
Effects of the Autochthonous Probiotic Bacteria Lactobacillus plantarum B and Lactococcus lactis Subsp. lactis S1 on the Proteolysis of Croatian Cheese Ripened in a Lambskin Sack (Sir iz Mišine)


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cheese Production
2.2. Sampling of Cheese
2.3. Analysis of Proteolytic Changes of the Cheese during Ripening
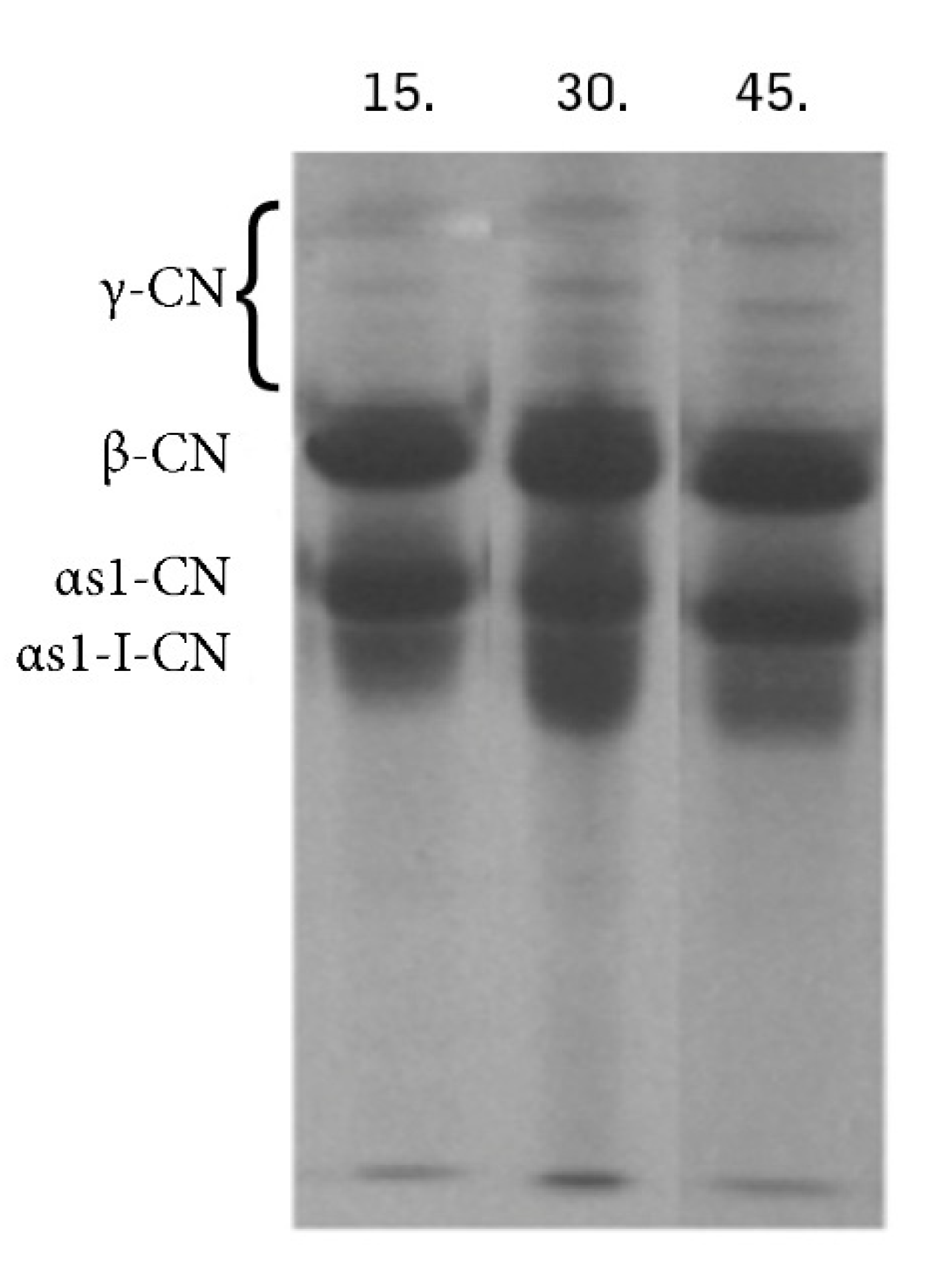
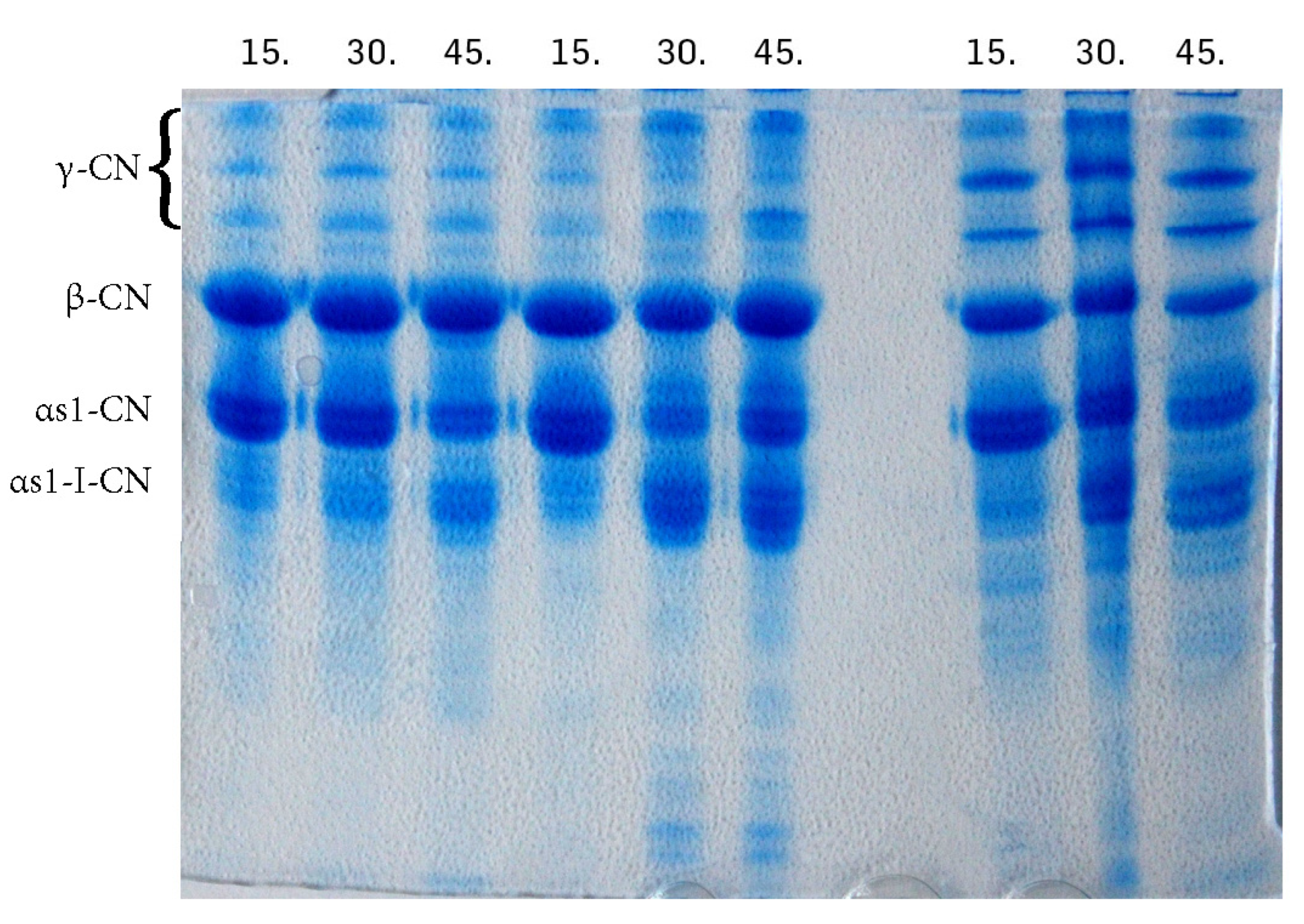
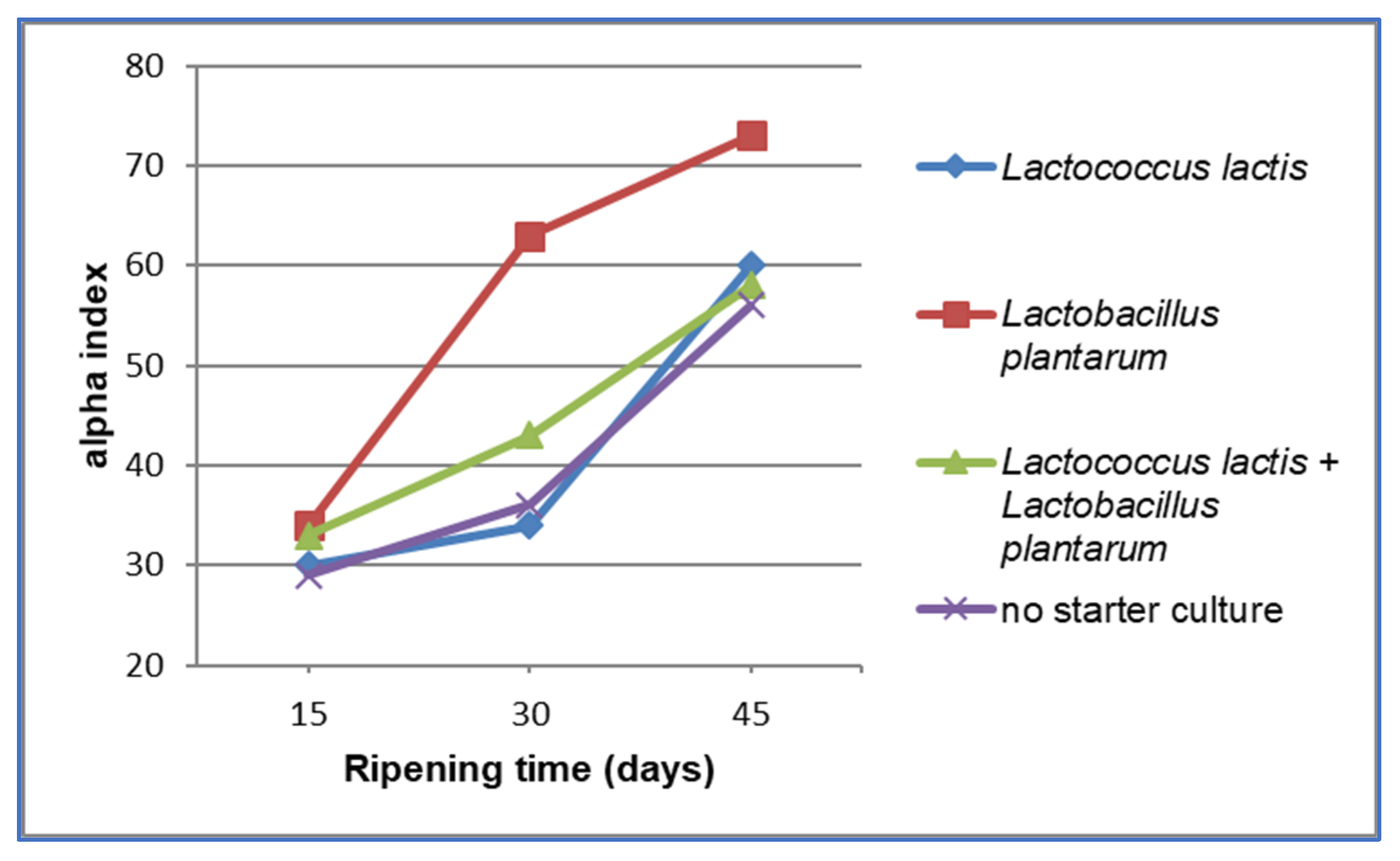
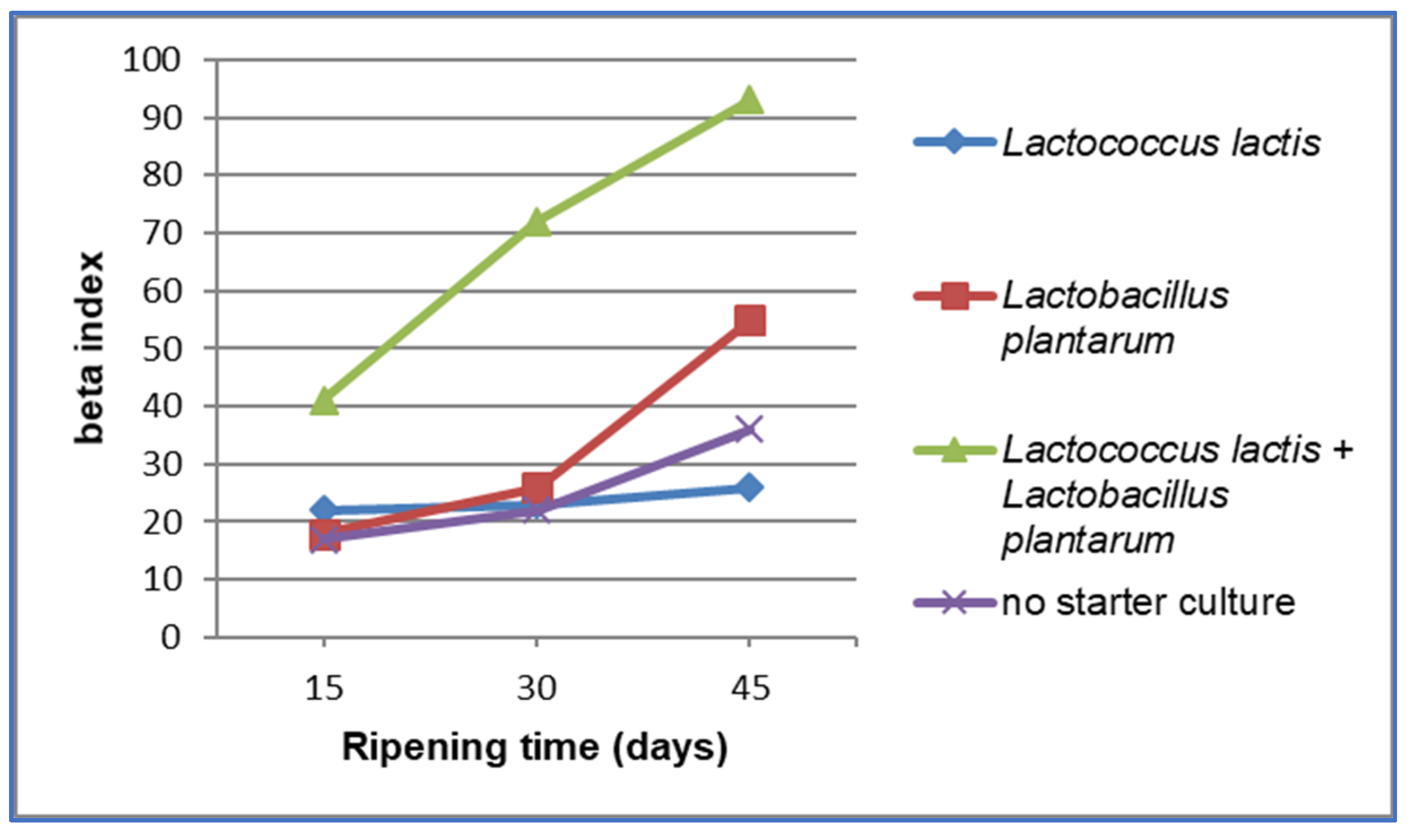
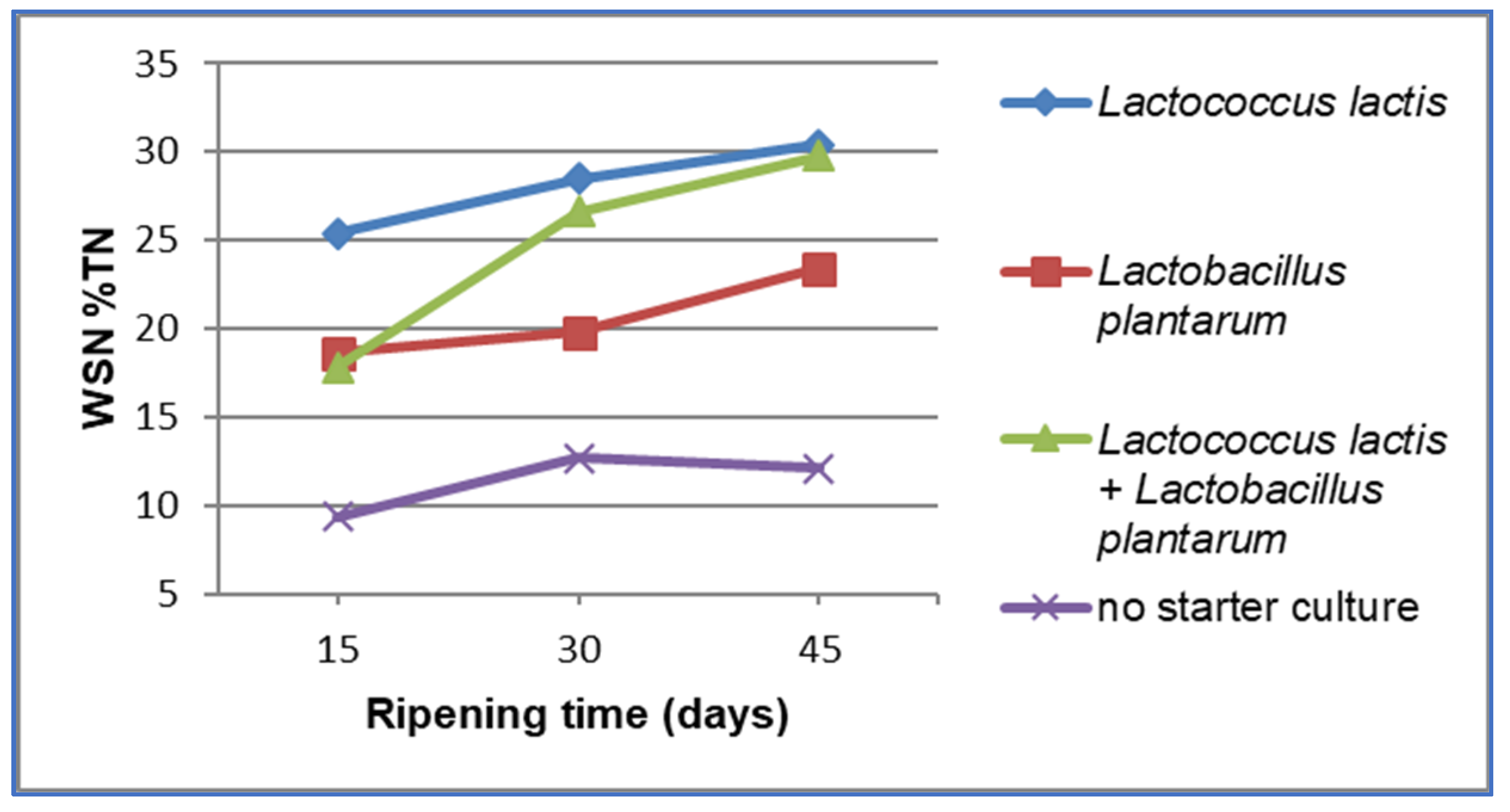
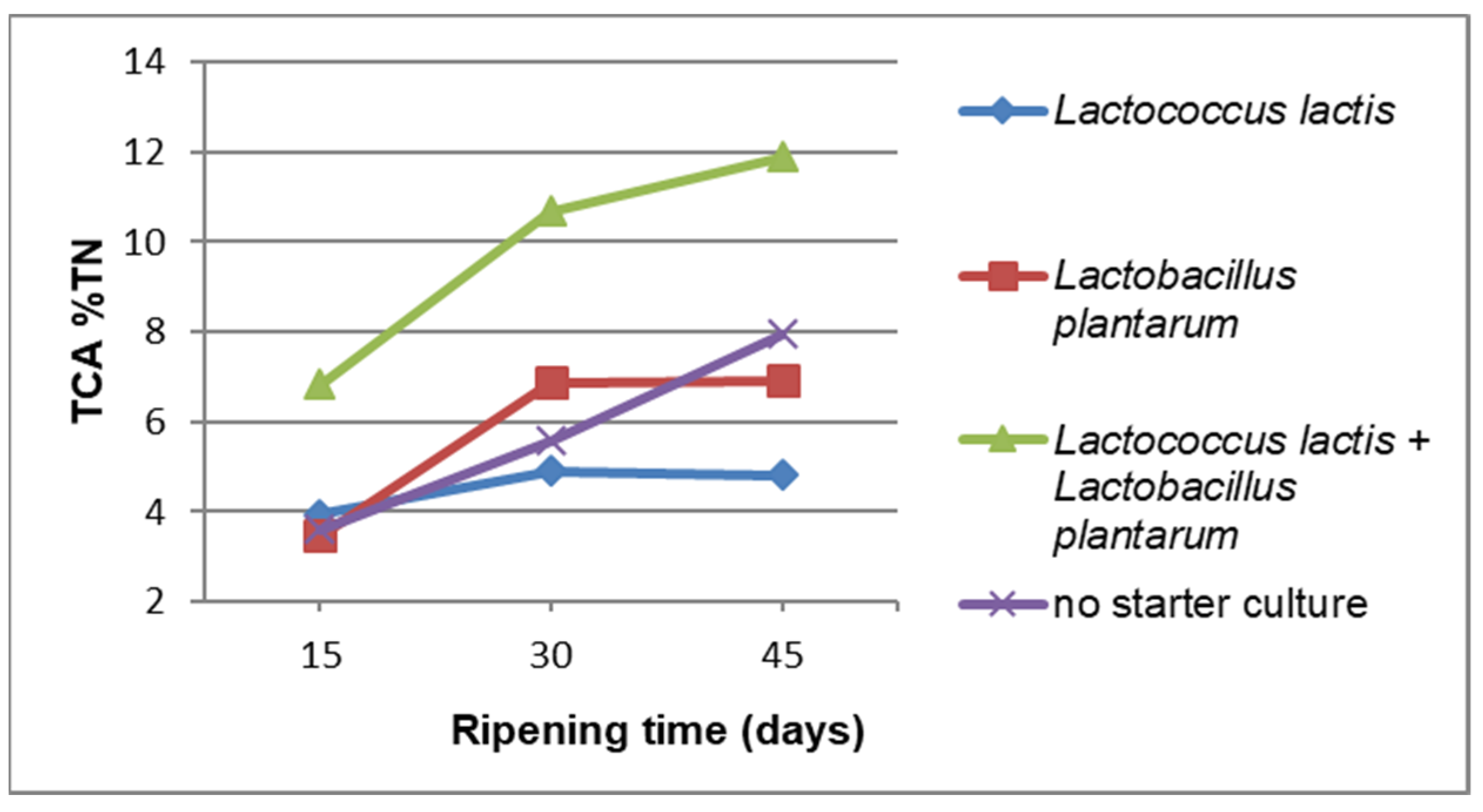
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McSweeney, P.L.H.; Sousa, M.J. Biochemical pathways for the production of flavour compounds in cheeses during ripening: A review. Le Lait 2000, 80, 293–324. [Google Scholar] [CrossRef]
- Havranek, J.; Kalit, S.; Antunac, N.; Samaržija, D. Sirarstvo, 1st ed.; Hrvatska Mljekarska Udruga: Zagreb, Croatia, 2014. [Google Scholar]
- Kalit, S.; Havranek, J.; Kapš, M.; Perko, B.; Čubrić-Čurik, V. Proteolysis and optimal ripening time of Tounj cheese. Int. Dairy J. 2005, 15, 619–624. [Google Scholar] [CrossRef]
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P.L.H. Cheese flavour. In Fundamentals of Cheese Science; Colilla, J., Ed.; Aspen Publishers: Gaithersburg, MD, USA, 2000; pp. 282–303. [Google Scholar]
- Cakmakci, S.; Gurses, M.; Gundogdu, E. The effect of different packaging materials on proteolysis, sensory scores and gross composition of tulum cheese. Afr. J. Biotechnol. 2011, 10, 4444–4449. [Google Scholar]
