

Effects of Co-Fermentation with Lactic Acid Bacteria and Yeast on Gliadin Degradation in Whole-Wheat Sourdough



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Microorganisms
2.3. Ultrasound-Assisted Fermentation
2.4. Fermentation Experiments
2.5. Proteolytic Activity Determination
2.6. Determination of Free Amino Acids
2.7. Gluten Protein Extraction
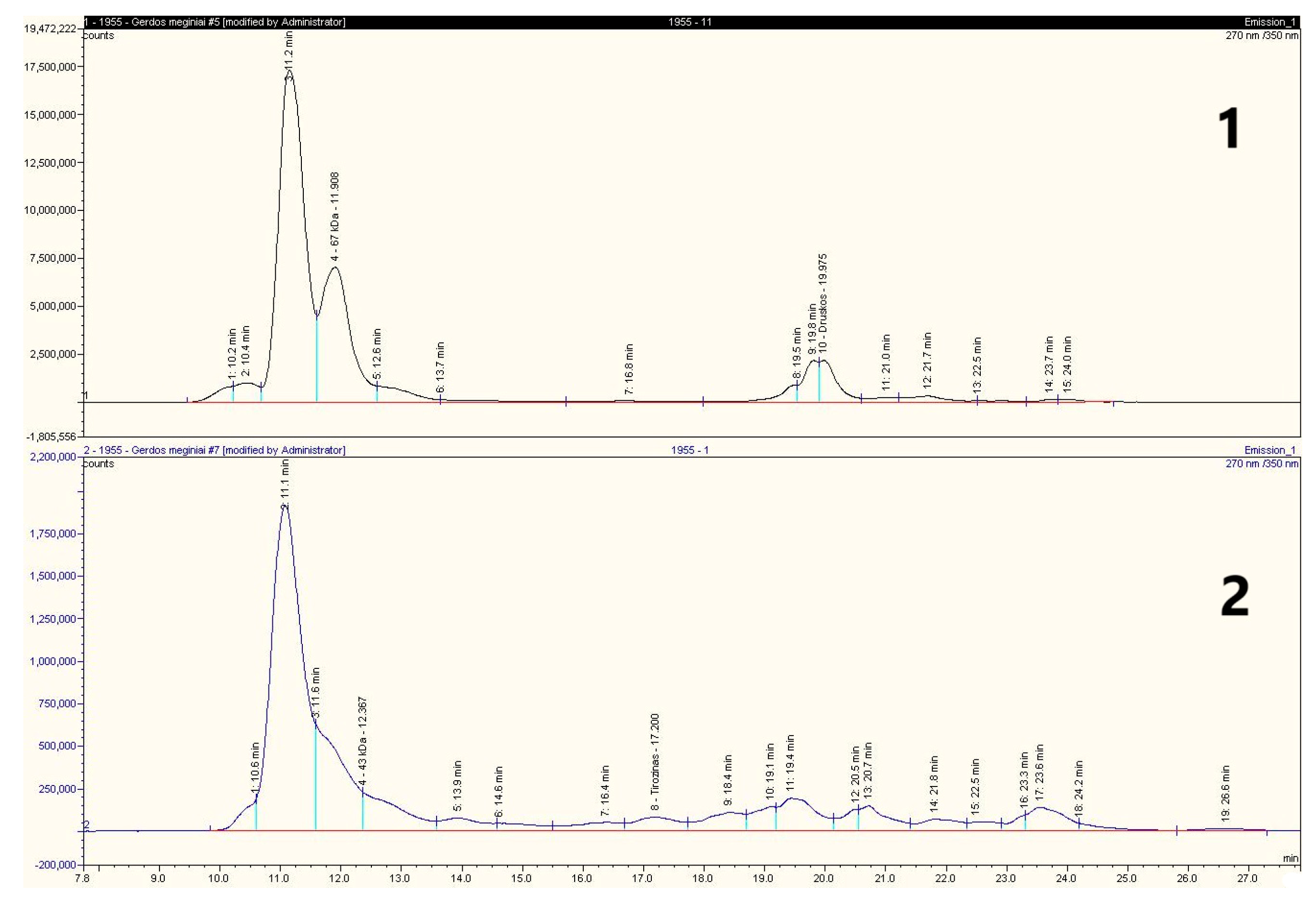
2.8. Capillary Electrophoresis
2.9. High-Performance Liquid Chromatography
2.10. Protein Electrophoresis
2.11. Analysis
2.12. Texture Analysis
2.13. Statistical Analysis
3. Results and Discussion
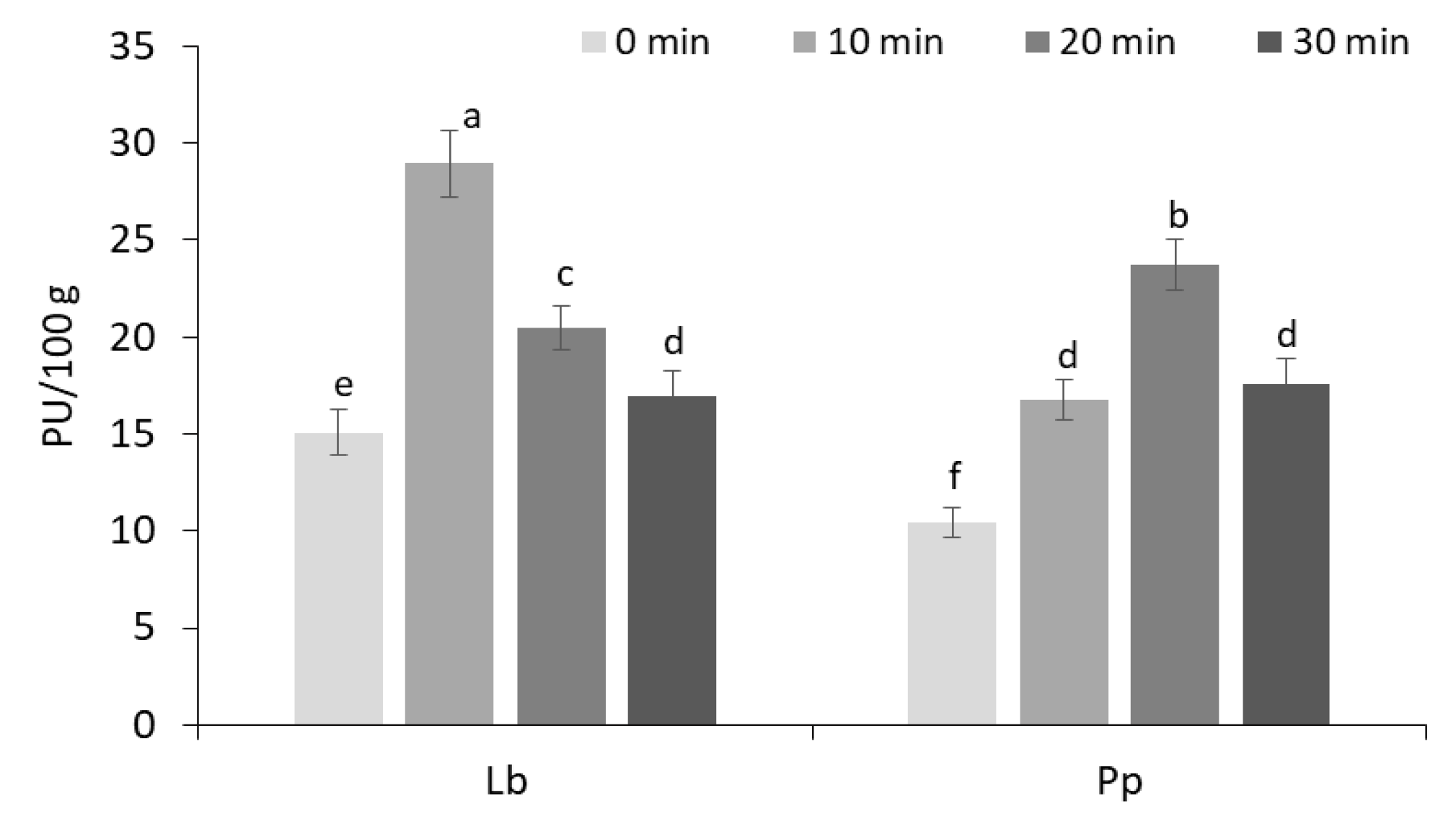
3.1. Proteolytic Activity of Lactic Acid Bacteria Strains in Wheat Wholemeal Fermentation Medium
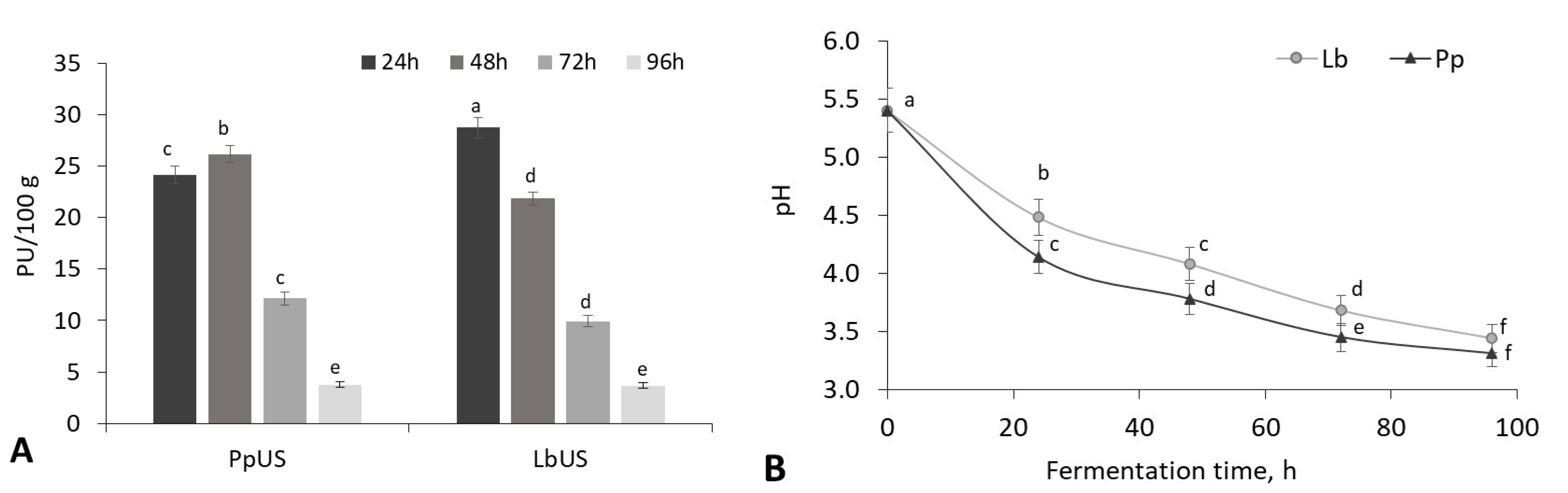
3.2. The Use of Ultrasound Technology to Activate Lactic Acid Bacteria for Sourdough Fermentation
3.3. The Effect of Ultrasound and Lactic Acid Fermentation on Pure Gluten Degradation
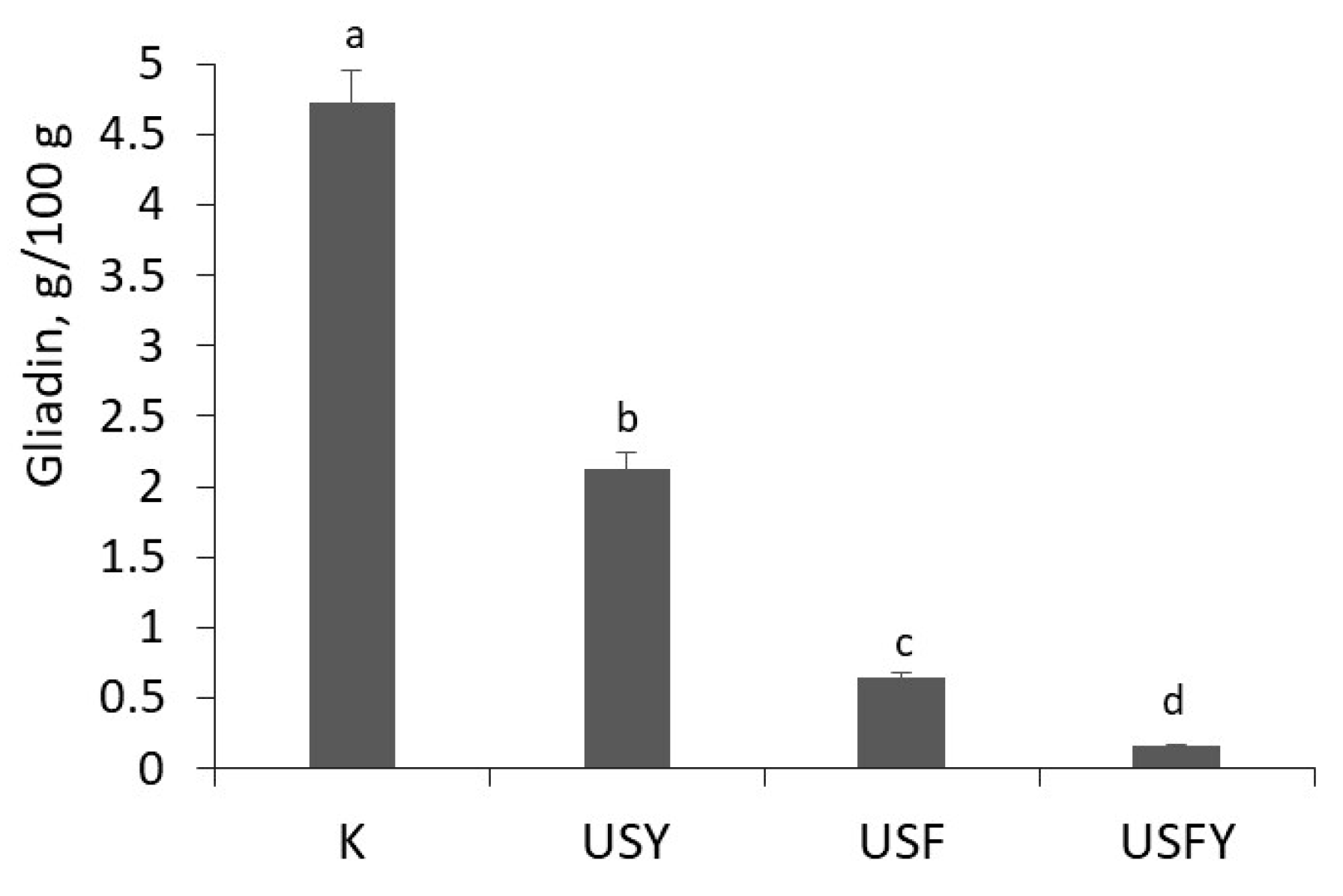
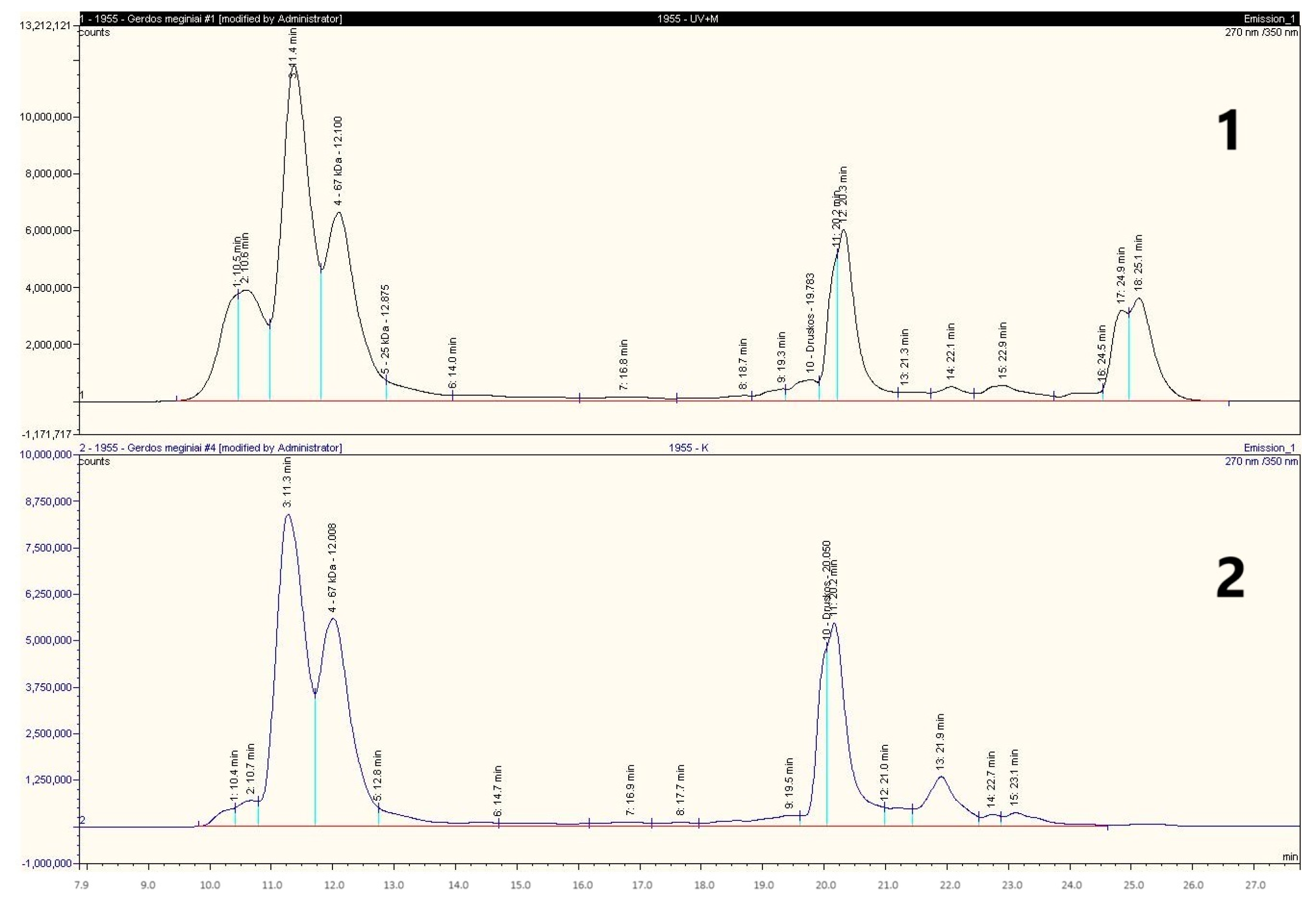
3.4. The Effect of Ultrasound on Wheat Flour Gluten Protein Degradation During Sourdough Fermentation
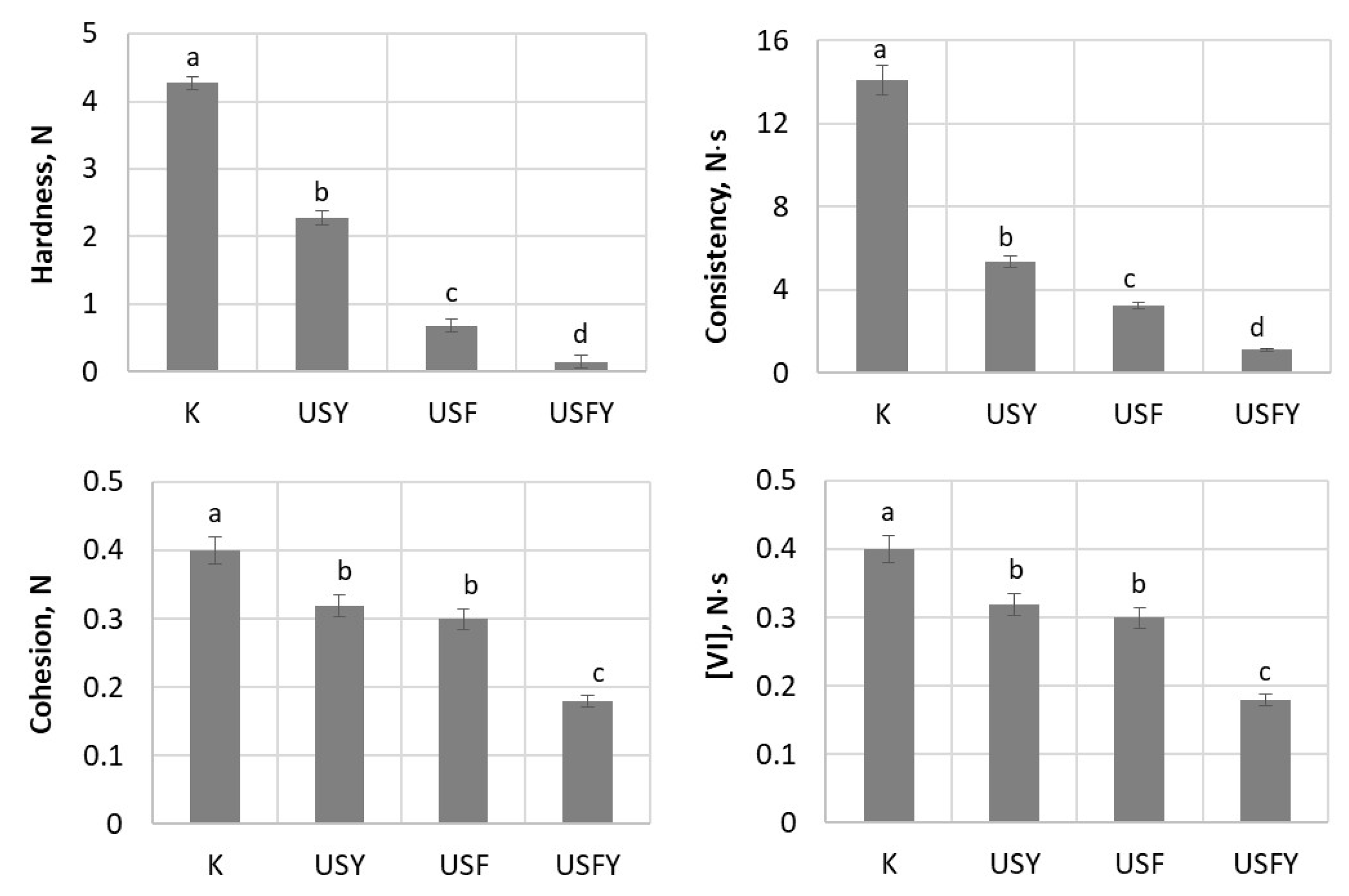
3.5. Influence of Biotechnological Tools and Ultrasound on Sourdough Texture Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gasparre, N.; Rosell, C.M. Wheat gluten: A functional protein still challenging to replace in gluten-free cereal-based foods. Cereal Chem. 2023, 100, 243–255. [Google Scholar] [CrossRef]
- Fasano, A.; Berti, I.; Gerarduzzi, T.; Not, T.; Colletti, R.B.; Drago, S.; Elitsur, Y.; Green, P.H.; Guandalini, S.; Hill, I.D.; et al. Prevalence of celiac disease in at-risk and not-at-risk groups in the United States: A large multicenter study. Arch. Intern. Med. 2012, 163, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, R.A.; Singh, T.P.; Rani, M.; Sogi, D.S. Electrophoretic characterization and proportion of different protein fractions in wheat cultivars of North-India. J. Agric. Food Res. 2021, 4, 100137. [Google Scholar] [CrossRef]
