The Effects of Halogenated Compounds on the Anaerobic Digestion of Macroalgae




Abstract
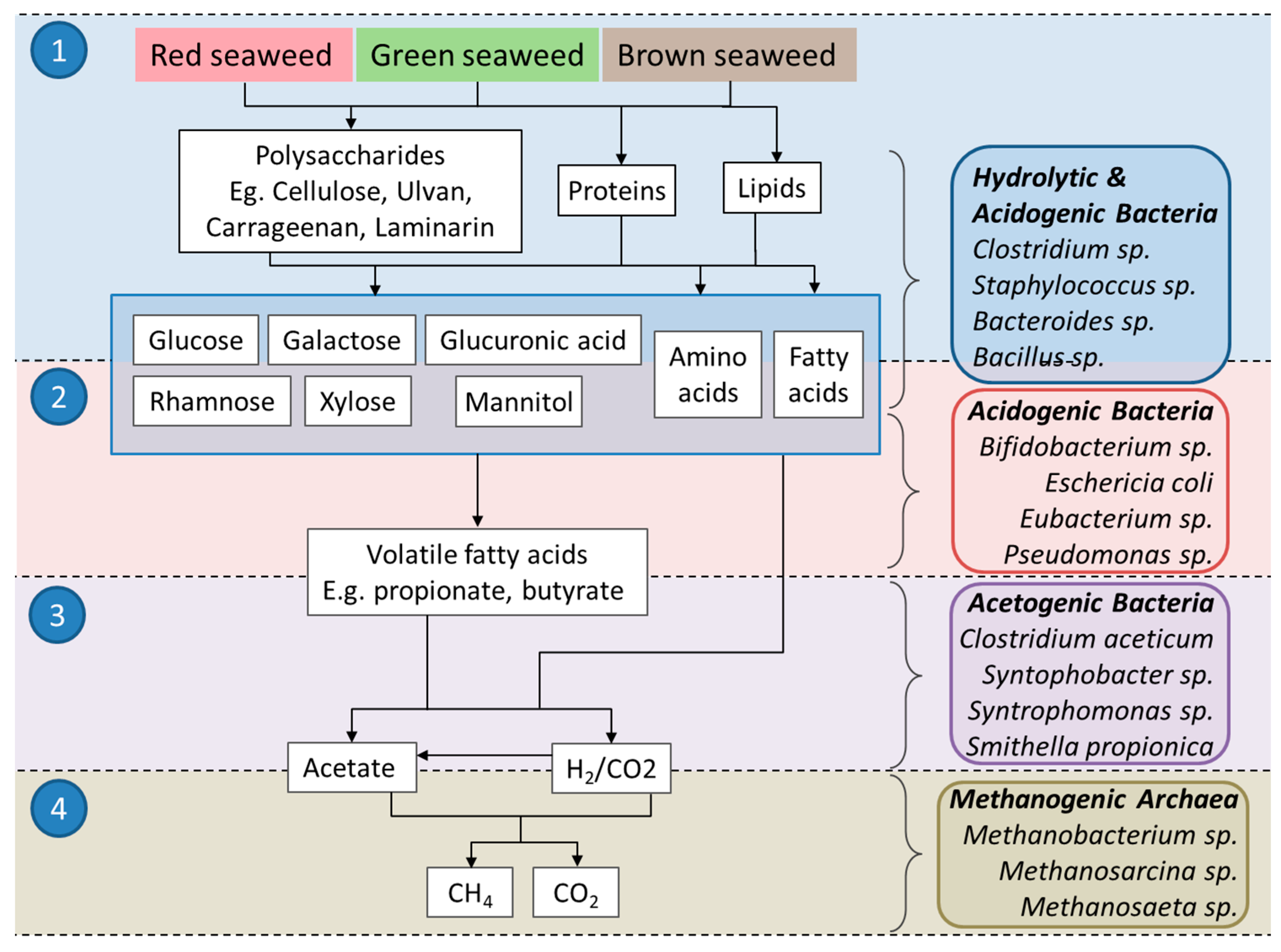
1. Introduction
2. Inhibitors of AD by Halogenated Organic Compounds
2.1. Chlorinated Organic Inhibitors of Anaerobic Digestion
2.2. Brominated Organic Inhibitors of Anaerobic Digestion
2.3. Iodinated and Fluorinated Organic Inhibitors of Anaerobic Digestion
3. Anaerobic Dehalogenation
4. Halogenated Compounds in Macroalgae and Their Effect on AD
4.1. Iodinated Compounds
4.2. Brominated Compounds
4.3. Chlorinated Compounds
4.4. Fluorinated Compounds
5. Bioremediation as Part of Refining Macroalgae for Biofuel Production and Its Challenges
6. Conclusions
Funding
Conflicts of Interest
References
- Kerrison, P.D.; Stanley, M.S.; Edwards, M.D.; Black, K.D.; Hughes, A.D. The cultivation of European kelp for bioenergy: Site and species selection. Biomass Bioenergy 2015, 80, 229–242. [Google Scholar] [CrossRef]
- Kraan, S. Mass-cultivation of carbohydrate rich macroalgae, a possible solution for sustainable biofuel production. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 27–46. [Google Scholar] [CrossRef]
- Chynoweth, D.P.; Owens, J.M.; Legrand, R. Renewable methane from anaerobic digestion of biomass. Renew. Energy 2001, 22, 1–8. [Google Scholar] [CrossRef]
- Milledge, J.; Nielsen, B.; Maneein, S.; Harvey, P. A brief review of anaerobic digestion of algae for bioenergy. Energies 2019, 12, 1166. [Google Scholar] [CrossRef]
- Gao, K.; McKinley, K.R. Use of macroalgae for marine biomass production and CO2 remediation: A review. J. Appl. Phycol. 1994, 6, 45–60. [Google Scholar] [CrossRef]
- Sode, S.; Bruhn, A.; Balsby, T.J.S.; Larsen, M.M.; Gotfredsen, A.; Rasmussen, M.B. Bioremediation of reject water from anaerobically digested waste water sludge with macroalgae (Ulva lactuca, Chlorophyta). Bioresour. Technol. 2013, 146, 426–435. [Google Scholar] [CrossRef]
- Kuroda, K.; Fujii, K.; Akase, M. Experimental Study of Hydrogen Fermentation of Seaweeds for Efficient Operation. In Proceedings of the 2018 OCEANS—MTS/IEEE Kobe Techno-Oceans (OTO), Kobe, Japan, 28–31 May 2018; pp. 1–4. [Google Scholar]
- Tabassum, M.R.; Xia, A.; Murphy, J.D. Biomethane production from various segments of brown seaweed. Energy Convers. Manag. 2018, 174, 855–862. [Google Scholar] [CrossRef]
- Amani, T.; Nosrati, M.; Sreekrishnan, T.R. Anaerobic digestion from the viewpoint of microbiological, chemical, and operational aspects—A review. Environ. Rev. 2010, 18, 255–278. [Google Scholar] [CrossRef]
- Sun, W.; Yu, G.; Louie, T.; Liu, T.; Zhu, C.; Xue, G.; Gao, P. From mesophilic to thermophilic digestion: The transitions of anaerobic bacterial, archaeal, and fungal community structures in sludge and manure samples. Appl. Microbiol. Biotechnol. 2015, 99, 10271–10282. [Google Scholar] [CrossRef]
- Prabhakaran, P.; Bhasi, A.; Ali, S.; Narayanan, N.; Balakrishnan, M.V.; Bhaskaran, K. Community dynamics and significance of anaerobic protozoa during biomethanation of lignocellulosic waste. Renew. Energy 2016, 98, 148–152. [Google Scholar] [CrossRef]
- Zhang, C.; Su, H.; Baeyens, J.; Tan, T. Reviewing the anaerobic digestion of food waste for biogas production. Renew. Sustain. Energy Rev. 2014, 38, 383–392. [Google Scholar] [CrossRef]
- Appels, L.; Van Assche, A.; Willems, K.; Degrève, J.; Van Impe, J.; Dewil, R. Peracetic acid oxidation as an alternative pre-treatment for the anaerobic digestion of waste activated sludge. Bioresour. Technol. 2011, 102, 4124–4130. [Google Scholar] [CrossRef] [PubMed]
- Parkin, G.F.; Owen, W.F. Fundamentals of Anaerobic Digestion of Wastewater Sludges. J. Environ. Eng. 1986, 112, 867–920. [Google Scholar] [CrossRef]
- Vanegas, C.H.; Bartlett, J. Green energy from marine algae: Biogas production and composition from the anaerobic digestion of Irish seaweed species. Environ. Technol. 2013, 34, 2277–2283. [Google Scholar] [CrossRef] [PubMed]
- Peu, P.; Sassi, J.-F.; Girault, R.; Picard, S.; Saint-Cast, P.; Béline, F.; Dabert, P. Sulphur fate and anaerobic biodegradation potential during co-digestion of seaweed biomass (Ulva sp.) with pig slurry. Bioresour. Technol. 2011, 102, 10794–10802. [Google Scholar] [CrossRef]
- Maneein, S.; Milledge, J.J.; Nielsen, V.B.; Harvey, J.P. A review of seaweed pre-treatment methods for enhanced biofuel production by anaerobic digestion or fermentation. Fermentation 2018, 4, 100. [Google Scholar] [CrossRef]
- Sutherland, A.D.; Varela, J.C. Comparison of various microbial inocula for the efficient anaerobic digestion of Laminaria hyperborea. BMC Biotechnol. 2014, 14, 7. [Google Scholar] [CrossRef]
- Milledge, J.; Nielsen, B.; Harvey, P. The inhibition of anaerobic digestion by model phenolic compounds representative of those from Sargassum muticum. J. Appl. Phycol. 2018. [Google Scholar] [CrossRef]
- Jha, P.; Schmidt, S. Reappraisal of chemical interference in anaerobic digestion processes. Renew. Sustain. Energy Rev. 2017, 75, 954–971. [Google Scholar] [CrossRef]
