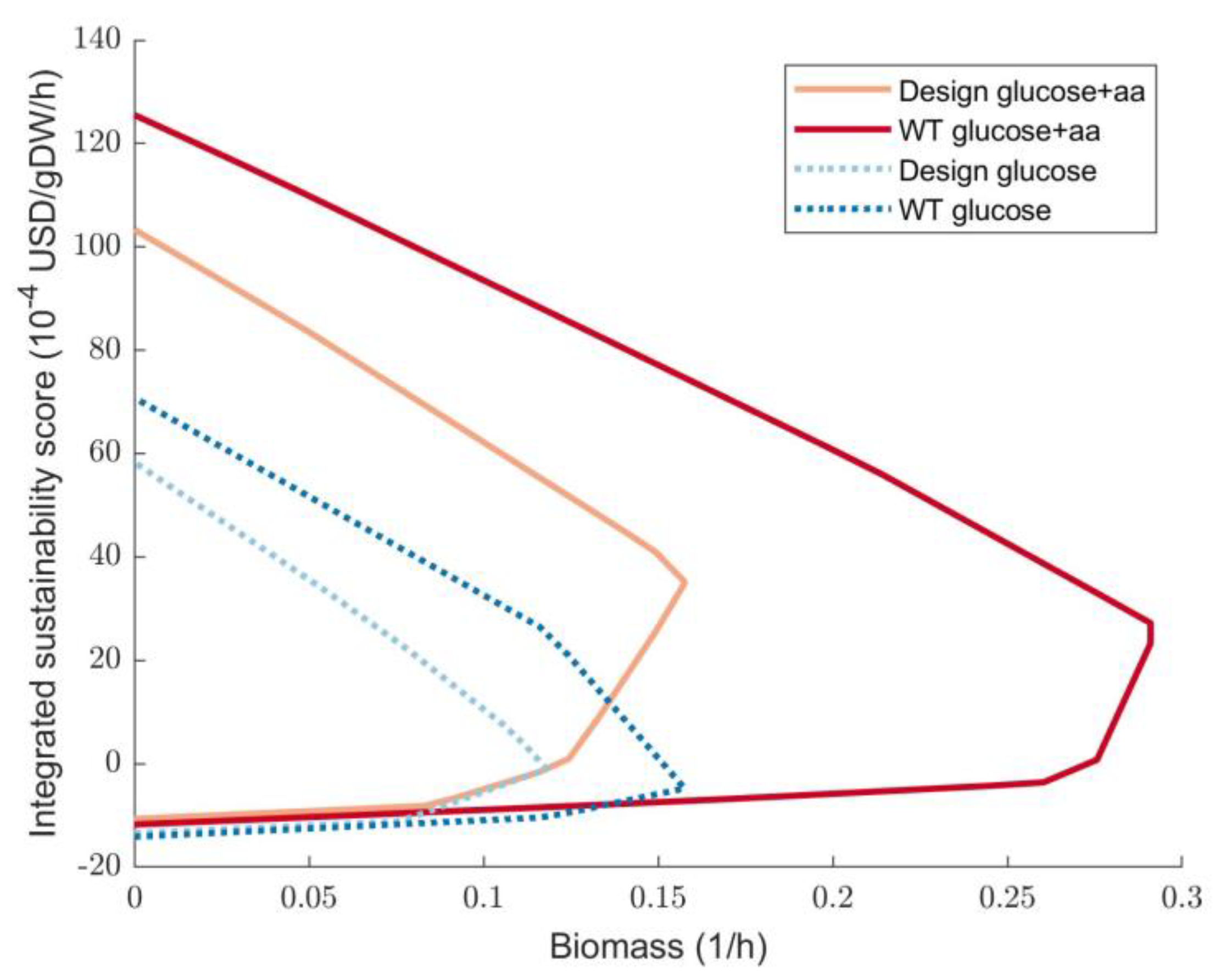
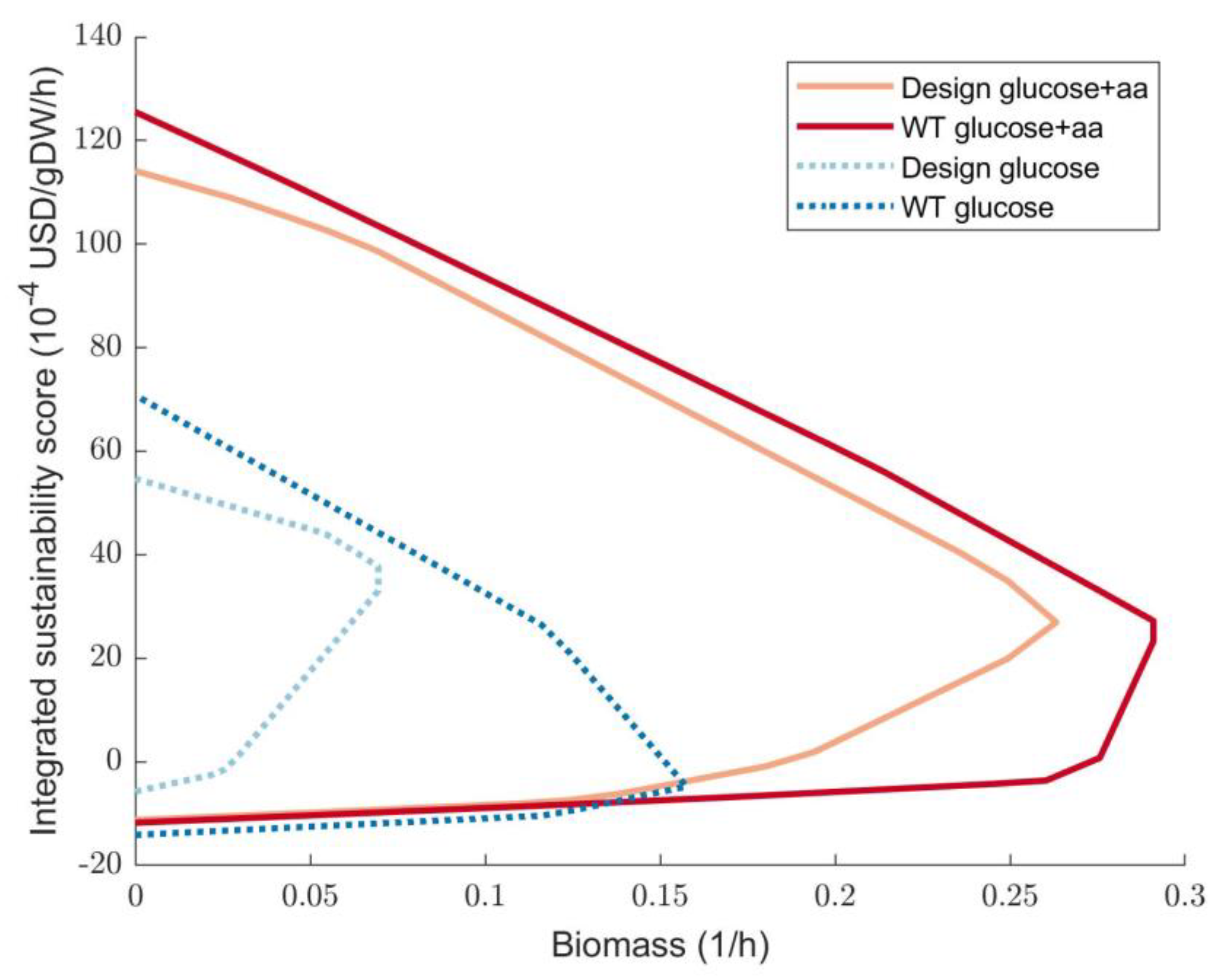
3.3. Sustainability Optimisation of SMB Re-Use as a Substrate
4. Discussion
4.1. SMB Formation from Waste Substrates
4.2. Optimizing Metabolic Designs for Sustainable SMB Use as a Substrate in Fermentation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, S.Y.; Kim, H.U.; Chae, T.U.; Cho, J.S.; Kim, J.W.; Shin, J.H.; Kim, D.I.; Ko, Y.-S.; Jang, W.D.; Jang, Y.-S. A Comprehensive Metabolic Map for Production of Bio-Based Chemicals. Nat. Catal. 2019, 2, 18–33. [Google Scholar] [CrossRef]
- Alonso, S.; Rendueles, M.; Díaz, M. Microbial Production of Specialty Organic Acids from Renewable and Waste Materials. Crit. Rev. Biotechnol. 2015, 35, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Pejó, E.; Morales-Palomo, S.; González-Fernández, C. Microbial Lipids from Organic Wastes: Outlook and Challenges. Bioresour. Technol. 2021, 323, 124612. [Google Scholar] [CrossRef] [PubMed]
- Karthick, C.; Nanthagopal, K. A Comprehensive Review on Ecological Approaches of Waste to Wealth Strategies for Production of Sustainable Biobutanol and Its Suitability in Automotive Applications. Energy Convers. Manag. 2021, 239, 114219. [Google Scholar] [CrossRef]
- Sirohi, R.; Prakash Pandey, J.; Kumar Gaur, V.; Gnansounou, E.; Sindhu, R. Critical Overview of Biomass Feedstocks as Sustainable Substrates for the Production of Polyhydroxybutyrate (PHB). Bioresour. Technol. 2020, 311, 123536. [Google Scholar] [CrossRef]
- Kircher, M. Bioeconomy—Present Status and Future Needs of Industrial Value Chains. New Biotechnol. 2021, 60, 96–104. [Google Scholar] [CrossRef]
- Alibardi, L.; Cossu, R. Effects of Carbohydrate, Protein and Lipid Content of Organic Waste on Hydrogen Production and Fermentation Products. Waste Manag. 2016, 47, 69–77. [Google Scholar] [CrossRef]