- Lee, J.; Kim, S.R.; Park, J.H.; Park, K.H.; Jeong, K.Y.; Lee, J.H.; Kang, C.S.; Kim, K.H.; Park, J.W. Evaluation of allergenicity on a ω-5 gliadin-deficient cultivar in wheat-dependent exercise-induced anaphylaxis. Allergy Asthma Immunol. Res. 2022, 14, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Dewar, D.H.; Amato, M.; Ellis, H.J.; Pollock, E.L.; Gonzalez-Cinca, N.; Wieser, H.; Ciclitira, P.J. The toxicity of high molecular weight glutenin subunits of wheat to patients with coceliac disease. Eur. J. Gastroenterol. Hepatol. 2006, 18, 483–491. [Google Scholar] [CrossRef]
- Conte, P.; Fadda, C.; Drabińska, N.; Krupa-Kozak, U. Technological and nutritional challenges, and novelty in gluten-free breadmaking: A review. Pol. J. Food Nutr. Sci. 2019, 69, 5–21. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; Auricchio, S.; Greco, L.; Clarke, C.; De Vincenzi, M.; Giovannini, C.; D’Archivio, M.; Landolfo, F.; Parrilli, G.; et al. Sourdough bread made from wheat and nontoxic flours and started with selected lactobacilli is tolerated in celiac sprue patients. Appl. Environ. Microbiol. 2010, 76, 1088–1096. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Curiel, J.A.; Nionelli, L.; Vincentini, O.; Di Cagno, R.; Silano, M.; Gobbetti, M.; Coda, R. Use of fungal proteases and selected sourdough lactic acid bacteria for making wheat bread with an intermediate content of gluten. Food Microbiol. 2014, 37, 59–68. [Google Scholar] [CrossRef]
- Arora, K.; Ameur, H.; Polo, A.; Di Cagno, R.; Rizzello, C.G.; Gobbetti, M. Thirty years of knowledge on sourdough fermentation: A systematic review. Trends Food Sci. Technol. 2021, 108, 71–83. [Google Scholar] [CrossRef]
- Leszczyńska, J.; Szczepankowska, A.K.; Majak, I.; Mańkowska, D.; Smolińska, B.; Ścieszka, S.; Diowksz, A.; Cukrowska, B.; Aleksandrzak-Piekarczyk, T. Reducing immunoreactivity of gluten peptides by probiotic lactic acid bacteria for dietary management of gluten-related diseases. Nutrients 2024, 16, 976. [Google Scholar] [CrossRef]
