
Algal Hydrogen Production and Exopolysaccharide Patterns in Chlorella–Bacillus Inter-Kingdom Co-Cultures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Growth of Axenic Algal Strains and Pure Bacterial Strains
2.2. Algal and Bacterial Co-Cultures in Hypo-Vial Bottles
2.3. Gas Phase Analysis
2.4. Morphological Studies
2.5. Chlorophyll and Carbohydrates Measurements
3. Results
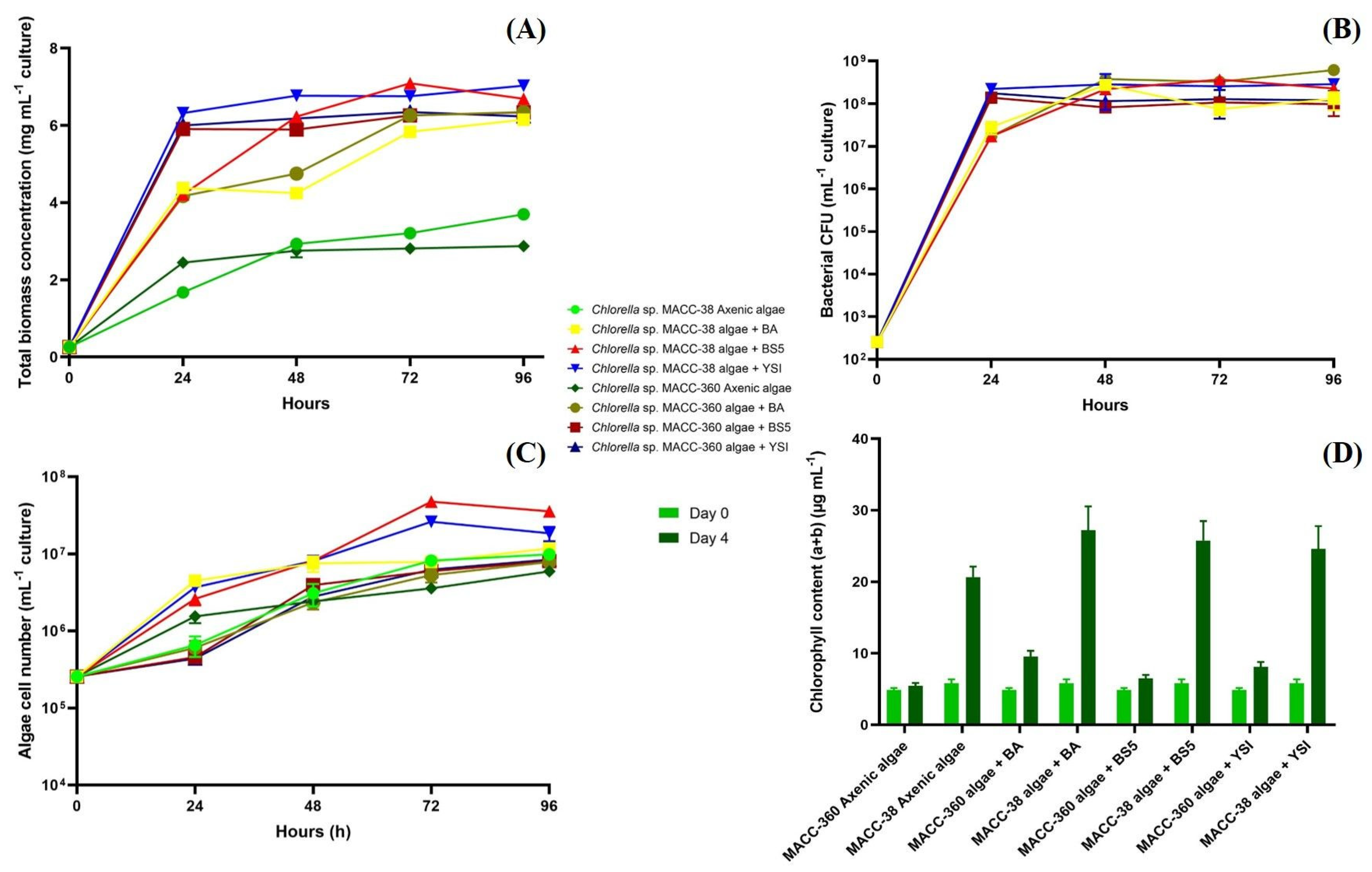
3.1. Algal Biomass Yield Is Facilitated by Bacterial Partners
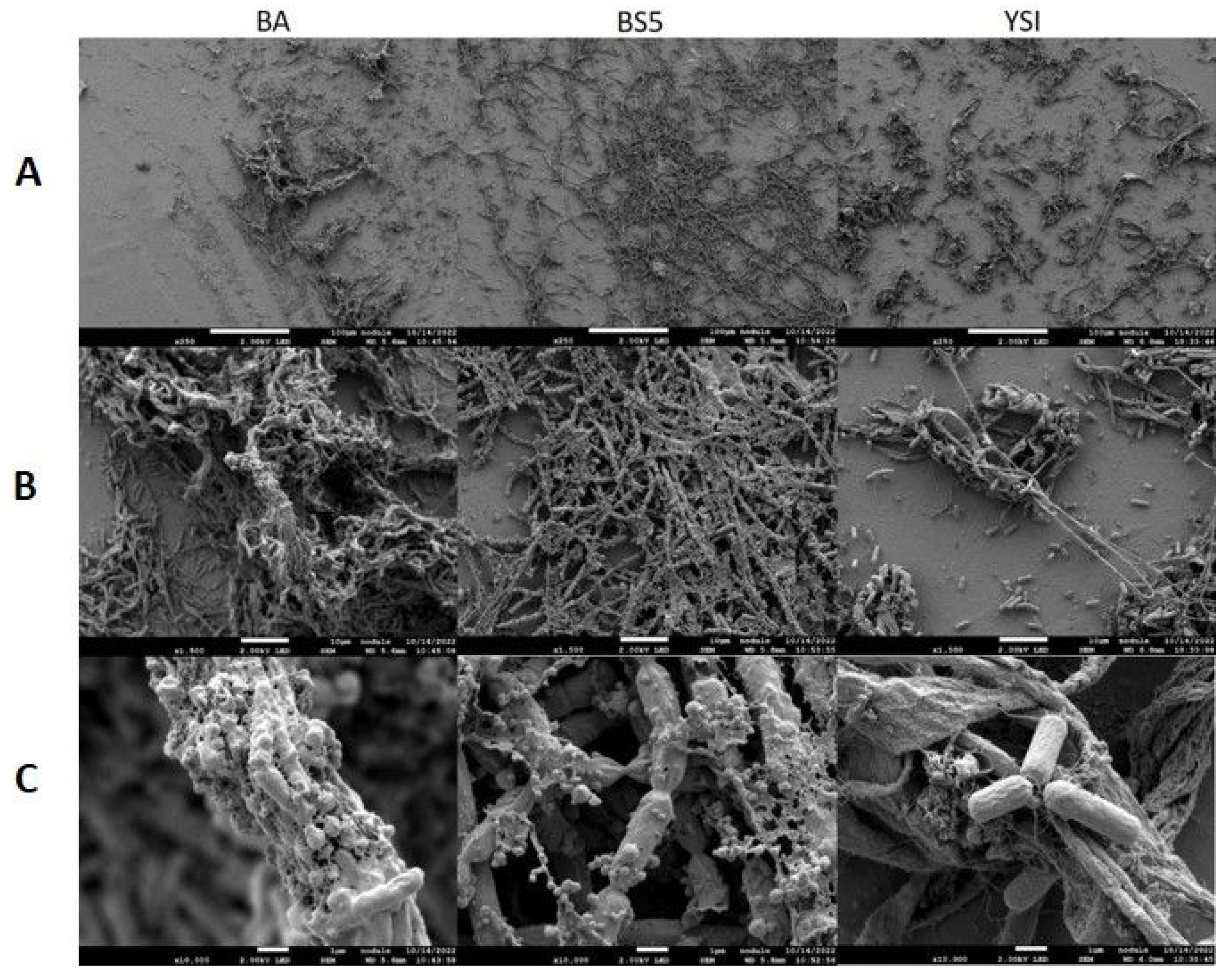
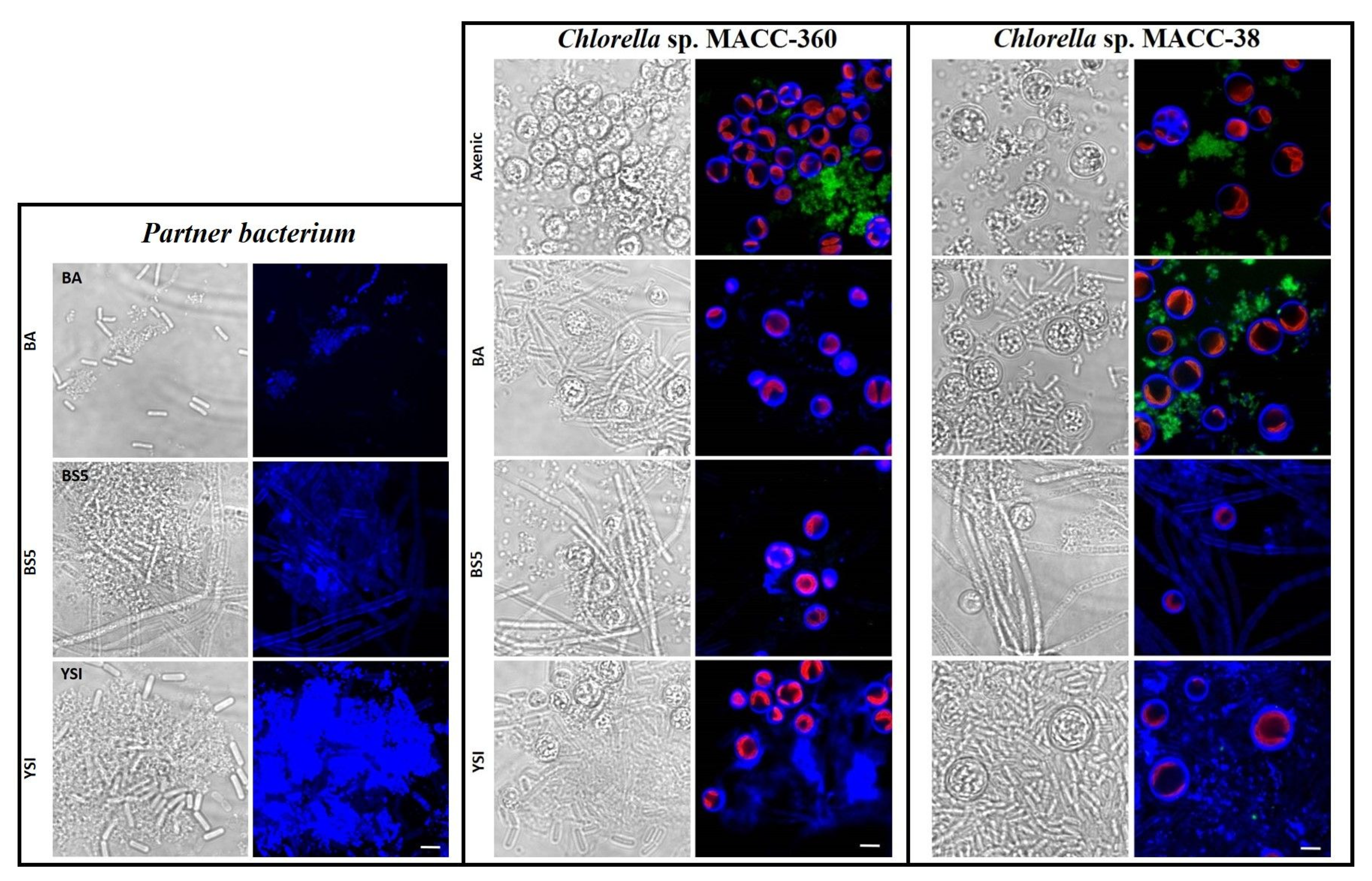
3.2. Co-Culture-Specific EPS Patterns
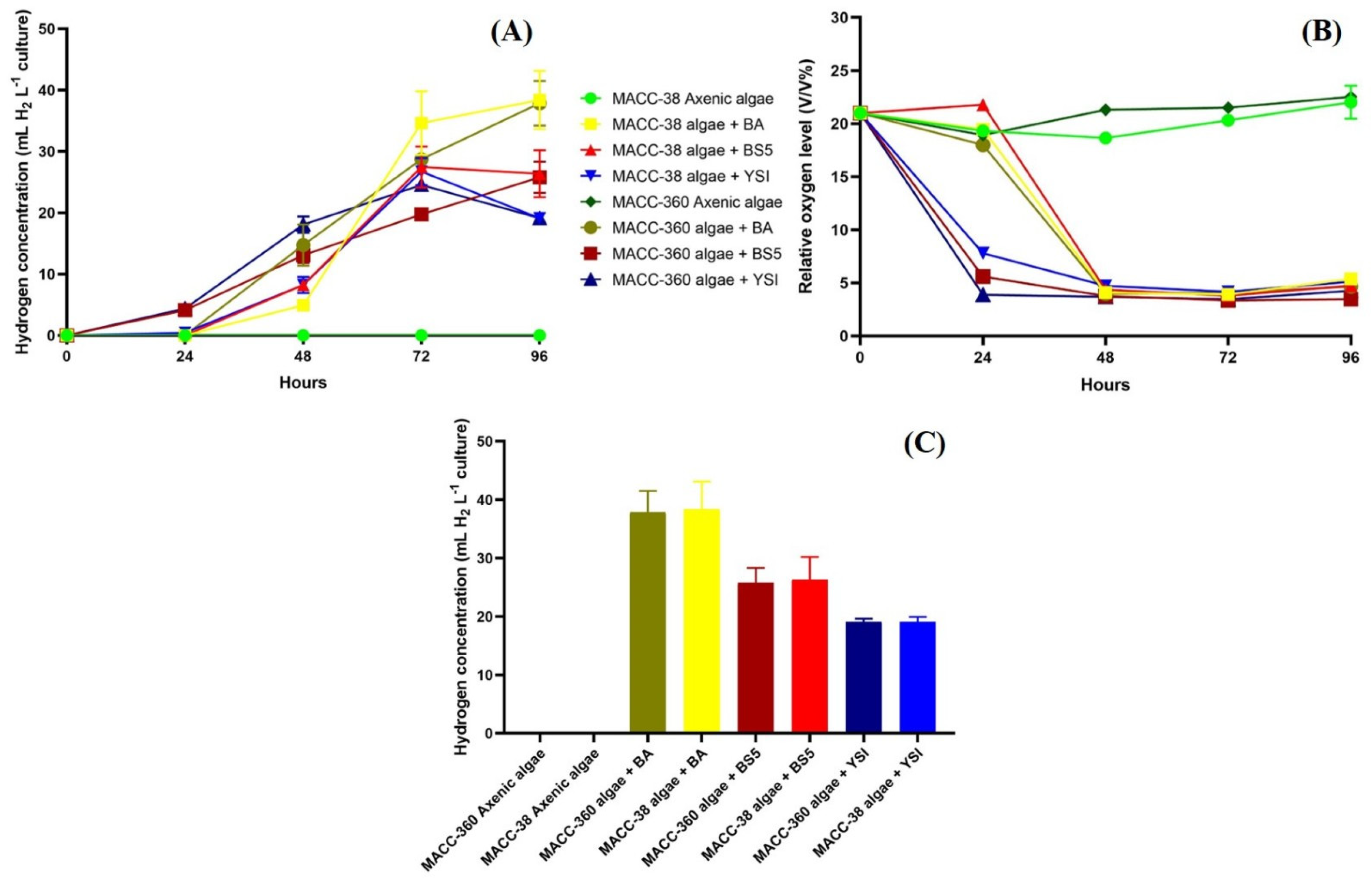
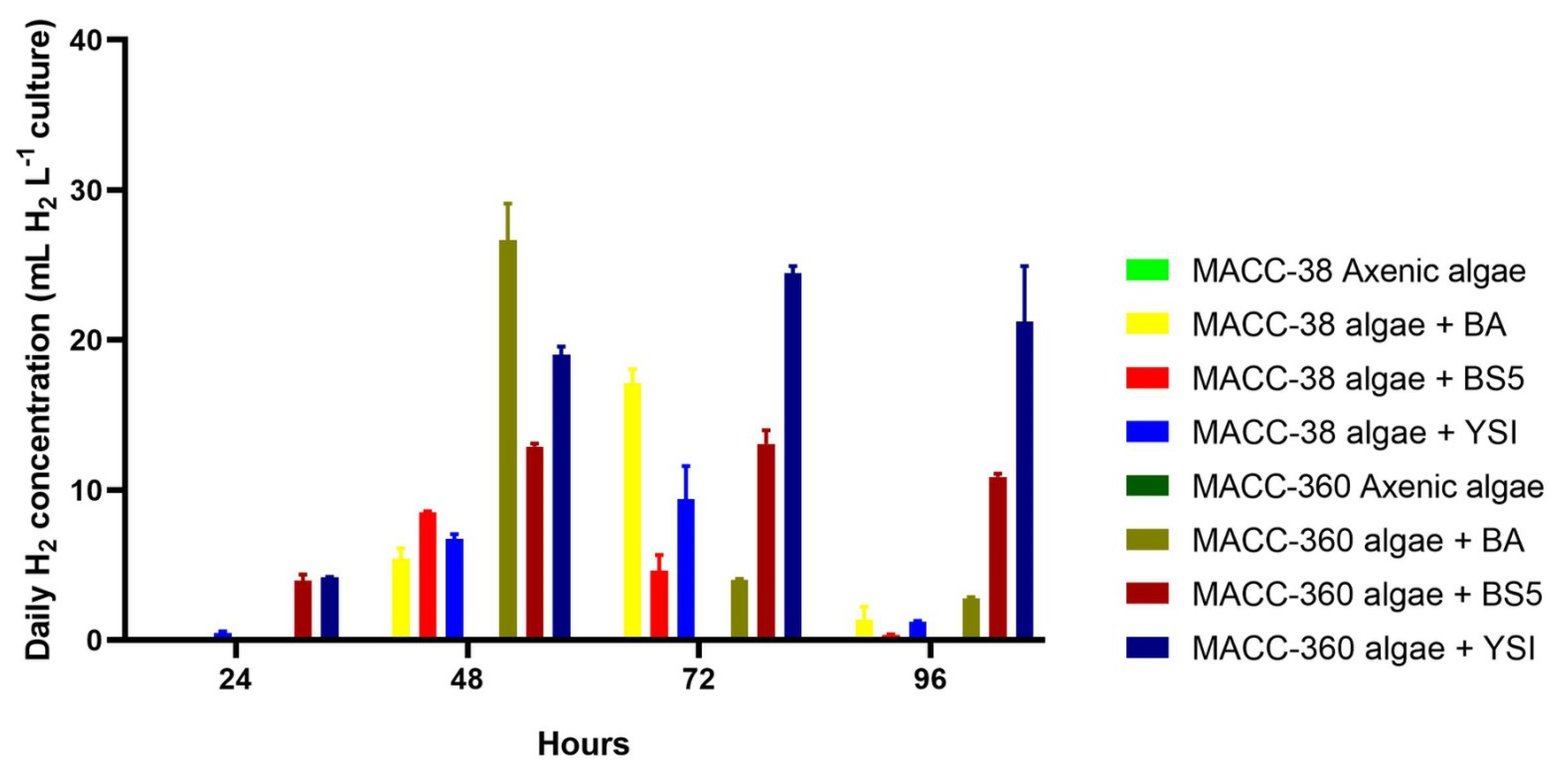
3.3. Algal Hydrogen Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Demirbas, A.; Demirbas, F.M. Importance of Algae Oil as a Source of Biodiesel. Energy Convers. Manag. 2011, 52, 163–170. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, H.; Zhang, X.; Han, F.; Tu, W.; Yang, W. Microalgal Hydrogen Production. Small Methods 2020, 4, 1900514. [Google Scholar] [CrossRef]
- Wang, J.; Yin, Y. Fermentative Hydrogen Production Using Pretreated Microalgal Biomass as Feedstock. Microb. Cell Fact. 2018, 17, 22. [Google Scholar] [CrossRef] [PubMed]
- Redding, K.E.; Appel, J.; Boehm, M.; Schuhmann, W.; Nowaczyk, M.M.; Yacoby, I.; Gutekunst, K. Advances and Challenges in Photosynthetic Hydrogen Production. Trends Biotechnol. 2022, 40, 1313–1325. [Google Scholar] [CrossRef] [PubMed]
- Ghirardi, M.L.; Dubini, A.; Yu, J.; Maness, P.C. Photobiological Hydrogen-Producing Systems. Chem. Soc. Rev. 2009, 38, 52–61. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Thavasi, V.; Kreslavski, V.D.; Zharmukhamedov, S.K.; Klimov, V.V.; Ramakrishna, S.; Los, D.A.; Mimuro, M.; Nishihara, H.; Carpentier, R. Photosynthetic Hydrogen Production. J. Photochem. Photobiol. C Photochem. Rev. 2010, 11, 101–113. [Google Scholar] [CrossRef]