- Ardö, Y.; McSweeney, P.L.H.; Magboul, A.A.A.; Upadhyay, V.K.; Fox, P.F. Biochemistry of cheese ripening: Proteolysis. In Cheese Chemistry, Physics & Microbiology; McSweeney, P.L.H., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Elsevier Academic Press: London, UK, 2017; Volume 1, pp. 445–482. [Google Scholar] [CrossRef]
- Rako, A.; Tudor Kalit, M.; Rako, Z.; Petrović, D.; Kalit, S. Effect of composition and proteolysis on textural characteristics of Croatian cheese ripen in a lamb skin sack (Sir iz mišine). Mljekarstvo 2019, 69, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Hayaloglu, A.A.; Cakmakci, S.; Brechany, E.Y.; Deegan, K.C.; McSweeneys, P.L.H. Microbiology, biochemistry and volatile composition of Tulum cheese ripened in goat’s skin or plastic bags. J. Dairy Sci. 2007, 90, 1102–1121. [Google Scholar] [CrossRef]
- Tudor Kalit, M.; Kalit, S.; Delaš, I.; Kelava, N.; Karolyi, D.; Kaić, D.; Vrdoljak, M.; Havranek, J. Changes in the composition and sensory properties of Croatian cheese in a lamb skin sack (Sir iz misine) during ripening. Int. J. Dairy Technol. 2014, 67, 255–264. [Google Scholar] [CrossRef]
- Bijeljac, S.; Saricć, Z. Autohtoni Mliječni Proizvodi sa Osnovama Sirarstva, 1st ed.; Poljoprivredni Fakultet Univerziteta u Sarajevu: Sarajevo, Bosnia and Herzegovina, 2005. [Google Scholar]
- Yilmaz, G.; Ahmet, A.; Akin, N. The effect of microbial lipase on the lipolysis during the ripening of Tulum cheese. J. Food Eng. 2005, 69, 269–274. [Google Scholar] [CrossRef]