- Fu, W.; Jia, X.; Liu, C.; Meng, X.; Zhang, K.; Tao, S.; Xue, W. Sourdough yeast-bacteria interactions results in reduced immunogenicity by increasing depolymerization and hydrolysis of gluten. Innov. Food Sci. Emerg. Technol. 2023, 84, 103281. [Google Scholar] [CrossRef]
- Režek Jambrak, A.; Da Cruz, A.G.; Chen, J. Ultrasonic food processing as a green and sustainable technology. Ultrason. Sonochem. 2024, 106, 106880. [Google Scholar] [CrossRef] [PubMed]
- Singla, M.; Sit, N. Application of ultrasound in combination with other technologies in food processing: A review. Ultrason. Sonochem. 2021, 73, 105506. [Google Scholar] [CrossRef] [PubMed]
- Ojha, K.S.; Mason, T.J.; O’Donnell, C.P.; Kerry, J.P.; Tiwari, B.K. Ultrasound technology for food fermentation applications. Ultrason. Sonochem. 2017, 34, 410–417. [Google Scholar] [CrossRef]
- Gallo, M.; Ferrara, L.; Naviglio, D. Application of ultrasound in food science and technology: A perspective. Foods 2018, 7, 164. [Google Scholar] [CrossRef]
- Bai, J.W.; Li, Z.J.; Wang, L.F.; He, J. Ultrasonic effect on the structure and allergenicity of wheat gluten. J. Cereal Sci. 2016, 69, 138–144. [Google Scholar]
- Huang, G.; Chen, S.; Dai, C.; Sun, L.; Sun, W.; Tang, Y.; Xiong, F.; He, R.; Ma, H. Effects of ultrasound on microbial growth and enzyme activity. Ultrason. Sonochem. 2017, 37, 144–149. [Google Scholar] [CrossRef]
- Chavan, P.; Sharma, P.; Sharma, S.R.; Mittal, T.C.; Jaiswal, A.K. Application of high-intensity ultrasound to improve food processing efficiency: A review. Foods 2022, 11, 122. [Google Scholar] [CrossRef]
- Jukonyte, R.; Zadeike, D.; Bartkiene, E.; Lele, V.; Cernauskas, D.; Suproniene, S.; Juodeikiene, G. A potential of brown rice polish as a substrate for the lactic acid and bioactive compounds production by the lactic acid bacteria newly isolated from cereal-based fermented products. LWT 2018, 97, 323–331. [Google Scholar] [CrossRef]