- Chen, J.L.; Ortiz, R.; Steele, T.W.J.; Stuckey, D.C. Toxicants inhibiting anaerobic digestion: A review. Biotechnol. Adv. 2014, 32, 1523–1534. [Google Scholar] [CrossRef]
- Wang, H.; Fotidis, I.A.; Angelidaki, I. Ammonia effect on hydrogenotrophic methanogens and syntrophic acetate-oxidizing bacteria. FEMS Microbiol. Ecol. 2015, 91, 5512–5520. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Smith, G.B. Inhibition of methanogenesis by C1- and C2-polychlorinated aliphatic hydrocarbons. Environ. Toxicol. Chem. 2000, 19, 2212–2217. [Google Scholar] [CrossRef]
- Van Beelen, P.; van Vlaardingen, P. Toxic effects of pollutants on the mineralization of 4-chlorophenol and benzoate in methanogenic river sediment. Environ. Toxicol. Chem. 1994, 13, 1051–1060. [Google Scholar] [CrossRef]
- Weathers, L.J.; Parkin, G.F. Toxicity of chloroform biotransformation to methanogenic bacteria. Environ. Sci. Technol. 2000, 34, 2764–2767. [Google Scholar] [CrossRef]
- Bagley, D.M.; Lalonde, M.; Kaseros, V.; Stasiuk, K.E.; Sleep, B.E. Acclimation of anaerobic systems to biodegrade tetrachloroethene in the presence of carbon tetrachloride and chloroform. Water Res. 2000, 34, 171–178. [Google Scholar] [CrossRef]
- Maymó-Gatell, X.; Nijenhuis, I.; Zinder, S.H. Reductive dechlorination of cis-1,2-dichloroethene and vinyl chloride by “Dehalococcoides ethenogenes”. Environ. Sci. Technol. 2001, 35, 516–521. [Google Scholar] [CrossRef]
- Gunsalus, R.P.; Wolfe, R.S. ATP activation and properties of the methyl coenzyme M reductase system in Methanobacterium thermoautotrophicum. J. Bacteriol. 1978, 135, 851–857. [Google Scholar] [CrossRef]
- Lian, S.; Nikolausz, M.; Nijenhuis, I.; da Rocha, U.N.; Liu, B.; Corrêa, F.B.; Saraiva, J.P.; Richnow, H.H. Biotransformation of hexachlorocyclohexanes contaminated biomass for energetic utilization demonstrated in continuous anaerobic digestion system. J. Hazard. Mater. 2020, 384, 121448. [Google Scholar] [CrossRef]
- Sanz, J.L.; Rodríguez, N.; Amils, R. Effect of chlorinated aliphatic hydrocarbons on the acetoclastic methanogenic activity of granular sludge. Appl. Microbiol. Biotechnol. 1997, 47, 324–328. [Google Scholar] [CrossRef]
- Patel, G.B.; Agnew, B.J.; Dicaire, C.J. Inhibition of pure cultures of methanogens by benzene ring compounds. Appl. Environ. Microbiol. 1991, 57, 2969–2974. [Google Scholar] [CrossRef]
- Liu, Y.; Tay, J.-H. Strategy for minimization of excess sludge production from the activated sludge process. Biotechnol. Adv. 2001, 19, 97–107. [Google Scholar] [CrossRef]
- Sikkema, J.; de Bont, J.A.; Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Rev. 1995, 59, 201–222. [Google Scholar] [CrossRef]
- Heipieper, H.J.; Weber, F.J.; Sikkema, J.; Keweloh, H.; de Bont, J.A.M. Mechanisms of resistance of whole cells to toxic organic solvents. Trends Biotechnol. 1994, 12, 409–415. [Google Scholar] [CrossRef]
- Sierra-Alvarez, R.; Lettinga, G. The effect of aromatic structure on the inhibition of acetoclastic methanogenesis in granular sludge. Appl. Microbiol. Biotechnol. 1991, 34, 544–550. [Google Scholar] [CrossRef]
- Uberoi, V.; Bhattacharya, S.K. Effects of chlorophenols and nitrophenols on the kinetics of propionate degradation in sulfate-reducing anaerobic systems. Environ. Sci. Technol. 1997, 31, 1607–1614. [Google Scholar] [CrossRef]
- Ennik-Maarsen, K.A.; Louwerse, A.; Roelofsen, W.; Stams, A.J.M. Influence of monochlorophenols on methanogenic activity in granular sludge. Water Res. 1998, 32, 2977–2982. [Google Scholar] [CrossRef]
- Ali, S.S.; Kornaros, M.; Manni, A.; Sun, J.; El-Shanshoury, A.E.-R.R.; Kenawy, E.-R.; Khalil, M.A. Enhanced anaerobic digestion performance by two artificially constructed microbial consortia capable of woody biomass degradation and chlorophenols detoxification. J. Hazard. Mater. 2020, 389, 122076. [Google Scholar] [CrossRef]
- Ali, S.S.; Abomohra, A.E.-F.; Sun, J. Effective bio-pretreatment of sawdust waste with a novel microbial consortium for enhanced biomethanation. Bioresour. Technol. 2017, 238, 425–432. [Google Scholar] [CrossRef]
- Liang, J.; Fang, X.; Lin, Y.; Wang, D. A new screened microbial consortium OEM2 for lignocellulosic biomass deconstruction and chlorophenols detoxification. J. Hazard. Mater. 2018, 347, 341–348. [Google Scholar] [CrossRef]
- Belay, N.; Daniels, L. Production of Ethane, Ethylene, and Acetylene from Halogenated Hydrocarbons by Methanogenic Bacteria. Appl. Environ. Microbiol. 1987, 53, 1604–1610. [Google Scholar] [CrossRef]
- Denman, S.E.; Tomkins, N.W.; McSweeney, C.S. Quantitation and diversity analysis of ruminal methanogenic populations in response to the antimethanogenic compound bromochloromethane. FEMS Microbiol. Ecol. 2007, 62, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Abecia, L.; Toral, P.G.; Martín-García, A.I.; Martínez, G.; Tomkins, N.W.; Molina-Alcaide, E.; Newbold, C.J.; Yáñez-Ruiz, D.R. Effect of bromochloromethane on methane emission, rumen fermentation pattern, milk yield, and fatty acid profile in lactating dairy goats. J. Dairy Sci. 2012, 95, 2027–2036. [Google Scholar] [CrossRef]
