
Preservation of Human Gut Microbiota Inoculums for In Vitro Fermentations Studies

 ,
,  ,
, 
Abstract

1. Introduction
2. Materials and Methods
2.1. Reagents/Chemicals and Apparatus
2.2. Procedures for Comparison of the Use of Individual and Pooled Inoculum
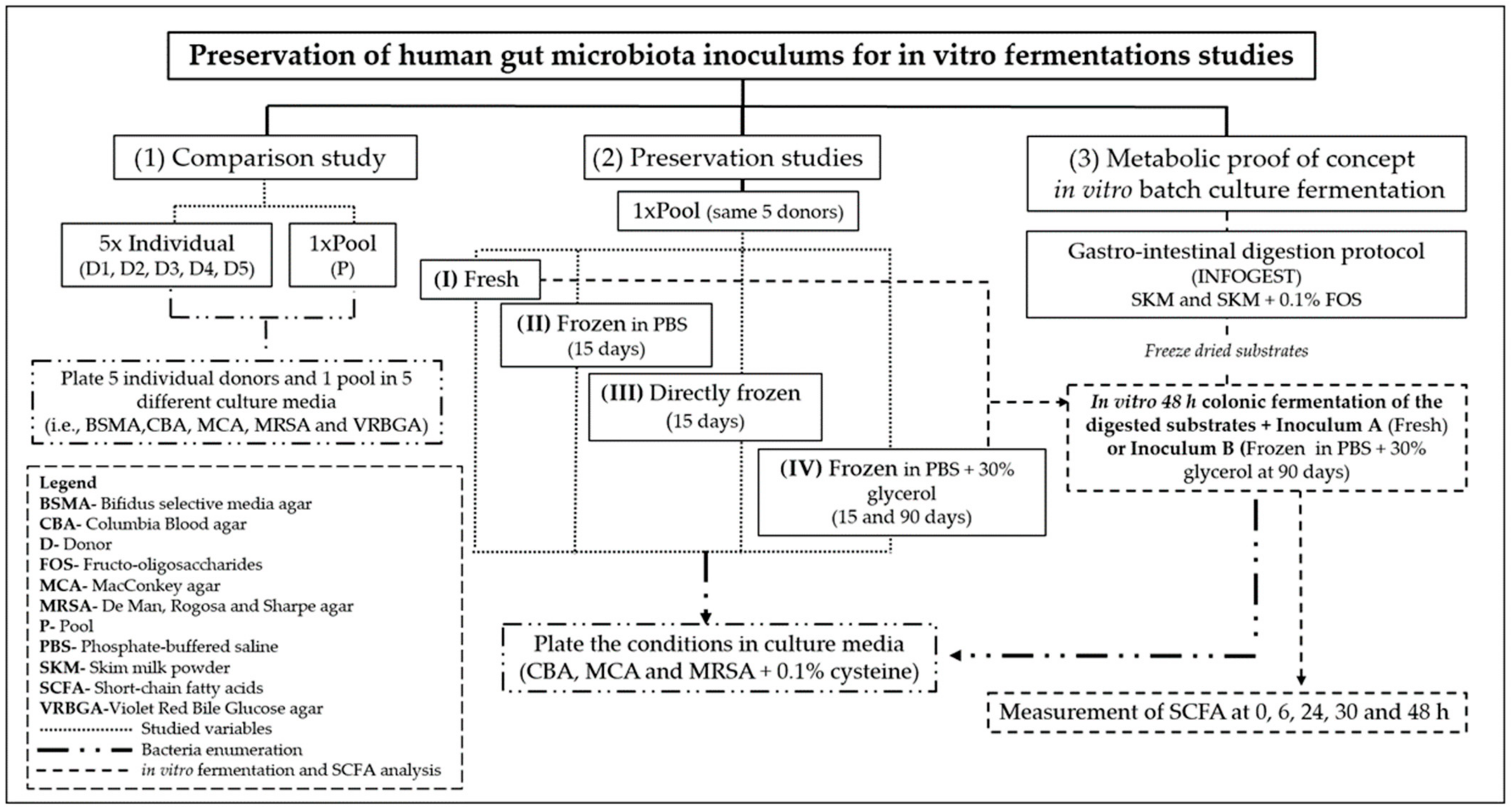
2.3. Sample Processing for Preservation Studies
2.4. Colonic Fermentations Using Fresh and Frozen Inoculum
2.5. Human Gastrointestinal Tract (GIT) Simulation Model Using Preserved Inoculum
2.5.1. In vitro Gastrointestinal Digestion Protocol (INFOGEST)
2.5.2. In vitro Batch-Culture Fermentations (Colonic Fermentation)
2.6. Microbiological Assays
2.7. Determination of Short Chain Fatty Acids (SCFA), Lactate, and Glycerol/Glycerol Derivatives Produced during In Vitro Fermentation
2.8. Statistical Analysis
3. Results and Discussion
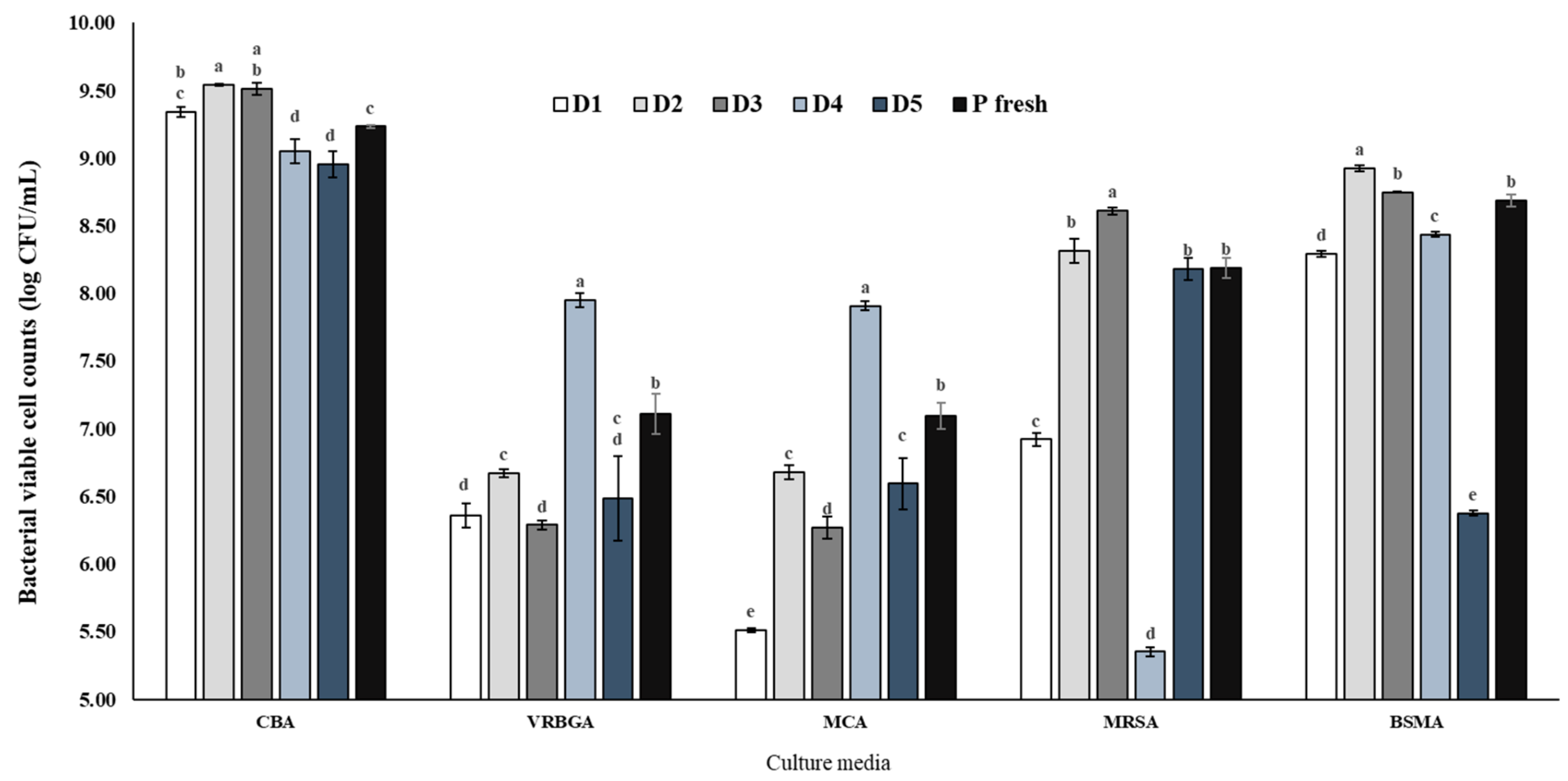
3.1. Differences between Using Individual and Pooled Inoculum
3.1.1. pH Variation
3.1.2. Bacterial Viability and Diversity on Individual and Pooled Inoculum