- Cupp-Enyard, C. Sigma’s Non-specific Protease Activity Assay—Casein as a Substrate. J. Vis. Exp. 2008, 19, e899. [Google Scholar]
- Dhaka, V.; Khatkar, B. Effects of gliadin/glutenin and HMW-GS/LMW-GS ratio on dough rheological properties and bread-making potential of wheat varieties: Gluten proteins, dough rheology and bread quality. J. Food Qual. 2015, 38, 71–82. [Google Scholar] [CrossRef]
- Siddiqi, R.A.; Sogi, D.S.; Sehajpal, P.K. Effect of short-term sourdough fermentation on wheat protein. Cogent Food Agric. 2016, 2, 1132983. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2018. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Miah, M.A.S.; Islam, M.F.; Bhuiyan, M.N.I.; Tisa, K.J.; Naim, M.R. Exploring the effects of spontaneous and solid-state fermentation on the physicochemical, functional and structural properties of whole wheat flour (Triticum aestivum L.). Innov. Food Sci. Emerg. Technol. 2024, 97, 103798. [Google Scholar] [CrossRef]
- Fraberger, V.; Ladurner, M.; Nemec, A.; Grunwald-Gruber, C.; Call, L.M.; Hochegger, R.; Domig, K.J.; D’Amico, S. Insights into the potential of sourdough-related lactic acid bacteria to degrade proteins in wheat. Microorganisms 2020, 8, 1689. [Google Scholar] [CrossRef]
- Canesin, M.R.; Cazarin, C.B.B. Nutritional quality and nutrient bioaccessibility in sourdough bread. Curr. Opin. Food Sci. 2021, 40, 81–86. [Google Scholar] [CrossRef]
- Collar, C.; Mascaros, A.F.; Prieto, J.A.; De Barber, C.B. Changes in free amino acids during fermentation of wheat doughs started with pure culture of lactic acid bacteria. Cereal Chem. 1991, 68, 66–72. [Google Scholar]
- Tseng, H.-C.; Yang, C.-Y. Assessment of ultrasonic stress on survival and β-glucosidase activity of encapsulated Lactiplantibacillus plantarum BCRC 10357 in fermentation of black soymilk. Foods 2022, 11, 1234. [Google Scholar] [CrossRef]
- Pitt, W.G.; Ross, S.A. Ultrasound increases the rate of bacterial cell growth. Biotechnol. Prog. 2003, 19, 1038–1044. [Google Scholar] [CrossRef]