- Healy, J.B.; Young, L.Y.; Reinhard, M. Methanogenic decomposition of ferulic Acid, a model lignin derivative. Appl. Environ. Microbiol. 1980, 39, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Zinder, S.H.; Anguish, T.; Cardwell, S.C. Selective Inhibition by 2-Bromoethanesulfonate of Methanogenesis from Acetate in a Thermophilic Anaerobic Digestor. Appl. Environ. Microbiol. 1984, 47, 1343–1345. [Google Scholar] [CrossRef]
- Bouwer, E.J.; McCarty, P.L. Effects of 2-bromoethanesulfonic Acid and 2- chloroethanesulfonic Acid on acetate utilization in a continuous-flow methanogenic fixed-film column. Appl. Environ. Microbiol. 1983, 45, 1408–1410. [Google Scholar] [CrossRef] [PubMed]
- Gerecke, A.C.; Hartmann, P.C.; Heeb, N.V.; Kohler, H.-P.E.; Giger, W.; Schmid, P.; Zennegg, M.; Kohler, M. Anaerobic degradation of decabromodiphenyl ether. Environ. Sci. Technol. 2005, 39, 1078–1083. [Google Scholar] [CrossRef]
- Shin, M.; Duncan, B.; Seto, P.; Falletta, P.; Lee, D.-Y. Dynamics of selected pre-existing polybrominated diphenylethers (PBDEs) in municipal wastewater sludge under anaerobic conditions. Chemosphere 2010, 78, 1220–1224. [Google Scholar] [CrossRef]
- Shi, C.; Hu, Y.; Kobayashi, T.; Zhang, N.; Kuramochi, H.; Zhang, Z.; Xu, K.-Q. Anaerobic degradation of deca-brominated diphenyl ether contaminated in products: Effect of temperature on degradation characteristics. Bioresour. Technol. 2019, 283, 28–35. [Google Scholar] [CrossRef]
- Lee, L.K.; He, J. Reductive debromination of polybrominated diphenyl ethers by anaerobic bacteria from soils and sediments. Appl. Environ. Microbiol. 2010, 76, 794–802. [Google Scholar] [CrossRef]
- Song, M.; Luo, C.; Li, F.; Jiang, L.; Wang, Y.; Zhang, D.; Zhang, G. Anaerobic degradation of polychlorinated biphenyls (PCBs) and Polychlorinated Biphenyls Ethers (PBDEs), and microbial community dynamics of electronic waste-contaminated soil. Sci. Total Environ. 2015, 502, 426–433. [Google Scholar] [CrossRef]
- Wu, Q.; Bedard, D.L.; Wiegel, J. Temperature determines the pattern of anaerobic microbial dechlorination of Aroclor 1260 primed by 2,3,4,6-tetrachlorobiphenyl in woods pond sediment. Appl. Environ. Microbiol. 1997, 63, 4818–4825. [Google Scholar] [CrossRef]
- Wu, Q.; Bedard, D.L.; Wiegel, J. Effect of incubation temperature on the route of microbial reductive dechlorination of 2,3,4,6-tetrachlorobiphenyl in polychlorinated biphenyl (PCB)-contaminated and PCB-free freshwater sediments. Appl. Environ. Microbiol. 1997, 63, 2836–2843. [Google Scholar] [CrossRef]
- Harmeson, R.H.; Dietz, J. The effect of radioactive substances on sludge digestionNo Title. Eng. Exp. Stn. Bull. 1957, 441. [Google Scholar]
- Amachi, S.; Minami, K.; Miyasaka, I.; Fukunaga, S. Ability of anaerobic microorganisms to associate with iodine: 125I tracer experiments using laboratory strains and enriched microbial communities from subsurface formation water. Chemosphere 2010, 79, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Barker, H.A. Corrinoid-dependent enzymic reactions. Annu. Rev. Biochem. 1972, 41, 55–90. [Google Scholar] [CrossRef] [PubMed]
- Londry, K.L.; Fedorak, P.M. Fluorophenols and 3-fluorobenzoate in phenol-degrading methanogenic cultures. Arch. Microbiol. 1993, 160, 137–143. [Google Scholar] [CrossRef]
- Kenealy, W.; Zeikus, J.G. Influence of corrinoid antagonists on methanogen metabolism. J. Bacteriol. 1981, 146, 133–140. [Google Scholar] [CrossRef]
- Penning, H.; Conrad, R. Effect of inhibition of acetoclastic methanogenesis on growth of archaeal populations in an anoxic model environment. Appl. Environ. Microbiol. 2006, 72, 178–184. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Janssen, P.H.; Frenzel, P. Inhibition of methanogenesis by methyl fluoride: Studies of pure and defined mixed cultures of anaerobic bacteria and archaea. Appl. Environ. Microbiol. 1997, 63, 4552–4557. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.-P.; Lü, F.; He, P.-J.; Li, L.; Shao, L.-M. Quantification of the inhibitory effect of methyl fluoride on methanogenesis in mesophilic anaerobic granular systems. Chemosphere 2011, 84, 1194–1199. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Feng, Y.; Feng, H.; Ghulam, A.; Su, Y.; Shen, D. Anaerobic biotransformation of fluoronitrobenzenes and microbial communities in methanogenic systems. J. Environ. Sci. Health Part A 2014, 49, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Vargas, C.; Song, B.; Camps, M.; Häggblom, M.M. Anaerobic degradation of fluorinated aromatic compounds. Appl. Microbiol. Biotechnol. 2000, 53, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Schoberth, S. A new strain of Desulfovibrio gigas isolated from a sewage plant. Arch. Mikrobiol. 1973, 92, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Remde, A.; Debus, R. Biodegradability of fluorinated surfactants under aerobic and anaerobic conditions. Chemosphere 1996, 32, 1563–1574. [Google Scholar] [CrossRef]