- Minervini, F.; Siragusa, S.; Faccia, M.; Dal Bello, F.; Gobbetti, M.; De Angelis, M. Manufacture of Fior di Latte cheese by incorporation of probiotic lactobacilli. Int. J. Dairy Sci. 2012, 95, 508–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezerra, T.K.A.; de Araujo, A.R.R.; do Nascimento, E.S.; de Matos Paz, J.E.; Gadelha, C.A.; Gadelha, T.S.; Pacheco, M.T.B.; Queiroga, R.C.R.E.; de Oliveira, M.E.G.; Madruga, M.S. Proteolysis in goat “coalho” cheese supplemented with probiotic lactic acid bacteria. Food Chem. 2016, 196, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Anihouvi, E.S.; Kesenkaş, H. Wagashi cheese: Probiotic bacteria incorporation and significance on microbiological, physicochemical, functional and sensory properties during storage. LWT 2022, 155, 112933. [Google Scholar] [CrossRef]
- Kostelac, D.; Vrdoljak, M.; Markov, K.; Delaš, I.; Jug, T.; Gajdoš Kljusurić, J.; Jakopović, Ž.; Čanak, I.; Jelić, M.; Frece, J. SPME-GC-MS and Multivariate Analysis of Sensory Properties of Cheese in a Sack Matured with Probiotic Starter Cultures. Food Technol. Biotechnol. 2020, 58, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Vinderola, G.; Prosello, W.; Molinari, F.; Ghiberto, D.; Reinheimer, J. Growth of Lactobacillus paracasei A13 in Argentinian probiotic cheese and its impact on the characteristics of the product. Int. J. Food Microbiol. 2009, 135, 171–174. [Google Scholar] [CrossRef]
- Albenzio, M.; Santillo, A.; Caroprese, M.; Ruggieri, D.; Napolitano, F.; Sevi, A. Physicochemical properties of Scamorza Ewe milk cheese manufactured with different probiotic cultures. J. Dairy Sci. 2013, 96, 2781–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cárdenas, N.; Calzada, J.; Peirotén, A.; Jiménez, E.; Escudero, R.; Rodríguez, J.M.; Medina, M.; Fernández, L. Development of a