- Stikane, A.; Dace, E.; Stalidzans, E. Closing the Loop in Bioproduction: Spent Microbial Biomass as a Resource within Circular Bioeconomy. New Biotechnol. 2022, 70, 109–115. [Google Scholar] [CrossRef]
- Paramasivam, P.; Kanagesan, K.; Bhuyar, P.; Govindan, N.; Ab. Rahim, M.H.; Maniam, G.P. Biomass and Lipid Production from Indigenous Nannochloropsis sp. by Employing Stress Factors for Improved Biodiesel Production. Environ. Dev. Sustain. 2021. [Google Scholar] [CrossRef]
- Research and Markets Ltd. Global Precision Fermentation Market by Ingredient (Whey & Casein Protein, Egg White, Collagen Protein, Heme Protein), Microbe (Yeast, Algae, Fungi, Bacteria), Application (Meat & Seafood, Dairy Alternatives, Egg Alternatives), and Region—Forecast to 2030; Research and Markets: Dublin, Ireland, 2022. [Google Scholar]
- Thiele, I.; Palsson, B.Ø. A Protocol for Generating a High-Quality Genome-Scale Metabolic Reconstruction. Nat. Protoc. 2010, 5, 93–121. [Google Scholar] [CrossRef]
- Stephanopoulos, G. Metabolic Engineering. Curr. Opin. Biotechnol. 1994, 5, 196–200. [Google Scholar] [CrossRef]
- Otero, J.M.; Nielsen, J. Industrial Systems Biology. Biotechnol. Bioeng. 2010, 105, 439–460. [Google Scholar] [CrossRef]
- Stalidzans, E.; Seiman, A.; Peebo, K.; Komasilovs, V.; Pentjuss, A. Model-Based Metabolism Design: Constraints for Kinetic and Stoichiometric Models. Biochem. Soc. Trans. 2018, 46, 261–267. [Google Scholar] [CrossRef]
- Stalidzans, E.; Dace, E. Sustainable Metabolic Engineering for Sustainability Optimisation of Industrial Biotechnology. Comput. Struct. Biotechnol. J. 2021, 19, 4770–4776. [Google Scholar] [CrossRef]
- Lange, H.C.; Heijnen, J.J. Statistical Reconciliation of the Elemental and Molecular Biomass Composition of Saccharomyces Cerevisiae. Biotechnol. Bioeng. 2001, 75, 334–344. [Google Scholar] [CrossRef]