- Meesters, R.J.W.; Schröder, H.F. Perfluorooctane sulfonate—A quite mobile anionic anthropogenic surfactant, ubiquitously found in the environment. Water Sci. Technol. J. Int. Assoc. Water Pollut. Res. 2004, 50, 235–242. [Google Scholar] [CrossRef]
- Wang, N.; Szostek, B.; Folsom, P.W.; Sulecki, L.M.; Capka, V.; Buck, R.C.; Berti, W.R.; Gannon, J.T. Aerobic biotransformation of 14C-labeled 8-2 telomer B alcohol by activated sludge from a domestic sewage treatment plant. Environ. Sci. Technol. 2005, 39, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Szostek, B.; Buck, R.C.; Folsom, P.W.; Sulecki, L.M.; Capka, V.; Berti, W.R.; Gannon, J.T. Fluorotelomer alcohol biodegradation-direct evidence that perfluorinated carbon chains breakdown. Environ. Sci. Technol. 2005, 39, 7516–7528. [Google Scholar] [CrossRef]
- Fetzner, S.; Lingens, F. Bacterial dehalogenases: Biochemistry, genetics, and biotechnological applications. Microbiol. Rev. 1994, 58, 641–685. [Google Scholar] [CrossRef]
- Lian, S.; Nikolausz, M.; Nijenhuis, I.; Francisco Leite, A.; Richnow, H.H. Biotransformation and inhibition effects of hexachlorocyclohexanes during biogas production from contaminated biomass characterized by isotope fractionation concepts. Bioresour. Technol. 2018, 250, 683–690. [Google Scholar] [CrossRef]
- Allard, A.-S.; Neilson, A.H. Degradation and Transformation of Organic Bromine and Iodine Compounds: Comparison with Their Chlorinated Analogues BT—Organic Bromine and Iodine Compounds; Neilson, A.H., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 1–74. [Google Scholar]
- DeWeerd, K.A.; Suflita, J.M. Anaerobic Aryl Reductive Dehalogenation of Halobenzoates by Cell Extracts of “Desulfomonile tiedjei”. Appl. Environ. Microbiol. 1990, 56, 2999–3005. [Google Scholar] [CrossRef]
- Tsang, J.S.H.; Sallis, P.J.; Bull, A.T.; Hardman, D.J. A monobromoacetate dehalogenase from Pseudomonas cepacia MBA4. Arch. Microbiol. 1988, 150, 441–446. [Google Scholar] [CrossRef]
- Barrio-Lage, G.; Parsons, F.Z.; Nassar, R.S.; Lorenzo, P.A. Sequential dehalogenation of chlorinated ethenes. Environ. Sci. Technol. 1986, 20, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; McCarty, P.L. Competition for Hydrogen within a Chlorinated Solvent Dehalogenating Anaerobic Mixed Culture. Environ. Sci. Technol. 1998, 32, 3591–3597. [Google Scholar] [CrossRef]
- Teng, Y.; Xu, Y.; Wang, X.; Christie, P. Function of Biohydrogen Metabolism and Related Microbial Communities in Environmental Bioremediation. Front. Microbiol. 2019, 10, 106. [Google Scholar] [CrossRef]
- Sanford, R.A.; Cole, J.R.; Tiedje, J.M. Characterization and Description of Anaeromyxobacter dehalogenans gen. nov., sp. nov., an Aryl-Halorespiring Facultative Anaerobic Myxobacterium. Appl. Environ. Microbiol. 2002, 68, 893–900. [Google Scholar] [CrossRef]
- Sun, B.; Cole, J.R.; Sanford, R.A.; Tiedje, J.M. Isolation and characterization of desulfovibrio dechloracetivorans sp. nov., a marine dechlorinating bacterium growing by coupling the oxidation of acetate to the reductive dechlorination of 2-chlorophenol. Appl. Environ. Microbiol. 2000, 66, 2408–2413. [Google Scholar] [CrossRef]
- Susarla, S.; Masunaga, S.; Yonezawa, Y. Biotransformation of halogenated benzenes in anaerobic sediments. Environ. Sci. Pollut. Res. 1996, 3, 71–74. [Google Scholar] [CrossRef]
- Paul, N.A.; de Nys, R.; Steinberg, P.D. Chemical defence against bacteria in the red alga Asparagopsis armata: Linking structure with function. Mar. Ecol. Prog. Ser. 2006, 306, 87–101. [Google Scholar] [CrossRef]
- Jesus, A.; Correia-da-silva, M.; Afonso, C.; Pinto, M.; Cidade, H. Isolation and potential biological applications of haloaryl secondary metabolites from macroalgae. Mar. Drugs 2019, 17, 73. [Google Scholar] [CrossRef]
- Cabrita, M.T.; Vale, C.; Rauter, A.P. Halogenated compounds from marine algae. Mar. Drugs 2010, 8, 2301–2317. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Srebnik, M. Natural halogenated fatty acids: Their analogues and derivatives. Prog. Lipid Res. 2002, 41, 315–367. [Google Scholar] [CrossRef]
- Mehrtens, G.; Laturnus, F. Halogenating activity in an arctic population of brown macroalga Laminaria saccharina (L.) Lamour. Polar Res. 1997, 16, 19–26. [Google Scholar] [CrossRef]
- Leblanc, C.; Colin, C.; Cosse, A.; Delage, L.; La Barre, S.; Morin, P.; Fiévet, B.; Voiseux, C.; Ambroise, Y.; Verhaeghe, E.; et al. Iodine transfers in the coastal marine environment: The key role of brown algae and of their vanadium-dependent haloperoxidases. Biochimie 2006, 88, 1773–1785. [Google Scholar] [CrossRef] [PubMed]
- Wever, R.; Plat, H.; de Boer, E. Isolation procedure and some properties of the bromoperoxidase from the seaweed Ascophyllum nodosum. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1985, 830, 181–186. [Google Scholar] [CrossRef]
- Krenn, B.E.; Plat, H.; Wever, R. The bromoperoxidase from the red alga Ceramium rubrum also contains vanadium as a prosthetic group. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1987, 912, 287–291. [Google Scholar] [CrossRef]