potential probiotic fresh cheese using two Lactobacillus salivarius strains isolated from human milk. Biomed Res. Int. 2014, 2014, 801918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dantas, A.B.; Jesus, V.F.; Silva, R.; Almada, C.N.; Esmerino, E.A.; Cappato, L.P.; Silva, M.C.; Raices, R.S.L.; Cavalcanti, R.N.; Carvalho, C.C.; et al. Manufacture of probiotic Minas frescal cheese with Lactobacillus casei Zhang. J. Dairy Sci. 2016, 99, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Mushtaq, M.; Gani, A.; Masoodi, F.A.; Ahmad, M. Himalayan cheese (Kalari/Kradi)—Effect of different probiotic strains on oxidative stability, microbiological, sensory and nutraceutical properties during storage. LWT 2016, 67, 74–81. [Google Scholar] [CrossRef]
- Yerlikaya, O.; Akbulut, N. Potential use of probiotic Enterococcus faecium and Enterococcus durans strains in Izmir Tulum cheese as adjunct culture. J. Food Sci. Technol. 2019, 56, 2175–2185. [Google Scholar] [CrossRef]
- Lopes, L.A.A.; Pimentel, T.C.; Carvalho, R.S.F.; Madruga, M.S.; Galvão, M.S.; Bezerra, T.K.A.; Barão, C.E.; Magnani, M.; Stamford, T.C.M. Spreadable goat Ricotta cheese added with Lactobacillus acidophilus La-05: Can microencapsulation improve the probiotic survival and the quality parameters? Food Chem. 2021, 346, 128769. [Google Scholar] [CrossRef]
- Silva, S.F.; Silva Rocha, R.; Almeida Esmerino, E.; Colombo Pimentel, T.; Gomes da Cruz, A.; Rodrigues Anjos, C.A. Impact of different modified atmosphere packaging on quality parameters and probiotic survival during storage of Minas Frescal cheese. Food Biosci. 2021, 43, 101338. [Google Scholar] [CrossRef]