- Fan, X.; Wang, H.; Guo, R.; Yang, D.; Zhang, Y.; Yuan, X.; Qiu, Y.; Yang, Z.; Zhao, X. Comparative Study of the Oxygen Tolerance of Chlorella Pyrenoidosa and Chlamydomonas Reinhardtii CC124 in Photobiological Hydrogen Production. Algal Res. 2016, 16, 240–244. [Google Scholar] [CrossRef]
- Kruse, O.; Hankamer, B. Microalgal Hydrogen Production. Curr. Opin. Biotechnol. 2010, 21, 238–243. [Google Scholar] [CrossRef]
- Lubitz, W.; Ogata, H.; Rüdiger, O.; Reijerse, E. Hydrogenases. Chem. Rev. 2014, 114, 4081–4148. [Google Scholar] [CrossRef]
- Lakatos, G.; Deák, Z.; Vass, I.; Rétfalvi, T.; Rozgonyi, S.; Rákhely, G.; Ördög, V.; Kondorosi, É.; Maróti, G. Bacterial Symbionts Enhance Photo-Fermentative Hydrogen Evolution of Chlamydomonas Algae. Green Chem. 2014, 16, 4716–4727. [Google Scholar] [CrossRef]
- Kargi, F.; Pamukoglu, M.Y. Dark Fermentation of Ground Wheat Starch for Bio-Hydrogen Production by Fed-Batch Operation. Int. J. Hydrogen Energy 2009, 34, 2940–2946. [Google Scholar] [CrossRef]
- Akano, T.; Miura, Y.; Fukatsu, K.; Mlyasaka, H.; Ikuta, Y.; Matsumoto, H.; Hamasaki, A.; Shioji, N.; Mlzoguchi, T.; Yagi, K.; et al. Hydrogen Production by Photosynthetic Microorganisms. Appl. Biochem. Biotechnol. Part A Enzym. Eng. Biotechnol. 1996, 57–58, 677–688. [Google Scholar] [CrossRef]
- Masojídek, J.; Torzillo, G.; Koblízek, M. Photosynthesis in Microalgae. In Handbook of Microalgal Culture: Applied Phycology and Biotechnology, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 21–36. [Google Scholar] [CrossRef]
- Khoeyi, Z.A.; Seyfabadi, J.; Ramezanpour, Z. Effect of Light Intensity and Photoperiod on Biomass and Fatty Acid Composition of the Microalgae, Chlorella vulgaris. Aquac. Int. 2012, 20, 41–49. [Google Scholar] [CrossRef]