- Almeida, M.G.; Humanes, M.; Melo, R.; Silva, J.A.; da Silva, J.J.; Wever, R. Purification and characterisation of vanadium haloperoxidases from the brown alga Pelvetia canaliculata. Phytochemistry 2000, 54, 5–11. [Google Scholar] [CrossRef]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef]
- Küpper, F.C.; Miller, E.P.; Andrews, S.J.; Hughes, C.; Carpenter, L.J.; Meyer-Klaucke, W.; Toyama, C.; Muramatsu, Y.; Feiters, M.C.; Carrano, C.J. Emission of volatile halogenated compounds, speciation and localization of bromine and iodine in the brown algal genome model Ectocarpus siliculosus. J. Biol. Inorg. Chem. 2018, 23, 1119–1128. [Google Scholar] [CrossRef]
- Czatzkowska, M.; Harnisz, M. Inhibitors of the methane fermentation process with particular emphasis on the microbiological aspect: A review. Energy Sci. Eng. 2020, 8, 1880–1897. [Google Scholar] [CrossRef]
- Leri, A.C.; Dunigan, M.R.; Wenrich, R.L.; Ravel, B. Particulate organohalogens in edible brown seaweeds. Food Chem. 2019, 272, 126–132. [Google Scholar] [CrossRef]
- La Barre, S.; Potin, P.; Leblanc, C.; Delage, L. The halogenated metabolism of brown algae (phaeophyta), its biological importance and its environmental significance. Mar. Drugs 2010, 8, 988–1010. [Google Scholar] [CrossRef] [PubMed]
- Wever, R.; van der Horst, M.A. The role of vanadium haloperoxidases in the formation of volatile brominated compounds and their impact on the environment. Dalton Trans. 2013, 42, 11778–11786. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N.; Shinya, M. Seasonal evolution of bromomethanes from coralline algae (Corallinaceae) and its effect on atmospheric ozone. Mar. Chem. 1994, 45, 95–103. [Google Scholar] [CrossRef]
- Nightingale, P.D.; Malin, G.; Liss, P.S. Production of chloroform and other low molecular-weight halocarbons by some species of macroalgae. Limnol. Oceanogr. 1995, 40, 680–689. [Google Scholar] [CrossRef]
- Gall, E.; Küpper, F.; Kloareg, B. A survey of iodine content in Laminaria digitata. Bot. Mar. 2004, 47, 30–37. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Crawford, B.A.; Cowell, C.T.; Emder, P.J.; Learoyd, D.L.; Chua, E.L.; Sinn, J.; Jack, M.M. Iodine toxicity from soy milk and seaweed ingestion is associated with serious thyroid dysfunction. Med. J. Aust. 2010, 193, 413–415. [Google Scholar] [CrossRef]
- Zava, T.T.; Zava, D.T. Assessment of Japanese iodine intake based on seaweed consumption in Japan: A literature-based analysis. Thyroid Res. 2011, 4, 14. [Google Scholar] [CrossRef]
- Hou, X.; Chai, C.; Qian, Q.; Yan, X.; Fan, X. Determination of chemical species of iodine in some seaweeds (I). Sci. Total Environ. 1997, 204, 215–221. [Google Scholar] [CrossRef]
- Teas, J.; Pino, S.; Critchley, A.; Braverman, L.E. Variability of iodine content in common commercially available edible seaweeds. Thyroid 2004, 14, 836–841. [Google Scholar] [CrossRef]
- Scott, R. Observations on the Iodo-Amino-Acids of Marine Algae using Iodine-131. Nature 1954, 173, 1098–1099. [Google Scholar] [CrossRef]
- Rosell, K.-G.; Srivastava, L.M. Binding of inorganic elements to kelp residues. Hydrobiology 1984, 116–117, 505–509. [Google Scholar] [CrossRef]
- Han, L.; Fan, X.; Xiancui, L. Studies on organic iodine in seaweed II. The states and content of organic iodine in seaneed. Stud. Mar. Sin. 2001, 43, 129–135. [Google Scholar]
- Domínguez-González, M.R.; Chiocchetti, G.M.; Herbello-Hermelo, P.; Vélez, D.; Devesa, V.; Bermejo-Barrera, P. Evaluation of iodine bioavailability in seaweed using in vitro methods. J. Agric. Food Chem. 2017, 65, 8435–8442. [Google Scholar] [CrossRef]
- Wong, M.H.; Cheung, Y.H. Gas production and digestion efficiency of sewage sludge containing elevated toxic metals. Bioresour. Technol. 1995, 54, 261–268. [Google Scholar] [CrossRef]
- Meguro, H.; Abe, T.; Ogsawara, T.; Tuzimura, K. Analytical studies of iodine in food substances. Agric. Biol. Chem. 1967, 31, 999–1002. [Google Scholar]
- Van Netten, C.; Hoption Cann, S.A.; Morley, D.R.; van Netten, J.P. Elemental and radioactive analysis of commercially available seaweed. Sci. Total Environ. 2000, 255, 169–175. [Google Scholar] [CrossRef]
- Lee, S.M.; Lewis, J.; Buss, D.H.; Holcombe, G.D.; Lawrance, P.R. Iodine in British foods and diets. Br. J. Nutr. 1994, 72, 435–446. [Google Scholar] [CrossRef]
- Romarís-hortas, V.; Moreda-pi, A.; Bermejo-barrera, P. Microwave assisted extraction of iodine and bromine from edible seaweed for inductively coupled plasma-mass spectrometry determination. Talanta 2009, 79, 947–952. [Google Scholar] [CrossRef]
- Jensen, A.; Haug, A. Geographical and Seasonal Variation in the Chemical Composition of Laminaria hyperborea and Laminaria digitata from the Norwegian Coast; Akademisk Trykningssentral: Oslo, Norway, 1956; Volume 14, pp. 1–8. [Google Scholar]
- Bruhn, A.; Brynning, G.; Johansen, A.; Lindegaard, M.S.; Sveigaard, H.H.; Aarup, B.; Fonager, L.; Andersen, L.L.; Rasmussen, M.B.; Larsen, M.M.; et al. Fermentation of sugar kelp (Saccharina latissima)—Effects on sensory properties, and content of minerals and metals. J. Appl. Phycol. 2019, 31, 3175–3187. [Google Scholar] [CrossRef]