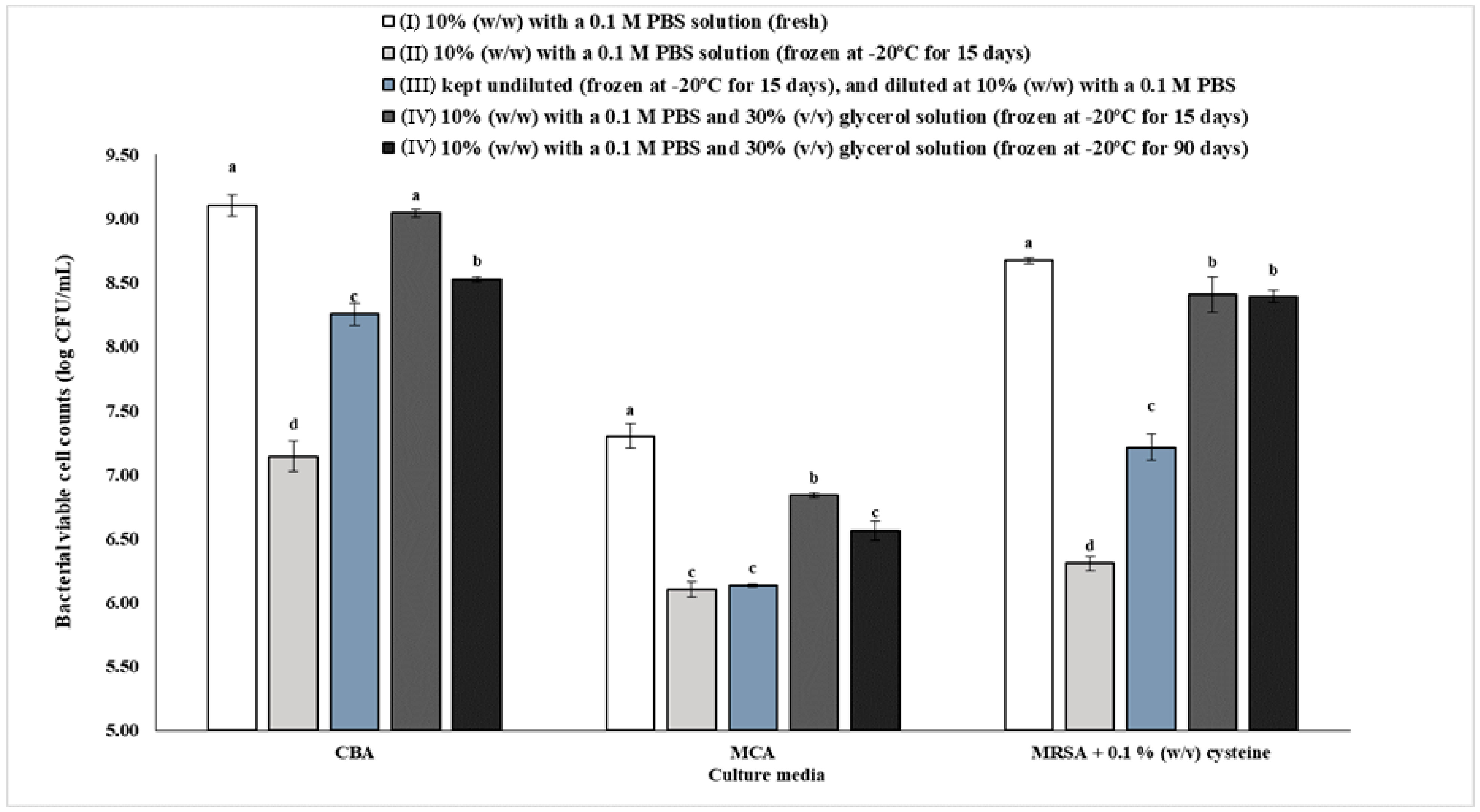
3.2. Effect of Preservation on Gut Bacteria Viability and Diversity
Gut Bacteria Viability of the Pooled Inoculums Preserved under Different Conditions (I–IV)
3.3. Impact of Using Fresh and Frozen Inoculum in Colonic Batch Fermentations
3.3.1. Glycerol Wash-out Confirmation
3.3.2. pH Variation
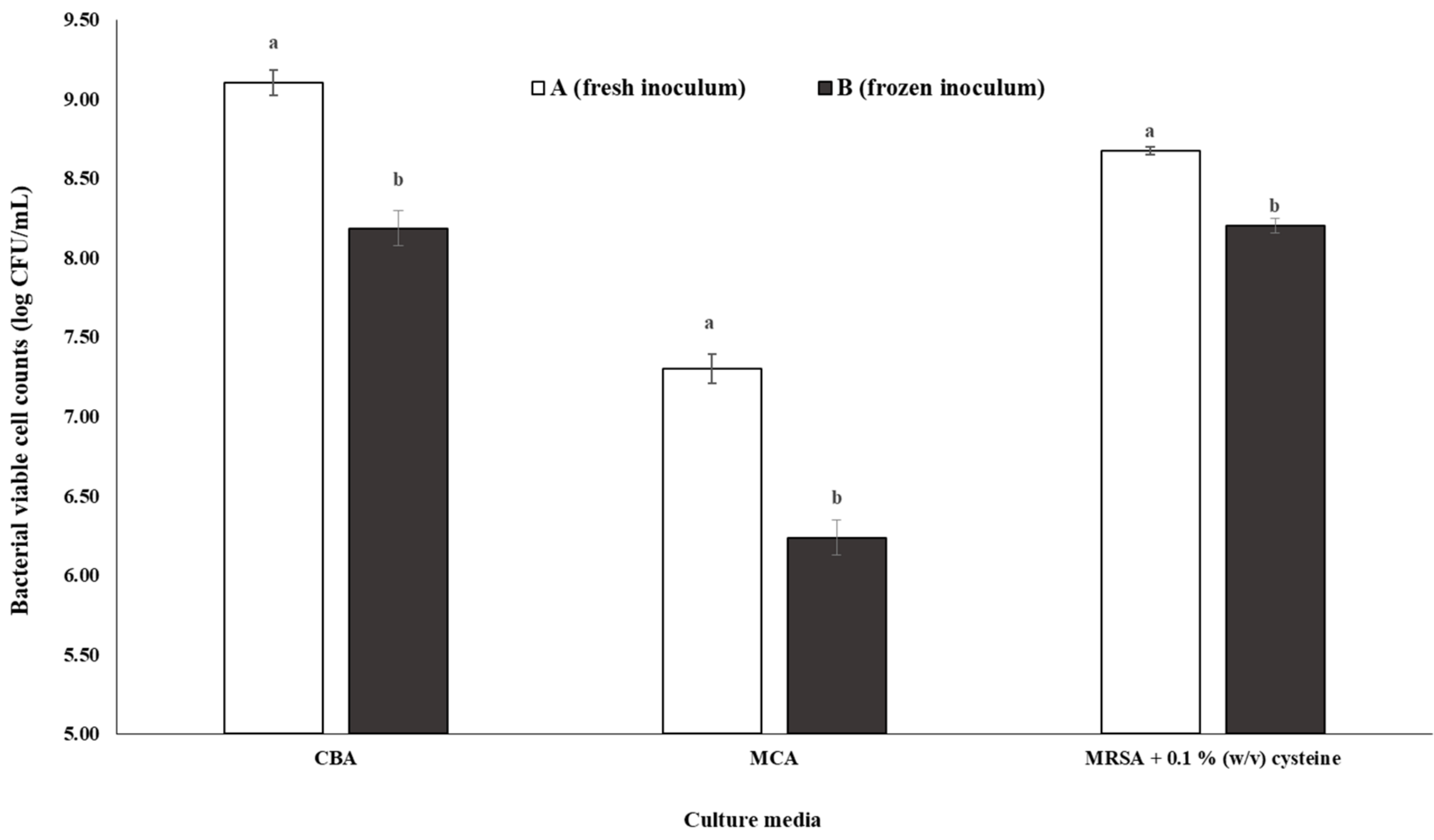
3.3.3. Gut Bacteria Enumeration of Inoculum A and B
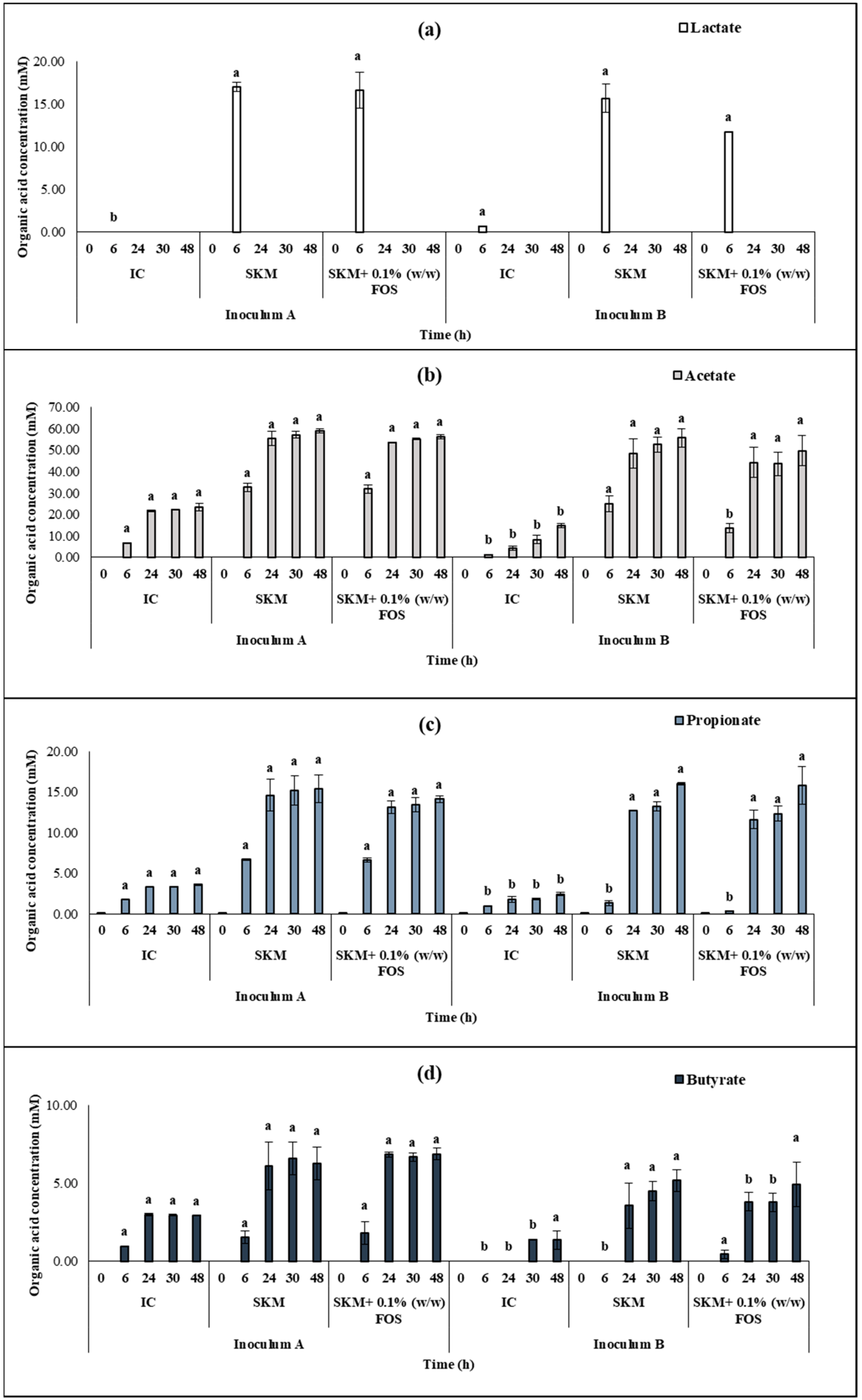
3.4. Human in vitro Colonic Fermentation Using Fresh and Frozen Inoculums
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Materials and Methods
Appendix A.1. Reagents/Chemicals Used in This Study
- 1-propanol (Sigma, St. Louis, MO, USA);
- 1,3-propanediol (Sigma, St. Louis, MO, USA);
- 2-propanol (Honeywell Riedel-de Haen, Munich, Germany);
- Acetic acid glacial (Sigma, St. Louis, MO, USA);
- Ammonium Carbonate-(NH4)2CO3 (Merck KGaA, Darmstadt, Germany);
- Bile bovine (Sigma, St. Louis, MO, USA);
- Bile salts (Sigma, St. Louis, MO, USA);
- Bile acid assay kit (Sigma, St. Louis, MO, USA);