- Wirth, R.; Lakatos, G.; Böjti, T.; Maróti, G.; Bagi, Z. Anaerobic gaseous biofuel production using microalgal biomass—A review. Anaerobe 2018, 52, 1–8. [Google Scholar] [CrossRef]
- Wirth, R.; Lakatos, G.; Maróti, G.; Bagi, Z.; Minárovics, J.; Nagy, K.; Kondorosi, É.; Rákhely, G.; Kovács, K.L. Exploitation of Algal-Bacterial Associations in a Two-Stage Biohydrogen and Biogas Generation Process. Biotechnol. Biofuels. 2015, 8, 59. [Google Scholar] [CrossRef]
- Lakatos, G.; Balogh, D.; Farkas, A.; Ördög, V.; Tamás, P.; Bíró, T.; Maróti, G. Factors in Fl Uencing Algal Photobiohydrogen Production in Algal-Bacterial Co-Cultures. Algal Res. 2017, 28, 161–171. [Google Scholar] [CrossRef]
- Gorman, D.S.; Levine, R.P. Cytochrome < if and Plastocyanin: Their Sequence in the Photosynthetic Electron Transport Chain of Chlamydomonas Reinhardi. Proc. Natl. Acad. Sci. USA 1965, 54, 1665–1669. [Google Scholar] [CrossRef]
- El-Naggar, N.E.A.; Hussein, M.H.; Shaaban-Dessuuki, S.A.; Dalal, S.R. Production, Extraction and Characterization of Chlorella Vulgaris Soluble Polysaccharides and Their Applications in AgNPs Biosynthesis and Biostimulation of Plant Growth. Sci. Rep. 2020, 10, 3011. [Google Scholar] [CrossRef]
- Hupp, B.; Pap, B.; Farkas, A.; Maróti, G. Development of a Microalgae-Based Continuous Starch-to-Hydrogen Conversion Approach. Fermentation 2022, 8, 294. [Google Scholar] [CrossRef]
- Sharma, R. Effects of Culture Conditions on Growth and Biochemical Profile of Chlorella vulgaris. J. Plant Pathol. Microbiol. 2012, 3, 1–6. [Google Scholar] [CrossRef]
- Yuan, Q.; Li, H.; Wei, Z.; Lv, K.; Gao, C.; Liu, Y.; Zhao, L. Isolation, Structures and Biological Activities of Polysaccharides from Chlorella: A Review. Int. J. Biol. Macromol. 2020, 163, 2199–2209. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, M.; Bajguz, A.