- Matanjun, P.; Mohamed, S.; Mustapha, N.M.; Muhammad, K. Nutrient content of tropical edible seaweeds, Eucheuma cottonii, Caulerpa lentillifera and Sargassum polycystum. J. Appl. Phycol. 2009, 21, 75–80. [Google Scholar] [CrossRef]
- Sheppard, M.I.; Hawkins, J.L. Iodine and microbial interactions in an organic soil. J. Environ. Radioact. 1995, 29, 91–109. [Google Scholar] [CrossRef]
- Gottardi, W. Iodine and Disinfection: Theoretical Study on Mode of Action, Efficiency, Stability, and Analytical Aspects in the Aqueous System. Arch. Pharm. (Weinheim) 1999, 332, 151–157. [Google Scholar] [CrossRef]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action, and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef] [PubMed]
- Chynoweth, D.P. Renewable biomethane from land and ocean energy crops and organic wastes. HortScience 2005, 40, 283–286. [Google Scholar] [CrossRef]
- Allen, E.; Wall, D.M.; Herrmann, C.; Xia, A.; Murphy, J.D. What is the gross energy yield of third generation gaseous biofuel sourced from seaweed? Energy 2015, 81, 352–360. [Google Scholar] [CrossRef]
- Milledge, J.J.; Harvey, P.J. Ensilage and anaerobic digestion of Sargassum muticum. J. Appl. Phycol. 2016, 28, 3021–3030. [Google Scholar] [CrossRef]
- Tabassum, M.R.; Xia, A.; Murphy, J.D. Comparison of pre-treatments to reduce salinity and enhance biomethane yields of Laminaria digitata harvested in different seasons. Energy 2017, 140, 546–551. [Google Scholar] [CrossRef]
- Liu, M.; Hansen, P.E.; Lin, X. Bromophenols in marine algae and their bioactivities. Mar. Drugs 2011, 9, 1273–1292. [Google Scholar] [CrossRef]
- Lane, A.L.; Nyadong, L.; Galhena, A.S.; Shearer, T.L.; Stout, E.P.; Parry, R.M.; Kwasnik, M.; Wang, M.D.; Hay, M.E.; Fernandez, F.M.; et al. Desorption electrospray ionization mass spectrometry reveals surface-mediated antifungal chemical defense of a tropical seaweed. Proc. Natl. Acad. Sci. USA 2009, 106, 7314–7319. [Google Scholar] [CrossRef]
- Xu, N.; Fan, X.; Yan, X.; Li, X.; Niu, R.; Tseng, C.K. Antibacterial bromophenols from the marine red alga Rhodomela confervoides. Phytochemistry 2003, 62, 1221–1224. [Google Scholar] [CrossRef]
- Xu, X.; Song, F.; Wang, S.; Li, S.; Xiao, F.; Zhao, J.; Yang, Y.; Shang, S. Dibenzyl Bromophenols with Diverse Dimerization Patterns from the Brown Alga Leathesia nana. J. Nat. Prod. 2004, 1, 1661–1666. [Google Scholar] [CrossRef] [PubMed]
- Borchardt, S.A.; Allain, E.J.; Michels, J.J.; Stearns, G.W.; Kelly, F.; Mccoy, W.F. Reaction of acylated homoserine lactone bacterial signaling molecules with oxidized halogen antimicrobials reaction of acylated homoserine lactone bacterial signaling molecules with oxidized halogen antimicrobials. Appl. Environ. Microbiol. 2001, 67, 3174–3179. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.P.; Batista, D.; Dobretsov, S.; Coutinho, R. Extracts of seaweeds as potential inhibitors of quorum sensing and bacterial growth. J. Appl. Phycol. 2017, 29, 789–797. [Google Scholar] [CrossRef]
- Flodin, C.; Whitfield, F.B. 4-hydroxybenzoic acid: A likely precursor of 2,4,6-tribromophenol in Ulva lactuca. Phytochemistry 1999, 51, 249–255. [Google Scholar] [CrossRef]
- Kladi, M.; Vagias, C.; Roussis, V. Volatile halogenated metabolites from marine red algae. Phytochem. Rev. 2004, 3, 337–366. [Google Scholar] [CrossRef]
- Bisaillon, J.G.; Lepine, F.; Beaudet, R.; Sylvestre, M. Potential for carboxylation-dehydroxylation of phenolic compounds by a methanogenic consortium. Can. J. Microbiol. 1993, 39, 642–648. [Google Scholar] [CrossRef]
- King, G. Inhibition of microbial activity in marine sediments by a bromophenol from a hemichordate. Nature 1986, 323, 257–259. [Google Scholar] [CrossRef]
- Vallecillo, A.; Garcia-Encina, P.A.; Peña, M. Anaerobic biodegradability and toxicity of chlorophenols. Water Sci. Technol. 1999, 40, 161–168. [Google Scholar] [CrossRef]
- Sahoo, N.K.; Pakshirajan, K.; Ghosh, P.K. Evaluation of 4-bromophenol biodegradation in mixed pollutants system by Arthrobacter chlorophenolicus A6 in an upflow packed bed reactor. Biodegradation 2014, 25, 705–718. [Google Scholar] [CrossRef]
- Machado, L.; Magnusson, M.; Paul, N.A.; Kinley, R.; De Nys, R.; Tomkins, N. Identification of bioactives from the red seaweed Asparagopsis taxiformis that promote antimethanogenic activity in vitro. J. Appl. Phycol. 2016, 28, 3117–3126. [Google Scholar] [CrossRef]
- Machado, L.; Magnusson, M.; Paul, N.A.; de Nys, R.; Tomkins, N. Effects of marine and freshwater macroalgae on in vitro total gas and methane production. PLoS ONE 2014, 9, e85289. [Google Scholar] [CrossRef]
- Nelson, D. Research Led by Ermias Kebreab Asks If Seaweed Can Cut Methane Emissions on Dairy Farms. Available online: https://animalscience.ucdavis.edu/news/research-led-ermias-kebreab-tests-if-seaweed-cuts-methane-emissions-dairy-farms (accessed on 26 March 2020).