- Suna, G.; Yilmaz-Ersan, L. Utilization of microalgae in probiotic white brined cheese. Mljekarstvo 2022, 72, 88–104. [Google Scholar] [CrossRef]
- Frece, J.; Vrdoljak, M.; Filipčić, M.; Jelić, M.; Čanak, I.; Jakopović, Ž.; Pleadin, J.; Gobin, I.; Landeka Dragičević, T.; Markov, K. Microbiological quality and variability of natural microbiota in Croatian cheese maturing in lambskin sacks. Food Technol. Biotechnol. 2016, 54, 129–134. [Google Scholar] [CrossRef]
- Vrdoljak, M. Enhancing of Functional Properties of the Cheese in a Sack by Using Probiotic Cultures Lactobacillus plantarum B and Lactococcus lactis subsp. lactis S1. Ph.D. Thesis, University of Zagreb, Faculty of Agriculture, Zagreb, Croatia, 16 December 2016. [Google Scholar]
- Mayer, H.K. Quality control of grated Parmesan products using an electrophoretic ripening index. Milchwissenschaft 1997, 52, 443–448. [Google Scholar]
- Hynes, E.R.; Meinardi, C.A.; Sabbag, N.; Cattaneo, T.; Candioti, M.C.; Zalazar, C.A. Influence of milk-clotting enzyme concentration on the αs1-casein hydrolysis during soft cheeses ripening. J. Dairy Sci. 2001, 84, 1335–1340. [Google Scholar] [CrossRef]
- ISO 8968-2; Milk—Determination of Nitrogen Content—Part 1: Milk and Milk Products—Determination of Nitrogen Content—Part 1: Kjeldahl Principle and Crude Protein Calculation. International Organization for Standards: Geneva, Switzerland, 2014.
- Magdić, V.; Kalit, S.; Mrkonjić Fuka, M.; Skelin, A.; Samaržija, D. A survey on hygienic and physicochemical properties of Istrian cheese. Mljekarstvo 2013, 63, 55–63. [Google Scholar]