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Uncovering Potential Applications of Cyanobacteria and Algal Metabolites in Biology, Agriculture and Medicine: Current Status and Future Prospects. Front. Microbiol. 2017, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing Sustainability by Improving Plant Salt Tolerance through Macro-and Micro-Algal Biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; He, X.; Mao, W.; Cao, S.; Qin, L.; He, M.; He, X.; Mao, W. Anticoagulant Properties of a Green Algal Rhamnan-Type Sulfated Polysaccharide and Its Low-Molecular-Weight Fragments Prepared by Mild Acid Degradation. Mar. Drugs 2018, 16, 445. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, L.; Ren, Y.; Chen, F. Characterization of Exopolysaccharides Produced by Microalgae with Antitumor Activity on Human Colon Cancer Cells. Int. J. Biol. Macromol. 2019, 128, 761–767. [Google Scholar] [CrossRef]
- Mousavian, Z.; Safavi, M.; Azizmohseni, F.; Hadizadeh, M.; Mirdamadi, S. Characterization, Antioxidant and Anticoagulant Properties of Exopolysaccharide from Marine Microalgae. AMB Express 2022, 12, 27. [Google Scholar] [CrossRef]
- Dubini, A.; Ghirardi, M.L. Engineering Photosynthetic Organisms for the Production of Biohydrogen. Photosynth. Res. 2015, 123, 241–253. [Google Scholar] [CrossRef]
- Boboescu, I.Z.; Gherman, V.D.; Lakatos, G.; Pap, B.; Bíró, T.; Maróti, G. Surpassing the Current Limitations of Biohydrogen Production Systems: The Case for a Novel Hybrid Approach. Bioresour. Technol. 2016, 204, 192–201. [Google Scholar] [CrossRef]
- Xiao, R.; Zheng, Y. Overview of Microalgal Extracellular Polymeric Substances (EPS) and Their Applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef]
- Zhang, B.; Li, W.; Guo, Y.; Zhang, Z.; Shi, W.; Cui, F.; Lens, P.N.L.; Tay, J.H. Microalgal-Bacterial Consortia: From Interspecies Interactions to Biotechnological Applications. Renew. Sustain. Energy Rev. 2020, 118, 109563. [Google Scholar] [CrossRef]