- Butyric acid (Sigma, St. Louis, MO, USA);
- Calcium chloride dihydrate—CaCl2(H2O)2 (Carlo Erba Reagents, Barcelona, Spain)
- Calcium chloride hexahydrate—CaCl2(H2O)6 (Sigma, St. Louis, MO, USA);
- Defibrinated sheep blood OxoidTM (Thermo Fischer Scientific, Waltham, MA, USA)
- Dipotassium hydrogen phosphate-K2HPO4 (Honeywell Fluka, Seelze, Germany);
- DL-lactic acid (Sigma, St. Louis, MO, USA);
- Fructo-oligosaccharides from chicory root (FOS) (Megazyme, Bray, Ireland);
- Glycerol—analytical grade (Fisher Scientific, Loughborough, UK);
- Hemin (Sigma, St. Louis, MO, USA);
- Hemoglobin (Sigma, St. Louis, MO, USA);
- Hydrochloric acid- HCl (Honeywell Fluka, Seelze, Germany);
- L-cysteine HCl (Sigma-Aldrich, St. Louis, MO, USA);
- Magnesium chloride hexahydrate-MgCl2(H2O)6 (Panreac, Barcelona, Spain);
- Magnesium sulfate heptahydrate-MgSO4(H2O)7 (Sigma, St. Louis, MO, USA);
- Molico skim milk powder-SKM (Nestlé S.A., Vevey, Switzerland);
- Na-p-tosyl-L-arginine methyl esther hydrochloride—TAME (Sigma, St. Louis, MO, USA);