- Cottle, D.; Nolan, J.; Wiedemann, S. Ruminant enteric methane mitigation: A review. Anim. Prod. Sci. 2011, 51, 491–514. [Google Scholar] [CrossRef]
- Chagas, J.C.; Ramin, M.; Krizsan, S.J. In vitro evaluation of different dietary methane mitigation strategies. Animals 2019, 9, 1120. [Google Scholar] [CrossRef] [PubMed]
- McCauley, J.I.; Labeeuw, L.; Jaramillo-Madrid, A.C.; Nguyen, L.N.; Nghiem, L.D.; Chaves, A.V.; Ralph, P.J. Management of enteric methanogenesis in ruminants by algal-derived feed additives. Curr. Pollut. Rep. 2020. [Google Scholar] [CrossRef]
- Hessami, M.J.; Phang, S.M.; Sohrabipoor, J.; Zafar, F.F.; Aslanzadeh, S. The bio-methane potential of whole plant and solid residues of two species of red seaweeds: Gracilaria manilaensis and Gracilariopsis persica. Algal Res. 2019, 42, 101581. [Google Scholar] [CrossRef]
- Gómez-Ordóñez, E.; Alonso, E.; Rupérez, P. A simple ion chromatography method for inorganic anion analysis in edible seaweeds. Talanta 2010, 82, 1313–1317. [Google Scholar] [CrossRef]
- Jard, G.; Marfaing, H.; Carrère, H.; Delgenes, J.P.; Steyer, J.P.; Dumas, C. French Brittany macroalgae screening: Composition and methane potential for potential alternative sources of energy and products. Bioresour. Technol. 2013, 144, 492–498. [Google Scholar] [CrossRef]
- Konig, G.M.; Wright, A.D.; Sticher, O.; Angerhofer, C.K.; Pezzuto, J.M. Biological activities of selected marine natural products. Planta Med. 1994, 60, 532–537. [Google Scholar] [CrossRef]
- Rao Kundeti, L.S.; Ambati, S.; Srividya, G.S.; Yadav, J.S.; Kommu, N. A review on chloro substituted marine natural product, chemical examination and biological activity. Curr. Trends Biotechnol. Pharm. 2019, 13, 72–82. [Google Scholar]
- Torres, F.A.E.; Passalacqua, T.G.; Velásquez, A.M.A.; de Souza, R.A.; Colepicolo, P.; Graminha, M.A.S. New drugs with antiprotozoal activity from marine Algae: A review. Braz. J. Pharmacogn. 2014, 24, 265–276. [Google Scholar] [CrossRef]
- Vedhagiri, K.; Manilal, A.; Valliyammai, T.; Shanmughapriya, S.; Sujith, S.; Selvin, J.; Natarajaseenivasan, K. Antimicrobial potential of a marine seaweed Asparagopsis taxiformis against Leptospira javanica isolates of rodent reservoirs. Ann. Microbial. 2009, 59, 431–437. [Google Scholar] [CrossRef]
- Sanz, J.L.; Rodríguez, N.; Amils, R. The action of antibiotics on the anaerobic digestion process. Appl. Microbiol. Biotechnol. 1996, 46, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Li, X.D.; Miao, F.P.; Li, K.; Ji, N.Y. Sesquiterpenes and acetogenins from the marine red alga Laurencia okamurai. Fitoterapia 2012, 83, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Smyrniotopoulos, V.; Vagias, C.; Rahman, M.M.; Gibbons, S.; Roussis, V. Structure and antibacterial activity of brominated diterpenes from the red alga Sphaerococcus coronopifolius. Chem. Biodivers. 2010, 7, 186–195. [Google Scholar] [CrossRef]
- Smyrniotopoulos, V.; Vagias, C.; Rahman, M.M.; Gibbons, S.; Roussis, V. Ioniols I and II, tetracyclic diterpenes with antibacterial activity, from Sphaerococcus coronopifolius. Chem. Biodivers. 2010, 7, 666–676. [Google Scholar] [CrossRef]
- Lane, A.L.; Mular, L.; Drenkard, E.J.; Shearer, T.L.; Engel, S.; Fredericq, S.; Fairchild, C.R.; Prudhomme, J.; Le Roch, K.; Hay, M.E.; et al. Ecological leads for natural product discovery: Novel sesquiterpene hydroquinones from the red macroalga Peyssonnelia sp. Tetrahedron 2011, 66, 455–461. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Suzuki, M.; Abe, T.; Masuda, M. Halogenated metabolites with antibacterial activity from the Okinawan Laurencia species. Phytochemistry 2001, 58, 517–523. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Kawamoto, T.; Miwa, H.; Suzuki, M. Potent antibacterial activity of halogenated compounds against antibiotic-resistant bacteria. Planta Med. 2004, 70, 1087–1090. [Google Scholar] [CrossRef]
- Sims, J.J.; Donnell, M.S.; Leary, J.V.; Lacy, G.H. Antimicrobial agents from marine algae. Antimicrob. Agents Chemother. 1975, 7, 320–321. [Google Scholar] [CrossRef]
- Dias, D.A.; White, J.M.; Urban, S. Laurencia filiformis: Phytochemical Profiling by Conventional and HPLC-NMR Approaches. Nat. Prod. Commun. 2009, 4, 157–172. [Google Scholar] [CrossRef] [PubMed]
- König, G.M.; Wright, A.D.; Franzblau, S.G. Assessment of antimycobacterial activity of a series of mainly marine derived natural products. Planta Med. 2000, 66, 337–342. [Google Scholar] [CrossRef] [PubMed]
- El-Said, G.F.; El-Sikaily, A. Chemical composition of some seaweed from Mediterranean Sea coast, Egypt. Environ. Monit. Assess. 2013, 185, 6089–6099. [Google Scholar] [CrossRef] [PubMed]
- Masoud, M. The effect of fluoride and other ions on algae and fish of coastal water of mediterranean sea, egypt. Am. J. Environ. Sci. 2006, 2, 53–63. [Google Scholar] [CrossRef]
- Cousins, I.T.; Palm, A. Physical-Chemical Properties and Estimated Environmental Fate of Brominated and Iodinated Organic Compounds BT—Organic Bromine and Iodine Compounds; Neilson, A.H., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 301–334. ISBN 978-3-540-37055-0. [Google Scholar]
- Mohn, W.; Tiedje, J.; Mohn, W.W.; Tiedje, J.M. Microbial reductive dehalogenation. Microbiol. Rev. 1992, 56, 482–507. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. Priority Pollutant List EPA. Available online: https://www.epa.gov/sites/production/files/2015-09/documents/priority-pollutant-list-epa.pdf (accessed on 18 April 2020).