- Sousa, M.J.; Ardo, Y.; McSweeney, P.L.H. Advances in the study of proteolysis during cheese ripening. Int. Dairy J. 2001, 11, 327–345. [Google Scholar] [CrossRef]
- Bergamini, C.V.; Hynes, E.R.; Palma, S.B.; Sabbag, N.G.; Zalazar, C.A. Proteolytic activity of three probiotic strains in semi-hard cheese as single and mixed cultures: Lactobacillus acidophilus, Lactobacillus paracasei and Bifidobacterium lactis. Int. Dairy J. 2009, 19, 467–475. [Google Scholar] [CrossRef]
- Børstig, M.W.; Karsten, B.Q.; Ardo, Y. Influence of pH on retention of camel chymosin in curd. Int. Dairy J. 2016, 38, 133–135. [Google Scholar] [CrossRef]
- Lane, C.N.; Fox, P.F. Contribution of starter and adjunct lactobacilli to proteolysis in Cheddar cheese during ripening. Int. Dairy J. 1996, 6, 715–728. [Google Scholar] [CrossRef]
- Kalit, S.; Lukač Havranek, J. Plasmin: Indogena proteinaza mlijeka. Mljekarstvo 2002, 52, 191–206. [Google Scholar]
- Lavasani, A.R.Z.; Ehsani, M.R.; Mirdamadi, S.; Mousavi, M.A.E.Z. Changes in physicochemical and organoleptic properties of traditional Iranian cheese Lighvan during ripening. Int. J. Dairy Technol. 2012, 65, 64–70. [Google Scholar] [CrossRef]
- Tudor Kalit, M.; Kalit, S.; Kelava, N.; Havranek, J. Physicochemical differences between Croatian cheese matured in a lamb skin sack (Sir iz misine) and cheese matured in a rind throughout ripening. Int. J. Dairy Technol. 2012, 65, 555–560. [Google Scholar] [CrossRef]
- Celik, O.F.; Tarakci, Z. The effects of starter cultures on chemical, biochemical and sensory properties of low-fat Tulum cheeses during ripening. Int. J. Dairy Technol. 2017, 70, 583–591. [Google Scholar] [CrossRef]