- Pancreatin from porcine pancreas (Sigma, St. Louis, MO, USA);
- Pepsin from porcine gastric mucosa powder (Sigma, St. Louis, MO, USA);
- Peptone from animal tissue (Sigma, St. Louis, MO, USA);
- Phosphate buffered saline (Dulbecco A) OxoidTM (Thermo Fischer Scientific, Waltham, MA, USA);
- Potassium chloride-KCl (Honeywell Fluka, Seelze, Germany);
- Potassium dihydrogen phosphate-KH2PO4 (Merck KGaA, Darmstadt, Germany);
- Propionic acid (Sigma, St. Louis, MO, USA);
- Resazurin sodium salt (Sigma, St. Louis, MO, USA);
- Sodium chloride-NaCl (Honeywell Fluka, Seelze, Germany);
- Sodium hydrogen carbonate-NaHCO3 (Panreac, Barcelona, Spain);
- Sodium hydroxide—NaOH (LabChem, Zelienople, USA);
- Sulfuric acid—H2SO4 (Honeywell Fluka, Seelze, Germany);
- Trichloroacetic acid—TCA (Sigma, St. Louis, MO, USA);
- Tris(hydroxymethyl)aminomethane hydrochloride (Merck KGaA, Darmstadt, Germany)
- Tween 80 (Sigma, St. Louis, MO, USA);
- Vitamin K1 (Sigma, St. Louis, MO, USA);
- Yeast extract (Sigma, St. Louis, MO, USA).