- Buchan, A.; LeCleir, G.R.; Gulvik, C.A.; González, J.M. Master Recyclers: Features and Functions of Bacteria Associated with Phytoplankton Blooms. Nat. Rev. Microbiol. 2014, 12, 686–698. [Google Scholar] [CrossRef]
- Yao, S.; Lyu, S.; An, Y.; Lu, J.; Gjermansen, C.; Schramm, A. Microalgae–Bacteria Symbiosis in Microalgal Growth and Biofuel Production: A Review. J. Appl. Microbiol. 2019, 126, 359–368. [Google Scholar] [CrossRef]
- Fakhimi, N.; Dubini, A.; Tavakoli, O.; González-Ballester, D. Acetic Acid Is Key for Synergetic Hydrogen Production in Chlamydomonas-Bacteria Co-Cultures. Bioresour. Technol. 2019, 289, 121648. [Google Scholar] [CrossRef]
- Prakash, J.; Sharma, R.; Patel, S.K.S.; Kim, I.W.; Kalia, V.C. Bio-Hydrogen Production by Co-Digestion of Domestic Wastewater and Biodiesel Industry Effluent. PLoS ONE. 2018, 13, e0199059. [Google Scholar] [CrossRef]
- Mazareli, R.C.d.S.; Sakamoto, I.K.; Silva, E.L.; Varesche, M.B.A. Bacillus sp. Isolated from Banana Waste and Analysis of Metabolic Pathways in Acidogenic Systems in Hydrogen Production. J. Environ. Manag. 2019, 247, 178–186. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying Mechanics of Plant Growth Promoting Rhizobacteria (PGPR): A Review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.R.; Reva, O.; et al. Comparative Analysis of the Complete Genome Sequence of the Plant Growth-Promoting Bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar] [CrossRef]
- Choi, S.K.; Jeong, H.; Kloepper, J.W.; Ryu, C.M. Genome Sequence of Bacillus Amyloliquefaciens GB03, an Active Ingredient of the First Commercial Biological Control Product. Genome Announc. 2014, 2, e01092-14. [Google Scholar] [CrossRef]