- Hayaloglu, A.A.; Tolu, C.; Yaser, K.; Sahingil, D. Volatiles and sensory evaluation of goat milk cheese Gokceada as affected by goat breeds (Gokceada and Turkish Saanen) and starter culture systems during ripening. J. Dairy Sci. 2013, 96, 2765–2780. [Google Scholar] [CrossRef] [PubMed]
- Hayaloglu, A.A.; Fox, P.F.; Guven, M.; Cakmakci, S. Cheeses of Turkey: 1. Varieties ripened in goat-skin bags. Le Lait 2007, 87, 79–95. [Google Scholar] [CrossRef]
- Bedel, A.; Kilic, S. Izmir Tulum peynirinin olgunlasma suresi uzerinde kulturun veticari enzimin rolu. EU Ziraat Fak. Derg. 2000, 29, 71–78. [Google Scholar]
- Bansal, N.; Piraino, P.; McSweeney, P.L.H. Determination of proteolysis in cheese. In Handbook of Dairy Food Analysis; Nollet, L.M.L., Toldrá, F., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2010; pp. 405–426. [Google Scholar]
- Medjoudj, H.; Zidoune, M.N.; Hayaloglu, A.A. Proteolysis and volatile profile in the Algerian traditional Bouhezza cheese made using raw goat’s milk. Int. J. Food Prop. 2017, 20, 1876–1893. [Google Scholar] [CrossRef] [Green Version]
- Desmazeaud, M.J.; Gripon, J.C. General mechanism of protein breakdown during cheese ripening. Milchwissenschaft 1977, 32, 731–734. [Google Scholar]
- Addeo, F.; Chianese, L.; Salzano, A.; Sacchi, R.; Cappuccio, U.; Ferranti, P.; Malorni, A. Characterization of the 12% trichloroacetic acid insoluble oligopeptides of Parmigiano-Reggiano cheese. J. Dairy Res. 1992, 59, 401–411. [Google Scholar] [CrossRef]