Appendix A.2. Culture Media
- Bifidus selective medium agar-BSMA (Sigma, St. Louis, MO, USA);
- Columbia agar base-CBA (Liofilchem, Roseto degli Abruzzi, Italy);
- de Man, Rogosa and Sharpe agar-MRSA (Biokar Diagnostics, Allonne, France);
- MacConkey agar-MCA (Biolife, Milan, Italy);
- Violet red bile glucose agar—VRBGA (Biokar diagnostics, Allonne, France).
Appendix A.3. Apparatus Used in This Study
- 1 kDa molecular weight cut-off regenerated cellulose dialysis tubing Spectra/Por® 6 (Spectrum, New Brunswick, NJ, USA);
- Agilent 1260 II series HPLC (Agilent, Santa Clara, CA, USA);
- Alpha 2–4 LSC plus model (Martin Christ Gefriertrocknungsanlagen GmbH, Osterode am Harz, Germany)
- Anaerobic cabinet, Whitley A35 workstation (Don Whitley Scientific, Bingley, UK);
- FerMac 260 pH controller (Electrolab Biotech Ltd., Gloucestershire, UK);
- Heraeus™ Megafuge™ 16R Centrifuge (Thermo Fischer Scientific, Waltham, MA, USA)
- Ion-exclusion Aminex HPX-87H column (Biorad, Hercules, CA, USA);
- Mixwel® laboratory blender (Alliance Bio Expertise, Guipry, France);
- MR Hei-Tec magnetic stirrer (Heidolph Instruments GmbH & CO. KG, Schwabach, Germany)
- MST magnetic stirrer (Velp Scientifica, Usmate Velate, Italy)
- OxoidTM AnaeroGenTM 2.5 L sachet (Thermo Fischer Scientific, Waltham, MA, USA);
- OxoidTM AnaeroJarTM 2.5 L (Thermo Fischer Scientific, Waltham, MA, USA);
- Reax top vortex (Heidolph Instruments GmbH & CO. KG, Schwabach, Germany);
- Refrigerator Beko RSNE445E33WN (Beko, Istanbul, Turkey);
- SevenCompact pH meter (Mettler Toledo, Urdorf, Switzerland);
- Synergy H1 Hybrid Multi-Mode Reader (BioTek, Winooski, VT, USA);
- UV-1900 UV-VIS spectrophotometer (Shimadzu, Kyoto, Japan);
- Tamper proof specimen 1-L containers (Sigma, St. Louis, MO, USA).
Appendix B. Human Fecal Sample Collection Protocol
References
- Maki, J.J.; Klima, C.L.; Sylte, M.J.; Looft, T. The Microbial Pecking Order: Utilization of Intestinal Microbiota for Poultry Health. Microorganisms 2019, 7, 376. [Google Scholar] [CrossRef] [PubMed]
- Grond, K.; Sandercock, B.K.; Jumpponen, A.; Zeglin, L.H. The avian gut microbiota: Community, physiology and function in wild birds. J. Avian Biol. 2018, 49, e01788. [Google Scholar] [CrossRef]
- Yan, W.; Sun, C.; Zheng, J.; Wen, C.; Ji, C.; Zhang, D.; Chen, Y.; Hou, Z.; Yang, N. Efficacy of Fecal Sampling as a Gut Proxy in the Study of Chicken Gut Microbiota. Front. Microbiol. 2019, 10, 2126. [Google Scholar] [CrossRef] [PubMed]
- Pastorelli, G.; Faustini, M.; Attard, E. In vitro fermentation of feed ingredients by fresh or frozen pig fecal inocula. Anim. Sci. J. 2014, 85, 690–697. [Google Scholar] [CrossRef]
- Doke, S.K.; Dhawale, S.C. Alternatives to animal testing: A review. Saudi Pharm. J. 2015, 23, 223–229. [Google Scholar] [CrossRef]
- Amorim, C.; Silvério, S.C.; Cardoso, B.B.; Alves, J.I.; Pereira, M.A.; Rodrigues, L.R. In vitro fermentation of raffinose to un-ravel its potential as prebiotic ingredient. LWT 2020, 126, 109322. [Google Scholar] [CrossRef]
- Aguirre, M.; Ramiro-Garcia, J.; Koenen, M.; Venema, K. To pool or not to pool? Impact of the use of individual and pooled fecal samples for in vitro fermentation studies. J. Microbiol. Methods 2014, 107, 1–7. [Google Scholar] [CrossRef]
- Ashaolu, T.J.; Ashaolu, J.O.; Adeyeye, S.A.O. Fermentation of prebiotics by human colonic microbiota in vitro and short-chain fatty acids production: A critical review. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef]
- Murray, J.-A.M.D.; McMullin, P.; Handel, I.; Hastie, P.M. The effect of freezing on the fermentative activity of equine faecal inocula for use in an in vitro gas production technique. Anim. Feed Sci. Technol. 2012, 178, 175–182. [Google Scholar] [CrossRef]
- Bosch, G.; Wrigglesworth, D.J.; Cone, J.W.; Pellikaan, W.F.; Hendriks, W.H. Effects of preservation conditions of canine feces on in vitro gas production kinetics and fermentation end products. J. Anim. Sci. 2013, 91, 259–267. [Google Scholar] [CrossRef]
- Prates, A.; De Oliveira, J.; Abecia, L.; Fondevila, M. Effects of preservation procedures of rumen inoculum on in vitro microbial diversity and fermentation. Anim. Feed. Sci. Technol. 2010, 155, 186–193. [Google Scholar] [CrossRef]
- Aguirre, M.; Eck, A.; Koenen, M.; Savelkoul, P.H.M.; Budding, A.E.; Venema, K. Evaluation of an optimal preparation of human standardized fecal inocula for in vitro fermentation studies. J. Microbiol. Methods 2015, 117, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Hervás, G.; Frutos, P.; Giráldez, F.J.; Mora, M.J.; Fernández, B.; Mantecón, Á.R. Effect of preservation on fermentative activity of rumen fluid inoculum for in vitro gas production techniques. Anim. Feed Sci. Technol. 2005, 123, 107–118. [Google Scholar] [CrossRef]