- Bolívar-Jacobo, N.A.; Reyes-Villagrana, R.A.; Espino-Solís, G.P.; Rentería-Monterrubio, A.L.; Arévalos-Sánchez, M.M.; Sánchez-Vega, R.; Santellano-Estrada, E.; Chávez-Flores, D.; Chávez-Martínez, A. The Effects of a High-Intensity Ultrasound on the Fermentative Activity and Kinetic Growth of Lactobacillus acidophilus and Lactobacillus helveticus. Fermentation 2023, 9, 356. [Google Scholar] [CrossRef]
- Shokri, S.; Shekarforoush, S.S.; Hosseinzadeh, S. Stimulatory effects of low-intensity ultrasound on the growth kinetics and metabolic activity of Lactococcus lactis subsp. lactis. Process Biochem. 2020, 89, 1–8. [Google Scholar] [CrossRef]
- Yu, Z.; Su, Y.; Zhang, Y.; Zhu, P.; Mei, Z.; Zhou, X.; Yu, H. Potential use of ultrasound to promote fermentation, maturation, and properties of fermented foods: A review. Food Chem. 2021, 357, 129805. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Chen, S.; Tang, Y.; Dai, C.; Sun, L.; Ma, H.; He, R. Stimulation of low-intensity ultrasound on fermentation of skim milk medium for yield of yoghurt peptides by Lactobacillus paracasei. Ultrason. Sonochem. 2019, 51, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.M.B.; Jafarpour, D.; Soto, E.R.; Barba, F.J. Ultrasound-assisted lactic acid fermentation of Bakraei (Citrus reticulata cv. Bakraei) juice: Physicochemical and bioactive properties. Fermentation 2023, 9, 37. [Google Scholar] [CrossRef]
- Bradauskiene, V.; Vaiciulyte-Funk, L.; Cernauskas, D.; Dzingeleviciene, R.; Lima, J.P.M.; Bradauskaite, A.; Tita, M.A. The efficacy of plant enzymes bromelain and papain as a tool for reducing gluten immunogenicity from wheat bran. Processes 2022, 10, 1948. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, H.; Wang, B.; Qu, W.; Li, Y.; He, R.; Wali, A. Effects of ultrasound pretreatment on the enzymolysis and structural characterization of wheat gluten. Food Biophys. 2015, 10, 385–395. [Google Scholar] [CrossRef]
- Marcuzzo, E.; Peressini, D.; Debeaufort, F.; Sensidoni, A. Effect of ultrasound treatment on properties of gluten-based film. Innov. Food Sci. Emerg. Technol. 2010, 11, 451–457. [Google Scholar] [CrossRef]