- Carnicer, M.; Baumann, K.; Töplitz, I.; Sánchez-Ferrando, F.; Mattanovich, D.; Ferrer, P.; Albiol, J. Macromolecular and Elemental Composition Analysis and Extracellular Metabolite Balances of Pichia Pastoris Growing at Different Oxygen Levels. Microb. Cell Factories 2009, 8, 65. [Google Scholar] [CrossRef]
- Ilmén, M.; Koivuranta, K.; Ruohonen, L.; Suominen, P.; Penttilä, M. Efficient Production of L-Lactic Acid from Xylose by Pichia Stipitis. Appl. Environ. Microbiol. 2007, 73, 117–123. [Google Scholar] [CrossRef]
- Orth, J.D.; Conrad, T.M.; Na, J.; Lerman, J.A.; Nam, H.; Feist, A.M.; Palsson, B.Ø. A Comprehensive Genome-Scale Reconstruction of Escherichia Coli Metabolism—2011. Mol. Syst. Biol. 2011, 7, 535. [Google Scholar] [CrossRef]
- Kjeldsen, K.R.; Nielsen, J. In Silico Genome-Scale Reconstruction and Validation of the Corynebacterium Glutamicum Metabolic Network. Biotechnol. Bioeng. 2009, 102, 583–597. [Google Scholar] [CrossRef]
- Amara, A.; Takano, E.; Breitling, R. Development and Validation of an Updated Computational Model of Streptomyces Coelicolor Primary and Secondary Metabolism. BMC Genom. 2018, 19, 519. [Google Scholar] [CrossRef]
- Upton, D.J.; McQueen-Mason, S.J.; Wood, A.J. In Silico Evolution of Aspergillus Niger Organic Acid Production Suggests Strategies for Switching Acid Output. Biotechnol. Biofuels 2020, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Albers, E.; Larsson, C.; Lidé, N.G.; Niklasson, C.; Gustafsson, L. Influence of the Nitrogen Source on Saccharomyces Cerevisiae Anaerobic Growth and Product Formation. Appl. Environ. Microbiol. 1996, 62, 3187–3195. [Google Scholar] [CrossRef] [PubMed]
- Nookaew, I.; Jewett, M.C.; Meechai, A.; Thammarongtham, C.; Laoteng, K.; Cheevadhanarak, S.; Nielsen, J.; Bhumiratana, S. The Genome-Scale Metabolic Model iIN800 of Saccharomyces Cerevisiae and Its Validation: A Scaffold to Query Lipid Metabolism. BMC Syst. Biol. 2008, 2, 71. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Li, F.; Sánchez, B.J.; Zhu, Z.; Li, G.; Domenzain, I.; Marcišauskas, S.; Anton, P.M.; Lappa, D.; Lieven, C.; et al. Cerevisiae Metabolic Model Yeast8 and Its Ecosystem for Comprehensively Probing Cellular Metabolism. Nat. Commun. 2019, 10, 3586. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 30 April 2023).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Muiznieks, R.; Dace, E.; Stalidzans, E. Sustainable Metabolic Engineering Design Development Employing Complex Objective Function. Preprints.org 2023, 2023050094. [Google Scholar] [CrossRef]
- Heirendt, L.; Arreckx, S.; Pfau, T.; Mendoza, S.N.; Richelle, A.; Heinken, A.; Haraldsdóttir, H.S.; Wachowiak, J.; Keating, S.M.; Vlasov, V.; et al. Creation and Analysis of Biochemical Constraint-Based Models Using the COBRA Toolbox v.3.0. Nat. Protoc. 2019, 14, 639–702. [Google Scholar] [CrossRef]
- Kostromins, A.; Stalidzans, E. Paint4Net: COBRA Toolbox Extension for Visualization of Stoichiometric Models of Metabolism. Biosystems 2012, 109, 233–239. [Google Scholar] [CrossRef]
- Petrovs, R.; Stalidzans, E.; Pentjuss, A. IMFLer: A Web Application for Interactive Metabolic Flux Analysis and Visualization. J. Comput. Biol. 2021, 28, 1021–1032. [Google Scholar] [CrossRef]
- Klamt, S.; Mahadevan, R. On the Feasibility of Growth-Coupled Product Synthesis in Microbial Strains. Metab. Eng. 2015, 30, 166–178. [Google Scholar] [CrossRef]
- von Kamp, A.; Klamt, S. Growth-Coupled Overproduction Is Feasible for Almost All Metabolites in Five Major Production Organisms. Nat. Commun. 2017, 8, 15956. [Google Scholar] [CrossRef]
- Motamedian, E.; Berzins, K.; Muiznieks, R.; Stalidzans, E. OptEnvelope: A Target Point Guided Method for Growth-Coupled Production Using Knockouts. 2023.03.10.532079. Available online: https://www.biorxiv.org/content/10.1101/2023.03.10.532079v1 (accessed on 29 April 2023).
- Patil, K.R.; Rocha, I.; Förster, J.; Nielsen, J. Evolutionary Programming as a Platform for in Silico Metabolic Engineering. BMC Bioinform. 2005, 6, 308. [Google Scholar] [CrossRef]