- Waite, D.W.; Deines, P.; Taylor, M.W. Quantifying the impact of storage procedures for faecal bacteriotherapy in the critically endangered New Zealand Parrot, the Kakapo (Strigops habroptilus). Zoo Biol. 2013, 32, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; Colombatto, D.; Morgavi, D.P.; Yang, W.Z.; Rode, L.M. Mode of action of exogenous cell wall degrading enzymes for ruminants. Can. J. Anim. Sci. 2004, 84, 13–22. [Google Scholar] [CrossRef]
- Henderson, A.R.; McDonald, P.; Anderson, D. The effect of a cellulase preparation derived fromTrichoderma viride on the chemical changes during the ensilage of grass, lucerne and clover. J. Sci. Food Agric. 1982, 33, 16–20. [Google Scholar] [CrossRef]
- Murray, J.M.D.; Longland, A.; Moore-Colyer, M.; Dunnett, C. The effect of feeding a low- or a high-starch diet on the in vitro fermentative capacity of equine faecal inocula. Anim. Sci. 2006, 82, 627–635. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Alegría-Torán, A.; Garcia-Llatas, G.; Cilla, A. Static Digestion Models: General Introduction; Springer Science and Business Media LLC: Cham, Germany, 2015; pp. 3–12. [Google Scholar]
- Verhoeckx, K.; Cotter, P.; López-Expósito, I.; Kleiveland, C.; Lea, T.; Mackie, A.; Requena, T.; Swiatecka, D.; Wichers, H. The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer Nature: London, UK; Berlin, Germany; New York, NY, USA, 2015. [Google Scholar]
- De Carvalho, N.M.; Walton, G.; Poveda, C.; Silva, S.; Amorim, M.; Madureira, A.; Pintado, M.; Gibson, G.; Jauregi, P. Study of in vitro digestion of Tenebrio molitor flour for evaluation of its impact on the human gut microbiota. J. Funct. Foods 2019, 59, 101–109. [Google Scholar] [CrossRef]
- Sánchez-Patán, F.; Cueva, C.; Monagas, M.; Walton, G.E.; Gibson, G.R.; Quintanilla-López, J.E.; Lebrón-Aguilar, R.; Martin-Alvarez, P.J.; Moreno-Arribas, M.V.; Bartolomé, B. In vitro fermentation of a red wine extract by human gut microbi-ota: Changes in microbial groups and formation of phenolic metabolites. J. Agric. Food Chem. 2012, 60, 2136–2147. [Google Scholar] [CrossRef]
- Miles, A.A.; Misra, S.S.; Irwin, J.O. The estimation of the bactericidal power of the blood. Epidemiol. Infect. 1938, 38, 732–749. [Google Scholar] [CrossRef] [PubMed]
- Ohigashi, S.; Sudo, K.; Kobayashi, D.; Takahashi, O.; Takahashi, T.; Asahara, T.; Nomoto, K.; Onodera, H. Changes of the Intestinal Microbiota, Short Chain Fatty Acids, and Fecal pH in Patients with Colorectal Cancer. Dig. Dis. Sci. 2013, 58, 1717–1726. [Google Scholar] [CrossRef] [PubMed]
- Osuka, A.; Shimizu, K.; Ogura, H.; Tasaki, O.; Hamasaki, T.; Asahara, T.; Nomoto, K.; Morotomi, M.; Kuwagata, Y.; Shimazu, T. Prognostic impact of fecal pH in critically ill patients. Crit. Care 2012, 16, R119. [Google Scholar] [CrossRef] [PubMed]
- Watson, E.-J.; Giles, J.; Scherer, B.L.; Blatchford, P. Human faecal collection methods demonstrate a bias in microbiome composition by cell wall structure. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Prakash, O.; Nimonkar, Y.; Desai, D. A Recent Overview of Microbes and Microbiome Preservation. Indian J. Microbiol. 2020, 60, 297–309. [Google Scholar] [CrossRef]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Ilhan, Z.E.; Marcus, A.K.; Kang, D.-W.; Rittmann, B.E.; Krajmalnik-Brown, R. pH-mediated microbial and metabolic inter-actions in fecal enrichment cultures. Msphere 2017, 2, e00047-17. [Google Scholar] [CrossRef]
- Henrick, B.M.; Hutton, A.A.; Palumbo, M.C.; Casaburi, G.; Mitchell, R.D.; Underwood, M.A.; Smilowitz, J.T.; Frese, S.A. El-evated fecal pH indicates a profound change in the breastfed infant gut microbiome due to reduction of Bifidobacterium over the past century. MSphere 2018, 3, e00041-18. [Google Scholar] [CrossRef]
- Mathur, H.; Beresford, T.P.; Cotter, P.D. Health Benefits of Lactic Acid Bacteria (LAB) Fermentates. Nutrients 2020, 12, 1679. [Google Scholar] [CrossRef]
- Saez-Lara, M.J.; Gomez-Llorente, C.; Plaza-Diaz, J.; Gil, A. The role of probiotic lactic acid bacteria and bifidobacteria in the prevention and treatment of inflammatory bowel disease and other related diseases: A systematic review of randomized human clinical trials. Biomed. Res. Int. 2015, 2015, 505878. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, M. Fermentation of Indigestible Carbohydrates by the Gut Microbiota: One Small Step for a Microbe, a Giant Leap for Mankind? Ph.D. Thesis, Maastricht University, Maastricht, The Netherlands, 2016. [Google Scholar]