- Liu, H.; Prajapati, V.; Prajapati, S.; Bais, H.; Lu, J. Comparative Genome Analysis of Bacillus amyloliquefaciens Focusing on Phylogenomics, Functional Traits, and Prevalence of Antimicrobial and Virulence Genes. Front. Genet. 2021, 12, 724217. [Google Scholar] [CrossRef]
- Di Franco, C.; Beccari, E.; Santini, T.; Pisaneschi, G.; Tecce, G. Colony Shape as a Genetic Trait in the Pattern-Forming Bacillus Mycoides. BMC Microbiol. 2002, 2, 33. [Google Scholar] [CrossRef]
- Bottone, E.J. Bacillus Cereus, a Volatile Human Pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef] [PubMed]
- Vilain, S.; Luo, Y.; Hildreth, M.B.; Brözel, V.S. Analysis of the Life Cycle of the Soil Saprophyte Bacillus cereus in Liquid Soil Extract and in Soil. Appl. Environ. Microbiol. 2006, 72, 4970–4977. [Google Scholar] [CrossRef] [PubMed]
- Toole, G.O.; Kaplan, H.B.; Kolter, R. Biofillm Formation as Microbial Development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef] [PubMed]
- Rani, V.; Maróti, G. Light-Dependent Nitrate Removal Capacity of Green Microalgae. Int. J. Mol. Sci. 2022, 24, 77. [Google Scholar] [CrossRef] [PubMed]
- Lichtenhaler, H.K.; Wellburn, A.R. Determinations of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Gitau, M.M.; Farkas, A.; Balla, B.; Ördög, V.; Futó, Z.; Maróti, G. Strain-Specific Biostimulant Effects of Chlorella and Chlamydomonas Green Microalgae on Medicago truncatula. Plants 2021, 10, 1060. [Google Scholar] [CrossRef]
- Fakhimi, N.; Gonzalez-Ballester, D.; Fernández, E.; Galván, A.; Dubini, A. Algae-Bacteria Consortia as a Strategy to Enhance H2 Production. Cells 2020, 9, 1353. [Google Scholar] [CrossRef]
- Lobos, J.; Wisniewski, C.; Heran, M.; Grasmick, A. Membrane Bioreactor Performances: Effluent Quality Ofcontinuous and Sequencing Systems for Water Reuse. Desalination 2007, 204, 39–45. [Google Scholar] [CrossRef]