- Law, B.A. Cheese ripening and cheese flavour technology. In Technology of Cheesemaking; Law, B.A., Tamime, A.Y., Eds.; Sheffield Academic Press: Sheffield, UK, 1999; pp. 163–192. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrdoljak, M.; Tudor Kalit, M.; Dolenčić Špehar, I.; Radeljević, B.; Jelić, M.; Mandinić, S.; Frece, J.; Kalit, S. Effects of the Autochthonous Probiotic Bacteria Lactobacillus plantarum B and Lactococcus lactis Subsp. lactis S1 on the Proteolysis of Croatian Cheese Ripened in a Lambskin Sack (Sir iz Mišine). Fermentation 2022, 8, 382. https://doi.org/10.3390/fermentation8080382
Vrdoljak M, Tudor Kalit M, Dolenčić Špehar I, Radeljević B, Jelić M, Mandinić S, Frece J, Kalit S. Effects of the Autochthonous Probiotic Bacteria Lactobacillus plantarum B and Lactococcus lactis Subsp. lactis S1 on the Proteolysis of Croatian Cheese Ripened in a Lambskin Sack (Sir iz Mišine). Fermentation. 2022; 8(8):382. https://doi.org/10.3390/fermentation8080382
Chicago/Turabian StyleVrdoljak, Marija, Milna Tudor Kalit, Iva Dolenčić Špehar, Biljana Radeljević, Marko Jelić, Sandra Mandinić, Jadranka Frece, and Samir Kalit. 2022. "Effects of the Autochthonous Probiotic Bacteria Lactobacillus plantarum B and Lactococcus lactis Subsp. lactis S1 on the Proteolysis of Croatian Cheese Ripened in a Lambskin Sack (Sir iz Mišine)" Fermentation 8, no. 8: 382. https://doi.org/10.3390/fermentation8080382
APA StyleVrdoljak, M., Tudor Kalit, M., Dolenčić Špehar, I., Radeljević, B., Jelić, M., Mandinić, S., Frece, J., & Kalit, S. (2022). Effects of the Autochthonous Probiotic Bacteria Lactobacillus plantarum B and Lactococcus lactis Subsp. lactis S1 on the Proteolysis of Croatian Cheese Ripened in a Lambskin Sack (Sir iz Mišine). Fermentation, 8(8), 382. https://doi.org/10.3390/fermentation8080382