- Vandeputte, D.; Tito, R.Y.; Vanleeuwen, R.; Falony, G.; Raes, J. Practical considerations for large-scale gut microbiome studies. FEMS Microbiol. Rev. 2017, 41, S154–S167. [Google Scholar] [CrossRef] [PubMed]
- Costello, S.P.; Conlon, M.A.; Vuaran, M.S.; Roberts-Thomson, I.C.; Andrews, J.M. Faecal microbiota transplant for recur-rent Clostridium difficile infection using long-term frozen stool is effective: Clinical efficacy and bacterial viability data. Aliment. Pharmacol. Ther. 2015, 42, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Achá, S.; Kühn, I.; Mbazima, G.; Colque-Navarro, P.; Möllby, R. Changes of viability and composition of the Escherichia coli flora in faecal samples during long time storage. J. Microbiol. Methods 2005, 63, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Hubálek, Z. Protectants used in the cryopreservation of microorganisms. Cryobiology 2003, 46, 205–229. [Google Scholar] [CrossRef]
- Gaci, N.; Chaudhary, P.P.; Tottey, W.; Alric, M.; Brugère, J.-F. Functional amplification and preservation of human gut microbiota. Microb. Ecol. Health Dis. 2017, 28, 1308070. [Google Scholar] [CrossRef]
- Bircher, L.; Schwab, C.; Geirnaert, A.; Lacroix, C. Cryopreservation of artificial gut microbiota produced with in vitro fermentation technology. Microb. Biotechnol. 2018, 11, 163–175. [Google Scholar] [CrossRef]
- Parkar, S.G.; Davis, P.; Trower, T.M.; Hedderley, D.I.; Sutton, K.H.; Ingram, J.R. Factors affecting microbial metabolism in a human fecal fermentation model to evaluate prebiotics. J. Nutr. Intermed. Metab. 2019, 15, 55–57. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; Reyes-Gavilán, C.G.D.L.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nu-trients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, N.M.; Costa, E.M.; Silva, S.; Pimentel, L.; Fernandes, T.H.; Pintado, M.E. Fermented Foods and Beverages in Human Diet and Their Influence on Gut Microbiota and Health. Fermentation 2018, 4, 90. [Google Scholar] [CrossRef]
- Beards, E.; Tuohy, K.; Gibson, G. Bacterial SCFA and gas profiles of a range of food ingredients following in vitro fer-mentation by human colonic microbiota. Anaerobe 2010, 16, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gibson, G.R.; Walton, G.E. An in vitro approach to study effects of prebiotics and probiotics on the faecal microbi-ota and selected immune parameters relevant to the elderly. PLoS ONE 2016, 11, e0162604. [Google Scholar] [CrossRef]
- Min, F.-F.; Hu, J.-L.; Nie, S.; Xie, J.-H.; Xie, M. In vitro fermentation of the polysaccharides from Cyclocarya paliurus leaves by human fecal inoculums. Carbohydr. Polym. 2014, 112, 563–568. [Google Scholar] [CrossRef]
- Rodrigues, D.; Walton, G.; Sousa, S.; Rocha-Santos, T.A.; Duarte, A.C.; Freitas, A.C.; Gomes, A.M.P. In vitro fermentation and prebiotic potential of selected extracts from seaweeds and mushrooms. LWT 2016, 73, 131–139. [Google Scholar] [CrossRef]
- Shen, Q.; Tuohy, K.; Gibson, G.; Ward, R.E. In vitro measurement of the impact of human milk oligosaccharides on the faecal microbiota of weaned formula-fed infants compared to a mixture of prebiotic fructooligosaccharides and galactooligosaccharides. Lett. Appl. Microbiol. 2011, 52, 337–343. [Google Scholar] [CrossRef]
- Scott, K.P.; Gratz, S.W.; Sheridan, P.O.; Flint, H.J.; Duncan, S.H. The influence of diet on the gut microbiota. Pharmacol. Res. 2013, 69, 52–60. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease; Elsevier BV: Amsterdam, The Netherlands, 2014; Volume 121, pp. 91–119. [Google Scholar]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491. [Google Scholar] [CrossRef]
- Scott, K.P.; Grimaldi, R.; Cunningham, M.; Sarbini, S.R.; Wijeyesekera, A.; Tang, M.L.; Lee, J.C.-Y.; Yau, Y.F.; Ansell, J.; Theis, S.; et al. Developments in understanding and applying prebiotics in research and practice—An ISAPP conference paper. J. Appl. Microbiol. 2019, 128, 934–949. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- González, C.; González, D.; Zúñiga, R.N.; Estay, H.; Troncoso, E. Simulation of Human Small Intestinal Digestion of Starch Using an In Vitro System Based on a Dialysis Membrane Process. Foods 2020, 9, 913. [Google Scholar] [CrossRef] [PubMed]