- Cheng, Y.; Donkor, P.O.; Yeboah, G.B.; Ayim, I.; Wu, J.; Ma, H. Modulating the in vitro digestion of heat-set whey protein emulsion gels via gelling properties modification with sequential ultrasound pretreatment. LWT 2021, 149, 111856. [Google Scholar] [CrossRef]
- Urade, R.; Sato, N.; Masaaki, S. Gliadins from wheat grain: An overview, from primary structure to nanostructures of aggregates. Biophys. Rev. 2018, 10, 435–443. [Google Scholar] [CrossRef]
- Zotta, T.; Piraino, P.; Ricciardi, A.; McSweeney, P.L.; Parente, E. Proteolysis in model sourdough fermentations. J. Agric. Food Chem. 2006, 54, 2567–2574. [Google Scholar] [CrossRef]
- Scherf, K.A.; Koehler, P.; Wieser, H. Gluten and wheat sensitivities. J. Cereal Sci. 2016, 67, 2–11. [Google Scholar] [CrossRef]
- Wieser, H.; Koehler, P.; Scherf, K.A. Chemistry of gluten proteins: Quantitative composition. Cereal Chem. 2023, 100, 36–55. [Google Scholar] [CrossRef]
- Nutter, J.; Saiz, A.I.; Iurlina, M.O. Microstructural and conformational changes of gluten proteins in wheat-rye sourdough. J. Cereal Sci. 2019, 87, 91–97. [Google Scholar] [CrossRef]
- Silva, N.H.C.S.; Vilela, C.; Marrucho, I.M.; Freire, C.S.R.; Neto, C.P.; Silvestre, A.J.D. Protein-based materials: From sources to innovative sustainable materials for biomedical applications. J. Mater. Chem. B 2014, 2, 3715–3740. [Google Scholar] [CrossRef]
- Loponen, J.; Mikola, M.; Katina, K.; Sontag-Strohm, T.; Salovaara, H. Degradation of HMW glutenins during wheat sourdough fermentations. Cereal Chem. 2004, 81, 87–93. [Google Scholar] [CrossRef]
- Dvoraček, V.; Čurn, V. Evaluation of protein fractions as biochemical markers for identification of spelt wheat cultivars (Triticum spelta L.). Plant Soil Environ. 2003, 49, 99–105. [Google Scholar] [CrossRef]