- Zampieri, M.; Hörl, M.; Hotz, F.; Müller, N.F.; Sauer, U. Regulatory Mechanisms Underlying Coordination of Amino Acid and Glucose Catabolism in Escherichia coli. Nat. Commun. 2019, 10, 3354. [Google Scholar] [CrossRef]
- King, Z.A.; Lu, J.; Dräger, A.; Miller, P.; Federowicz, S.; Lerman, J.A.; Ebrahim, A.; Palsson, B.O.; Lewis, N.E. BiGG Models: A Platform for Integrating, Standardizing and Sharing Genome-Scale Models. Nucleic Acids Res. 2016, 44, D515–D522. [Google Scholar] [CrossRef]
- Monk, J.M.; Lloyd, C.J.; Brunk, E.; Mih, N.; Sastry, A.; King, Z.; Takeuchi, R.; Nomura, W.; Zhang, Z.; Mori, H.; et al. iML1515, a Knowledgebase That Computes Escherichia Coli Traits. Nat. Biotechnol. 2017, 35, 904–908. [Google Scholar] [CrossRef]
- Misailidis, N.; Petrides, D. Yeast Extract Production—Process Modeling and Techno-Economic Assessment (TEA) Using SuperPro Designer. August 2020. preprint. Available online: https://www.researchgate.net/publication/343922223_Yeast_Extract_Production_-_Process_Modeling_and_Techno-Economic_Assessment_TEA_using_SuperPro_Designer (accessed on 30 April 2023).
- Adom, F.; Dunn, J.B. (Eds.) Material and Energy Flows in the Production of Macro and Micronutrients, Buffers, and Chemicals Used in Biochemical Processes for the Production of Fuels and Chemicals from Biomass; Energy Systems Division, Argonne National Laboratory: Argonne, IL, USA, 2015; p. 16. [Google Scholar]
- Patel, M.; Crank, M.; Dornburg, V.; Hermann, B.; Roes, A.L.; Hüsing, B.; Overbeek, L.; Terragni, F.; Recchia, E. Medium and Long-Term Opportunities and Risks of the Biotechnological Production of Bulk Chemicals from Renewable Resources—The BREW Project. Utrecht University, Utrecht, The Netherlands. 2006. Project Report. Available online: https://dspace.library.uu.nl/handle/1874/21824 (accessed on 30 April 2023).