- Milledge, J.J.; Harvey, P.J. Potential process ‘hurdles’ in the use of macroalgae as feedstock for biofuel production in the British Isles. J. Chem. Technol. Biotechnol. 2016, 91, 2221–2234. [Google Scholar] [CrossRef]
- Milledge, J.J.; Thompson, E.P.; Sauvêtre, A.; Schroeder, P.; Harvey, P.J. Chapter 8—Novel developments in biological technologies for wastewater processing. In Sustainable Water and Wastewater Processing; Galanakis, C.M., Agrafioti, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 239–278. [Google Scholar]
- Da Copertino, M.S.; Tormena, T.; Seeliger, U. Biofiltering efficiency, uptake and assimilation rates of Ulva clathrata (Roth) J. Agardh (Clorophyceae) cultivated in shrimp aquaculture waste water. J. Appl. Phycol. 2009, 21, 31–45. [Google Scholar] [CrossRef]
- Robertson-Andersson, D.V.; Potgieter, M.; Hansen, J.; Bolton, J.J.; Troell, M.; Anderson, R.J.; Halling, C.; Probyn, T. Integrated seaweed cultivation on an abalone farm in South Africa. J. Appl. Phycol. 2008, 20, 579–595. [Google Scholar] [CrossRef]
- Tsagkamilis, P.; Danielidis, D.; Dring, M.J.; Katsaros, C. Removal of phosphate by the green seaweed Ulva lactuca in a small-scale sewage treatment plant (Ios Island, Aegean Sea, Greece). J. Appl. Phycol. 2010, 22, 331–339. [Google Scholar] [CrossRef]
- Tampio, E.; Marttinen, S.; Rintala, J. Liquid fertilizer products from anaerobic digestion of food waste: Mass, nutrient and energy balance of four digestate liquid treatment systems. J. Clean. Prod. 2016, 125, 22–32. [Google Scholar] [CrossRef]
- Han, W.; Clarke, W.; Pratt, S. Stabilisation of microalgae: Iodine mobilisation under aerobic and anaerobic conditions. Bioresour. Technol. 2015, 193, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Piringer, G.; Bhattacharya, S.K. Toxicity and fate of pentachlorophenol in anaerobic acidogenic systems. Water Res. 1999, 33, 2674–2682. [Google Scholar] [CrossRef]
- Rodríguez, N.; Sanz, J. Response of an anaerobic granular sludge to chlorinated aliphatic hydrocarbons in different conditions. J. Ferment. Bioeng. 1998, 86, 226–232. [Google Scholar] [CrossRef]
- Nicholson, D.K.; Woods, S.L.; Istok, J.D.; Peek, D.C. Reductive dechlorination of chlorophenols by a pentachlorophenol- acclimated methanogenic consortium. Appl. Environ. Microbiol. 1992, 58, 2280–2286. [Google Scholar] [CrossRef]
- Guthrie, M.A.; Kirsch, E.J.; Wukasch, R.F.; Grady, C.P.L. Pentachlorophenol biodegradation—II: Anaerobic. Water Res. 1984, 18, 451–461. [Google Scholar] [CrossRef]
- Boucquey, J.-B.; Renard, P.; Amerlynck, P.; Filho, P.M.; Agathos, S.N.; Naveau, H.; Nyns, E.-J. High-rate continuous biodegradation of concentrated chlorinated aliphatics by a durable enrichment of methanogenic origin under carrier-dependent conditions. Biotechnol. Bioeng. 1995, 47, 298–307. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef]


{kind=link}
| Species | Total Iodine Content (ug g−1) | Water-Soluble Iodine (%) | Reference |
|---|---|---|---|
| Laminaria japonica | 88 | [107] | |
| Laminaria japonica, Ecklonia sp., Hi- jikia jiui&orme, Undaria pinnatifida | 85–89 | [108] | |
| Laminaria japonica | 36.3 mg L−1 | 99 | [101] |
| Laminaria japonica | 2110 | [109] | |
| Laminaria japonica | 2660 | [110] | |
| Codium fragile, Ulva pertusa, Monostroma nitidum, Gracilatia corzfervoides, Sargassum Kjellmanianum, Dictyoptetis divaricata | 0.052–18.70 mg L−1 | 16–41 | [101] |
| Porphyra spp. and Palmaria spp. (red macroalgae) | 35–128 | [111] | |
| Undaria pinnatifida, Himanthalia elongate and Laminaria ochroleuca (brown macroalgae) | 63–7088 | [111] | |
| Ulva rigida | 66–137 | [111] | |
| Laminaria hyperborea and Laminaria digitata | 150–1200 | [112] | |
| Saccharina latissima | 2630 ± 1610 | [113] | |
| Sargassum polycystum and Sargassum muticum | 6.1–7.7 | [114] |
| Macroalgae | Compound | Antibacterial Activity | References |
|---|---|---|---|
| Sphaerococcus coronopifolius | Tetracyclic brominated diterpenes | Epidemic Methicillin-Resistant Staphylococcus aureus (EMRSA-16) | [149,150] |
| Coronopifoliol | |||
| Bromotetrasphaerol | |||
| Ioniols II | |||
| Peyssonnelia sp. | Peyssonoic acids A | P. bacteriolytica | [151] |
| Laurencia mariannensis, L. majuscula, L. nidific, L. intermedia | C15 acetogenin, (12E)-lembyne-A, halogenated sesquiterpene | Alcaligenes aquamarines, Azomonas agilis, Erwinia amylovora, Escherichia coli | [152] |
| L. nidific, L. intermedia, L. okamurai | Isolaurinterol | Alteromonas sp., Streptococcus sp., Staphylococcus sp., Enterococcus faecium, Enterococcus faecalis | [152,153] |
| L. majuscula, L. nidific, L. intermedia | (6R,9R,10S)-10-bromo-9-hydroxy-chamigra-2,7(14)-diene | Azobacter beijerinckii | [154] |
| L. pacifica, L. nipponica, L. okamurai, L. johnstonii | Laurinterol | S. aureus, M. smegmatis, Streptococcus sp., Staphylococcus sp., Enterococcus faecium, Enterococcus faecalis | [153,154] |
| Chrondria oppositiclada | Chondriol | S. aureus, M. smegmatis | [154] |
| L. okamurai | Allolaurinterol acetate | E. coli, S. aureus | [148] |
| L. filiformis | Pre-pacifenol | S. aureus, M. smegmatis | [152] |
| filiforminol | B. subtilis | [155] | |
| L. obtuse, L. filiformis | Allolaurinterol | Mycobacterium tuberculosis, B. megaterium, Enterococcus faecium, Staphylococcus aureus | [153,156] |
| Rhodomela confervoides | bis (2,3-dibromo-4,5-dihydroxybenzyl) ether | Pseudomonas aeruginosa, S. epidermidis, S. aureus | [124] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, B.V.; Maneein, S.; Al Farid, M.M.; Milledge, J.J. The Effects of Halogenated Compounds on the Anaerobic Digestion of Macroalgae. Fermentation 2020, 6, 85. https://doi.org/10.3390/fermentation6030085
Nielsen BV, Maneein S, Al Farid MM, Milledge JJ. The Effects of Halogenated Compounds on the Anaerobic Digestion of Macroalgae. Fermentation. 2020; 6(3):85. https://doi.org/10.3390/fermentation6030085
Chicago/Turabian StyleNielsen, Birthe V, Supattra Maneein, Md Mahmud Al Farid, and John J Milledge. 2020. "The Effects of Halogenated Compounds on the Anaerobic Digestion of Macroalgae" Fermentation 6, no. 3: 85. https://doi.org/10.3390/fermentation6030085
APA StyleNielsen, B. V., Maneein, S., Al Farid, M. M., & Milledge, J. J. (2020). The Effects of Halogenated Compounds on the Anaerobic Digestion of Macroalgae. Fermentation, 6(3), 85. https://doi.org/10.3390/fermentation6030085