- Rizzello, C.G.; De Angelis, M.; Di Cagno, R.; Camarca, A.; Silano, M.; Losito, I.; De Vincenzi, M.; De Bari, M.D.; Palmisano, F.; Maurano, F.; et al. Highly efficient gluten degradation by lactobacilli and fungal proteases during food processing: New perspectives for celiac disease. Appl. Environ. Microbiol. 2007, 73, 4499–4507. [Google Scholar] [CrossRef]
- Pronin, D.; Geisslitz, S.; Börner, A.; Scherf, K.A. Fingerprinting of wheat protein profiles for improved distinction between wheat cultivars and species. Cereal Chem. 2020, 97, 999–1009. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y.; Tang, K.; Hu, Y.; Xu, X.; Gänzle, M.G. Effect of mixed cultures of yeast and lactobacilli on the quality of wheat sourdough bread. Front. Microbiol. 2019, 10, 2113. [Google Scholar] [CrossRef]
- Luo, D.; Wu, R.; Zhang, J.; Zhang, K.; Xu, B.; Li, P.; Yuan, Y.; Li, X. Effects of ultrasound-assisted dough fermentation on the quality of steamed bread. J. Cereal Sci. 2018, 83, 147–152. [Google Scholar] [CrossRef]
- Kłosok, K.; Welc, R.; Fornal, E.; Nawrocka, A. Effects of physical and chemical factors on the structure of gluten, gliadins and glutenins as studied with spectroscopic methods. Molecules 2021, 26, 508. [Google Scholar] [CrossRef] [PubMed]
- Dangi, P.; Chaudhary, N.; Khatkar, B.S. Rheological and microstructural characteristics of low molecular weight glutenin subunits of commercial wheats. Food Chem. 2019, 297, 124989. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, J.; Li, L.; Xiang, L.; Zhao, L.; Liu, J.; Liu, S.; Yang, Q.; Wu, J.; Chen, X. Effect of gliadin from Psathrostachys huashanica on dough rheological properties and biscuit quality. Food Chem. 2023, 425, 136537. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | pH | SP, mg/100 g | PA, PU/100 g | |||
|---|---|---|---|---|---|---|
| Pp | Lb | Pp | Lb | Pp | Lb | |
| WF0 h | 5.43 ± 0.01 a | 5.46 ± 0.02 a | 102.12 ± 1.43 e | 94.89 ± 1.37 e | 3.98 ± 0.51 e | 4.25 ± 0.57 d |
| WF24 h | 4.95 ± 0.02 b | 5.15 ± 0.01 b | 136.86 ± 1.51 d | 145.32 ± 1.41 c | 10.93 ± 0.61 d | 15.29 ± 0.61 b |
| WF48 h | 3.96 ± 0.01 c | 4.43 ± 0.02 c | 158.78 ± 2.13 b | 174.91 ± 2.53 a | 11.67 ± 0.47 c | 26.57 ± 0.56 a |