- Mills, D.; Tuohy, K.; Booth, J.; Buck, M.; Crabbe, M.J.C.; Gibson, G.; Ames, J. Dietary glycated protein modulates the colonic microbiota towards a more detrimental composition in ulcerative colitis patients and non-ulcerative colitis subjects. J. Appl. Microbiol. 2008, 105, 706–714. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donors Information (1) | |||||||
| Donor | Genre | Height (m) | Weight (kg) | Age (years) | BMI * (kg/m2) | Dairy Consumption | Regular Exercise |
| D1 | Male | 1.82 | 97 | 25 | 29.37 | Yes | Yes |
| D2 | Female | 1.60 | 60 | 31 | 23.44 | Yes | No |
| D3 | Female | 1.76 | 81 | 35 | 26.15 | Yes | Yes |
| D4 | Male | 1.68 | 72 | 32 | 25.51 | Yes | Yes |
| D5 | Female | 1.62 | 60 | 40 | 22.86 | Yes | Yes |
| Individual Inoculums (2) | |||||||
| Donor | Individual Fecal Weight (g) | Individual Feces Plus Dilution Solution (g) | Fecal Dilution (%) | ||||
| D1 | 2.01 | 20.08 | 10.01 | ||||
| D2 | 2.01 | 20.06 | 10.02 | ||||
| D3 | 2.16 | 20.20 | 10.69 | ||||
| D4 | 2.21 | 20.02 | 11.04 | ||||
| D5 | 2.19 | 20.07 | 10.91 | ||||
| Pooled Inoculum-P Fresh (3) | |||||||
| Donor | Individual Fecal Weight (g) | Fecal Pooled Weight (g) | Fecal Pooled Plus Dilution Solution (g) | Fecal Dilution (%) | Individual Representation (%) | ||
| D1 | 1.23 | 5.48 | 54.86 | 9.99 | 22.45 | ||
| D2 | 1.15 | 20.99 | |||||
| D3 | 1.11 | 20.26 | |||||
| D4 | 0.76 | 13.87 | |||||
| D5 | 1.23 | 22.45 | |||||
| Pooled Inoculum (1) | ||||||
| Donor | Individual Fecal Weight (g) | Fecal Pool Weight (g) | Individual Representation (%) | |||
| D1 | 2.2 | 25.1 | 13.94 | |||
| D2 | 3.5 | 8.76 | ||||
| D3 | 4.3 | 17.13 | ||||
| D4 | 7.0 | 27.89 | ||||
| D5 | 8.1 | 32.27 | ||||
| Pooled Inoculum Preparation Conditions (2) | ||||||
| Condition | Fecal Pool Weight (g) | Fecal Pooled Plus Dilution Solution (g) | Fecal Dilution (%) | Storage Solution | Storage Time at −20 °C (Days) | Fecal Pool Diluted in |
| I | 6.7 | 67.4 | 9.94 | --- | --- | 0.1 M PBS |
| II | 6.7 | 67.4 | 9.94 | 0.1 M PBS | 15 | 0.1 M PBS |
| III | 2.8 | 28.3 | 9.89 | --- | 15 | 0.1 M PBS |
| IV | 6.6 | 66.4 | 9.94 | 0.1 M PBS + 30% glycerol | 15 and 90 | 0.1 M PBS + 30% glycerol |
| Microorganism Enumeration of Fresh Individual (D1–D5) and P Fresh Inoculums (1) | ||
|---|---|---|
| Culture Media | Incubation Conditions | Target Bacteria Group |
| Bifidus selective medium agar (BSMA) | Anaerobic 37 °C for 72 h | Bifidobacteria |
| Columbia blood agar (CBA) with 5% (v/v) defibrinated sheep blood | Total bacteria (included fastidious microorganisms) | |
| MacConkey agar (MCA) | Gram negative and enteric bacteria (e.g., Escherichia coli and Salmonella spp.) | |
| de Man, Rogosa and Sharpe agar (MRSA) | Acid lactic bacteria (LAB) | |
| Violet red bile glucose agar (VRBGA) | Enterobacteriaceae (e.g., Escherichia coli and Salmonella spp.) | |
| Inoculums Submitted to Conditions I–IV and Inoculums A and B (2) | ||
| Culture Media | Incubation Conditions | Target Bacteria Group |
| Columbia blood agar (CBA) with 5% (v/v) defibrinated sheep blood | Anaerobic37 °Cfor 72 h | Total bacteria (included fastidious microorganisms) |
| MacConkey agar (MCA) | Gram negative and enteric bacteria (e.g., Escherichia coli and Salmonella spp.) | |
| de Man, Rogosa and Sharpe agar (MRSA) with 0.1% (w/v) cysteine | Acid lactic bacteria (LAB) and Bifidobacteria | |
| Condition | pH |
|---|---|
| D1 | 6.58 |
| D2 | 6.98 |
| D3 | 6.97 |
| D4 | 7.07 |
| D5 | 7.31 |
| Average * | 6.98 ± 0.24 |
| P fresh | 6.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Carvalho, N.M.; Oliveira, D.L.; Dib Saleh, M.A.; Pintado, M.; Madureira, A.R. Preservation of Human Gut Microbiota Inoculums for In Vitro Fermentations Studies. Fermentation 2021, 7, 14. https://doi.org/10.3390/fermentation7010014
de Carvalho NM, Oliveira DL, Dib Saleh MA, Pintado M, Madureira AR. Preservation of Human Gut Microbiota Inoculums for In Vitro Fermentations Studies. Fermentation. 2021; 7(1):14. https://doi.org/10.3390/fermentation7010014
Chicago/Turabian Stylede Carvalho, Nelson Mota, Diana Luazi Oliveira, Mayra Anton Dib Saleh, Manuela Pintado, and Ana Raquel Madureira. 2021. "Preservation of Human Gut Microbiota Inoculums for In Vitro Fermentations Studies" Fermentation 7, no. 1: 14. https://doi.org/10.3390/fermentation7010014
APA Stylede Carvalho, N. M., Oliveira, D. L., Dib Saleh, M. A., Pintado, M., & Madureira, A. R. (2021). Preservation of Human Gut Microbiota Inoculums for In Vitro Fermentations Studies. Fermentation, 7(1), 14. https://doi.org/10.3390/fermentation7010014