- Philip, S.; Keshavarz, T.; Roy, I. Polyhydroxyalkanoates: Biodegradable Polymers with a Range of Applications. J. Chem. Technol. Biotechnol. 2007, 82, 233–247. [Google Scholar] [CrossRef]
- Lee, S.M.; Cho, D.-H.; Jung, H.J.; Kim, B.; Kim, S.H.; Bhatia, S.K.; Gurav, R.; Jeon, J.-M.; Yoon, J.-J.; Kim, W.; et al. Finding of Novel Polyhydroxybutyrate Producer Loktanella Sp. SM43 Capable of Balanced Utilization of Glucose and Xylose from Lignocellulosic Biomass. Int. J. Biol. Macromol. 2022, 208, 809–818. [Google Scholar] [CrossRef]
- Wang, J.; Su, Y.; Jia, F.; Jin, H. Characterization of Casein Hydrolysates Derived from Enzymatic Hydrolysis. Chem. Cent. J. 2013, 7, 62. [Google Scholar] [CrossRef]
- Burkovski, A.; Krämer, R. Bacterial Amino Acid Transport Proteins: Occurrence, Functions, and Significance for Biotechnological Applications. Appl. Microbiol. Biotechnol. 2002, 58, 265–274. [Google Scholar] [CrossRef]
- Bianchi, F.; van’t Klooster, J.S.; Ruiz, S.J.; Poolman, B. Regulation of Amino Acid Transport in Saccharomyces Cerevisiae. Microbiol. Mol. Biol. Rev. 2019, 83, e00024-19. [Google Scholar] [CrossRef]
- Brown, C.J.; Todd, K.M.; Rosenzweig, R.F. Multiple Duplications of Yeast Hexose Transport Genes in Response to Selection in a Glucose-Limited Environment. Mol. Biol. Evol. 1998, 15, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Gontard, N.; Sonesson, U.; Birkved, M.; Majone, M.; Bolzonella, D.; Celli, A.; Angellier-Coussy, H.; Jang, G.W.; Verniquet, A.; Broeze, J.; et al. A Research Challenge Vision Regarding Management of Agricultural Waste in a Circular Bio-Based Economy. Crit. Rev. Environ. Sci. Technol. 2018, 48, 614–654. [Google Scholar] [CrossRef]
- Comelli, R.N.; Seluy, L.G.; Benzzo, M.T.; Isla, M.A. Combined Utilization of Agro-Industrial Wastewaters for Non-Lignocellulosic Second-Generation Bioethanol Production. Waste Biomass Valorization 2020, 11, 265–275. [Google Scholar] [CrossRef]
- Jaeger, A.; Arendt, E.K.; Zannini, E.; Sahin, A.W. Brewer’s Spent Yeast (BSY), an Underutilized Brewing By-Product. Fermentation 2020, 6, 123. [Google Scholar] [CrossRef]
- Wernick, D.G.; Liao, J.C. Protein-Based Biorefining: Metabolic Engineering for Production of Chemicals and Fuel with Regeneration of Nitrogen Fertilizers. Appl. Microbiol. Biotechnol. 2013, 97, 1397–1406. [Google Scholar] [CrossRef]
- Sánchez, B.J.; Zhang, C.; Nilsson, A.; Lahtvee, P.J.; Kerkhoven, E.J.; Nielsen, J. Improving the Phenotype Predictions of a Yeast Genome-Scale Metabolic Model by Incorporating Enzymatic Constraints. Mol. Syst. Biol. 2017, 13, 935. [Google Scholar] [CrossRef]
- Domenzain, I.; Sánchez, B.; Anton, M.; Kerkhoven, E.J.; Millán-Oropeza, A.; Henry, C.; Siewers, V.; Morrissey, J.P.; Sonnenschein, N.; Nielsen, J. Reconstruction of a Catalogue of Genome-Scale Metabolic Models with Enzymatic Constraints Using GECKO 2.0. Nat. Commun. 2022, 13, 3766. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Design | #1 | #7776 | Wild-Type |