| WF72 h | 3.64 ± 0.01 d | 3.91 ± 0.01 d | 165.24 ± 1.39 a | 161.26 ± 1.89 b | 15.53 ± 0.64 a | 12.83 ± 0.57 c |
| WF96 h | 3.43 ± 0.02 e | 3.62 ± 0.01 e | 142.51 ± 1.52 c | 122.05 ± 1.42 d | 13.74 ± 0.85 b | 11.76 ± 0.64 c |
| Amino Acid | UWF | P. pentosaceus FST22 | L. brevis FST140 |
|---|---|---|---|
| Valine (Val) | 53 | 68 | 83 |
| Isoleucine (Ile) | 31 | 61 | 71 |
| Leucine (Leu) | 63 | 217 | 245 |
| Lysine (Lys) | 22 | 157 | 141 |
| Methionine (Met) | 37 | 50 | 151 |
| Phenylalanine (Phe) | 39 | 102 | 134 |
| Threonine (Thr) | 24 | 69 | 94 |
| Histidine (His) | 72 | 59 | 46 |
| Alanine (Ala) | 61 | 33 | 20 |
| Asparagine (Asp) | 69 | 50 | 227 |
| Serine (SER) | 48 | 158 | 115 |
| Glutamine (Glu) | 132 | 218 | 155 |
| Cystine (Cys) | 10 | 13 | 15 |
| Proline (Pro) | 38 | 249 | 253 |
| Glycine (Gly) | 32 | 98 | 106 |
| Tyrosine (Tyr) | 58 | 119 | 131 |
| Arginine (Arg) | 79 | 126 | 138 |
| Total FAAs | 868 c | 1847 b | 2125 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zadeike, D.; Cipkute, K.; Cizeikiene, D. Effects of Co-Fermentation with Lactic Acid Bacteria and Yeast on Gliadin Degradation in Whole-Wheat Sourdough. Fermentation 2025, 11, 238. https://doi.org/10.3390/fermentation11050238
Zadeike D, Cipkute K, Cizeikiene D. Effects of Co-Fermentation with Lactic Acid Bacteria and Yeast on Gliadin Degradation in Whole-Wheat Sourdough. Fermentation. 2025; 11(5):238. https://doi.org/10.3390/fermentation11050238
Chicago/Turabian StyleZadeike, Daiva, Kamile Cipkute, and Dalia Cizeikiene. 2025. "Effects of Co-Fermentation with Lactic Acid Bacteria and Yeast on Gliadin Degradation in Whole-Wheat Sourdough" Fermentation 11, no. 5: 238. https://doi.org/10.3390/fermentation11050238
APA StyleZadeike, D., Cipkute, K., & Cizeikiene, D. (2025). Effects of Co-Fermentation with Lactic Acid Bacteria and Yeast on Gliadin Degradation in Whole-Wheat Sourdough. Fermentation, 11(5), 238. https://doi.org/10.3390/fermentation11050238