- Orhon, D.; Artan, N. Nutrient Removal Performance of a Five-Step Sequencing Batch Reactor as a Function of Wastewater Composition. Process Biochem. 2006, 41, 216–220. [Google Scholar] [CrossRef]
- Wang, C.; Li, J.; Wang, B.; Zhang, G. Development of an Empirical Model for Domestic Wastewater Treatment by Biological Aerated Filter. Process Biochem. 2006, 41, 778–782. [Google Scholar] [CrossRef]
- El Moussaoui, T.; Kessraoui, A.; Ouazzani, N.; Seffen, M.; Mandi, L. Synthetic Urban Wastewater Treatment by an Activated Sludge Reactor: Evolution of Bacterial Biomass and Purifying Efficiency. J. Mater. Environ. Sci. 2018, 9, 817–827. [Google Scholar]
- Yerushalmi, L.; Alimahmoodi, M.; Mulligan, C.N. Treatment of Synthetic Wastewater and Hog Waste with Reduced Sludge Generation by the Multi-Environment BioCAST Technology. Water Sci. Technol. 2013, 67, 587–593. [Google Scholar] [CrossRef]
- Wirth, R.; Pap, B.; Böjti, T.; Shetty, P.; Lakatos, G.; Bagi, Z.; Kovács, K.L.; Maróti, G. Chlorella vulgaris and Its Phycosphere in Wastewater: Microalgae-Bacteria Interactions During Nutrient Removal. Front. Bioeng. Biotechnol. 2020, 8, 557572. [Google Scholar] [CrossRef]
- Rani, V.; Maróti, G. Assessment of Nitrate Removal Capacity of Two Selected Eukaryotic Green Microalgae. Cells 2021, 10, 2490. [Google Scholar] [CrossRef]
- Grossman, A.R.; Catalanotti, C.; Yang, W.; Dubini, A.; Magneschi, L.; Subramanian, V.; Posewitz, M.C.; Seibert, M. Multiple Facets of Anoxic Metabolism and Hydrogen Production in the Unicellular Green Alga Chlamydomonas reinhardtii. New Phytol. 2011, 190, 279–288. [Google Scholar] [CrossRef]
- Ghirardi, M.L.; King, P.; Kosourov, S.; Forestier, M.; Zhang, L.; Seibert, M. Development of Algal Systems for Hydrogen Photoproduction: Addressing the Hydrogenase Oxygen-Sensitivity Problem. In Artificial Photosynthesis; Wiley-VCH: Weinheim, Germany, 2006; pp. 211–227. [Google Scholar] [CrossRef]