|---|---|---|---|
| Fitness score of solution envelope | −59.07 | −53.49 | −47.39 |
| Main product | Succinate | Succinate | Succinate |
| Maximum growth rate (1/h) | 0.158 | 0.135 | 0.291 |
| Main production flux (mmol/gDW/h) | 7.98 | 8.01 | 6.79 |
| Glucose flux (mmol/gDW/h) | −7.50 | −7.50 | −7.50 |
| Number of knockouts | 5 | 6 | 0 |
| Minimum ISS at maximum GR (1 × 10−4 USD/gDW/h) | 34.90 | 29.26 | 23.27 |
| Minimum ISS at maximum GR (1 × 10−4 USD/gDW/h) | −10.68 | −10.35 | −11.77 |
| Environmental SS at maximum GR (1 × 10−4 USD/gDW/h) | −2.80 | −2.73 | −2.39 |
| Economic SS at maximum GR (1 × 10−4 USD/gDW/h) | 27.72 | 23.53 | 17.60 |
| Social SS at maximum GR (1 × 10−4 USD/gDW/h) | 9.98 | 8.47 | 8.05 |
| Gene knockouts | b3844, b0010, b4115, b3927, b1380 | b2502, b3844, b2905, b0010, b0114, b1380 |
| Exchange Reaction | Reaction Sustainability Coefficient (1 × 10−4 USD/mmol) | Reaction Flux (mmol/gDW/h) | Integrated Sustainability Score (1 × 10−4 USD/gDW) | Economic Sustainability Score (1 × 10−4 USD/gDW) | Environmental Sustainability Score (1 × 10−4 USD/gDW) | Social Sustainability Score (1 × 10−4 USD/gDW) |
|---|---|---|---|---|---|---|
| CO2 | −0.027 | 24.254 | −0.662 | 0.000 | −0.662 | 0.000 |
| Succinate | 3.950 | 7.976 | 31.507 | 26.795 | −2.918 | 7.630 |
| Glucose | 1.275 | −7.500 | −9.562 | −7.817 | 0.000 | −1.746 |
| SO42− | 0.288 | −0.013 | −0.004 | −0.003 | 0.000 | −0.001 |
| NH4+ | 0.020 | 13.094 | 0.265 | 0.217 | 0.000 | 0.048 |
| Ethanol | 0.547 | 18.928 | 10.347 | 7.229 | 0.824 | 2.294 |
| Biomass | 19.239 | 0.158 | 3.031 | 1.308 | −0.043 | 1.766 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stikane, A.; Baumanis, M.R.; Muiznieks, R.; Stalidzans, E. Impact of Waste as a Substrate on Biomass Formation, and Optimization of Spent Microbial Biomass Re-Use by Sustainable Metabolic Engineering. Fermentation 2023, 9, 531. https://doi.org/10.3390/fermentation9060531
Stikane A, Baumanis MR, Muiznieks R, Stalidzans E. Impact of Waste as a Substrate on Biomass Formation, and Optimization of Spent Microbial Biomass Re-Use by Sustainable Metabolic Engineering. Fermentation. 2023; 9(6):531. https://doi.org/10.3390/fermentation9060531
Chicago/Turabian StyleStikane, Anna, Matiss Ricards Baumanis, Reinis Muiznieks, and Egils Stalidzans. 2023. "Impact of Waste as a Substrate on Biomass Formation, and Optimization of Spent Microbial Biomass Re-Use by Sustainable Metabolic Engineering" Fermentation 9, no. 6: 531. https://doi.org/10.3390/fermentation9060531
APA StyleStikane, A., Baumanis, M. R., Muiznieks, R., & Stalidzans, E. (2023). Impact of Waste as a Substrate on Biomass Formation, and Optimization of Spent Microbial Biomass Re-Use by Sustainable Metabolic Engineering. Fermentation, 9(6), 531. https://doi.org/10.3390/fermentation9060531