- Noth, J.; Krawietz, D.; Hemschemeier, A.; Happe, T. Pyruvate:Ferredoxin Oxidoreductase Is Coupled to Light-Independent Hydrogen Production in Chlamydomonas reinhardtii. J. Biol. Chem. 2013, 288, 4368–4377. [Google Scholar] [CrossRef]
- Ortiz-Moreno, M.L.; Sandoval-Parra, K.X.; Solarte-Murillo, L.V. Chlorella, a potential biofertilizer? Orinoquia 2019, 23, 71–78. [Google Scholar] [CrossRef]
- Kuhn, D.M.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Comparison of Biofilms Formed by Candidaalbicans and Candidaparapsilosis on Bioprosthetic Surfaces. Society 2002, 70, 878–888. [Google Scholar]
- Chen, M.Y.; Lee, D.J.; Tay, J.H. Distribution of Extracellular Polymeric Substances in Aerobic Granules. Appl. Microbiol. Biotechnol. 2007, 73, 1463–1469. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hupp, B.; Huszár, G.; Farkas, A.; Maróti, G. Algal Hydrogen Production and Exopolysaccharide Patterns in Chlorella–Bacillus Inter-Kingdom Co-Cultures. Fermentation 2023, 9, 424. https://doi.org/10.3390/fermentation9050424
Hupp B, Huszár G, Farkas A, Maróti G. Algal Hydrogen Production and Exopolysaccharide Patterns in Chlorella–Bacillus Inter-Kingdom Co-Cultures. Fermentation. 2023; 9(5):424. https://doi.org/10.3390/fermentation9050424
Chicago/Turabian StyleHupp, Bettina, Gabriella Huszár, Attila Farkas, and Gergely Maróti. 2023. "Algal Hydrogen Production and Exopolysaccharide Patterns in Chlorella–Bacillus Inter-Kingdom Co-Cultures" Fermentation 9, no. 5: 424. https://doi.org/10.3390/fermentation9050424
APA StyleHupp, B., Huszár, G., Farkas, A., & Maróti, G. (2023). Algal Hydrogen Production and Exopolysaccharide Patterns in Chlorella–Bacillus Inter-Kingdom Co-Cultures. Fermentation, 9(5), 424. https://doi.org/10.3390/fermentation9050424