
Lactic Acid Production by Enterococcus durans Is Improved by Cell Recycling and pH Control


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Media and Culture Conditions
2.2. Cell Growth Profile in Liquid Medium
2.3. Lactic Acid Production at Two Different Substrate Concentrations
2.4. Lactic Acid Production under Anaerobic Conditions
2.5. Lactic Acid Production with Cell Recycling
2.6. Batch with Pulse Lactic Acid Production with pH Control
2.7. Analytical Methods
2.8. Fermentation Yield
3. Results and Discussion
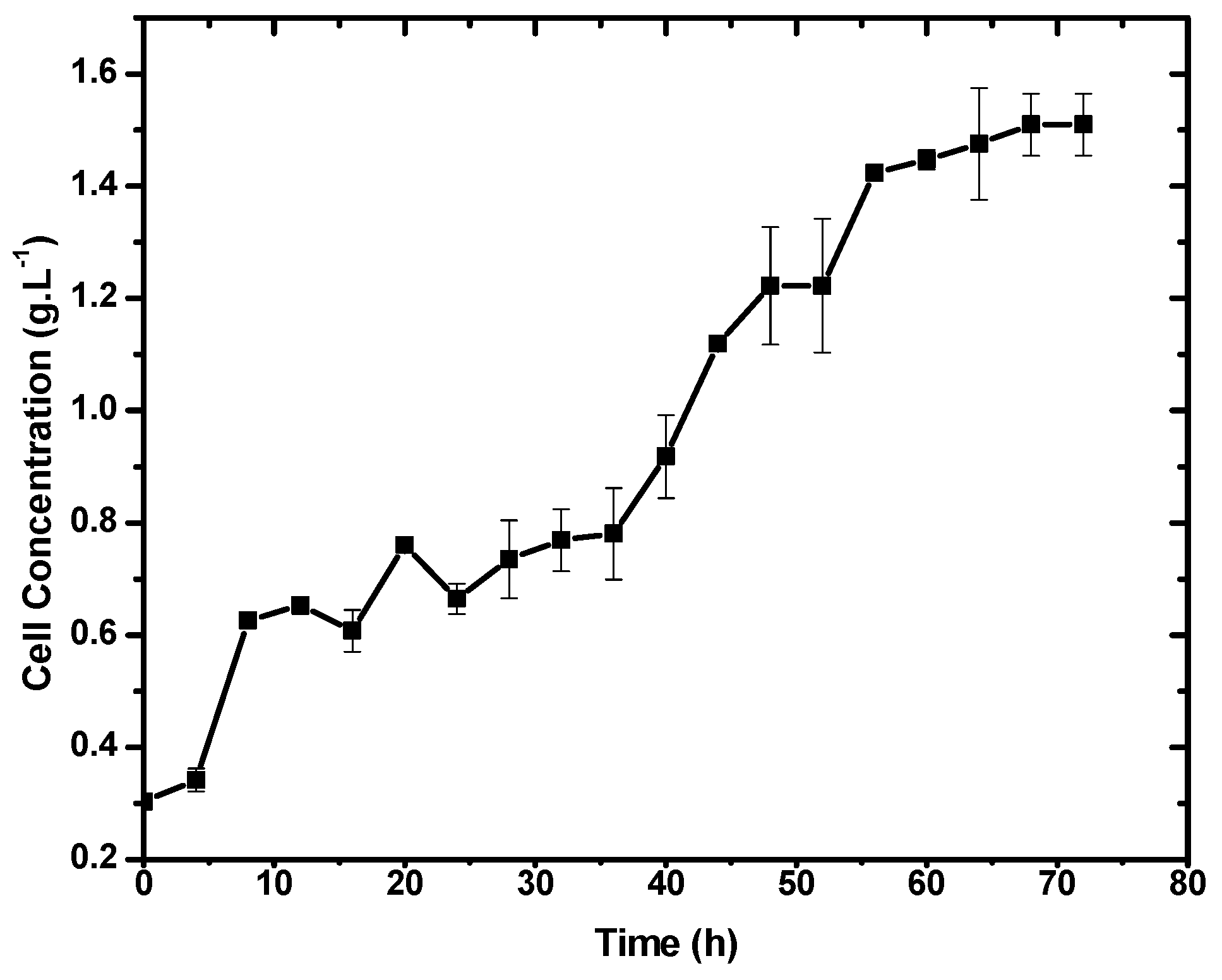
3.1. Cell Growth Profile in Liquid Medium
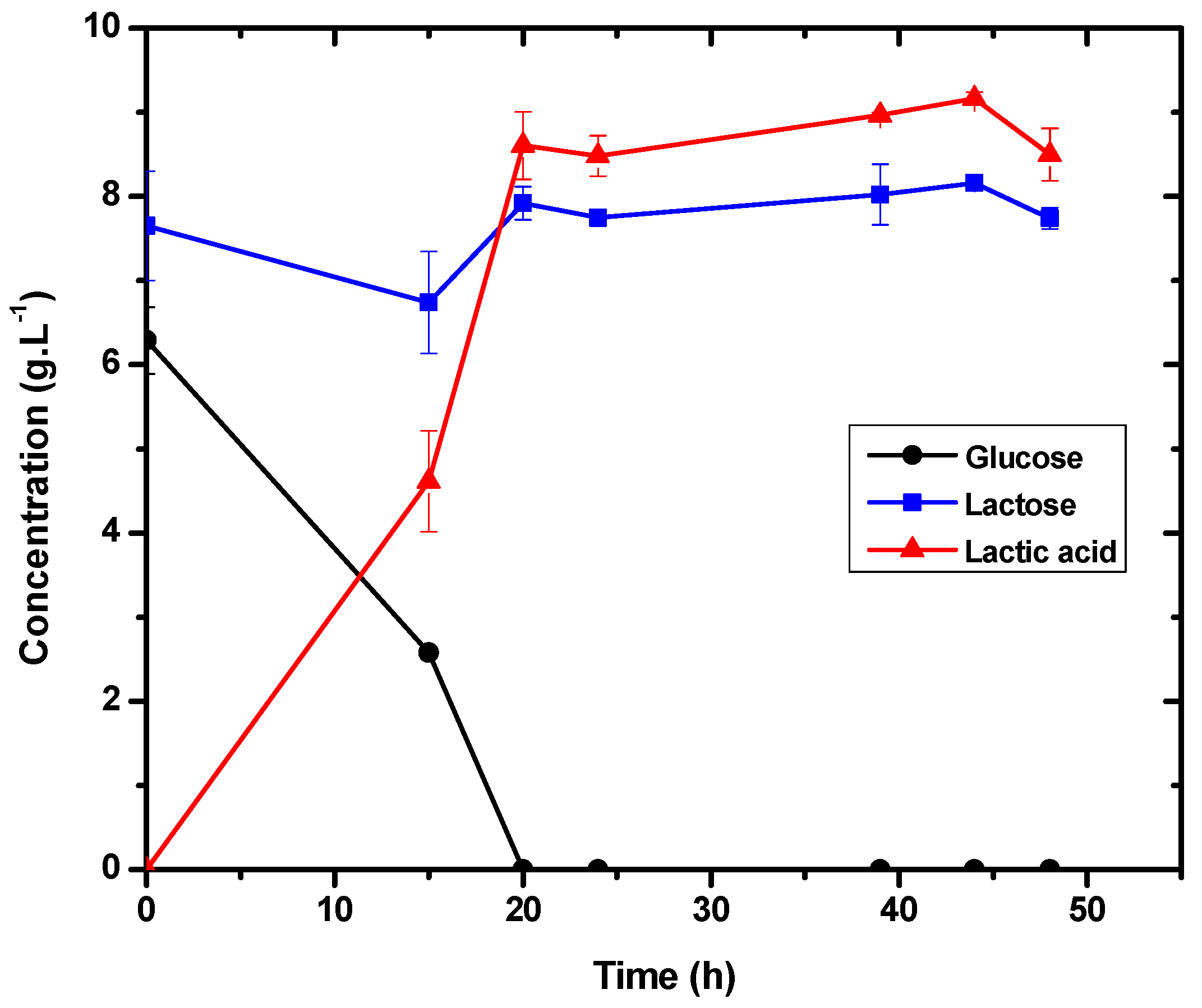
3.2. Evaluation of Lactic Acid Production at Two Different Substrate Concentrations
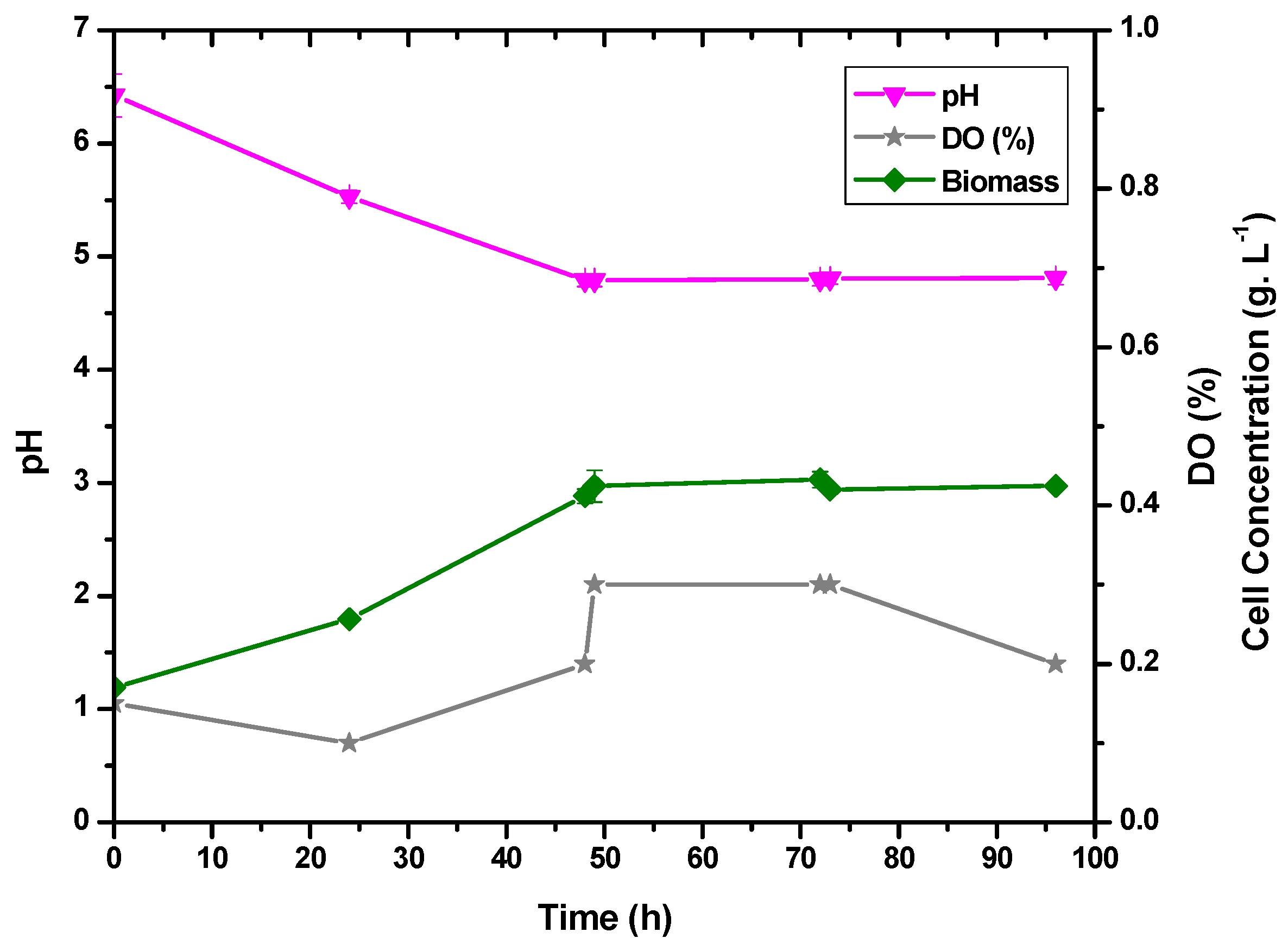
3.3. Evaluation of the Anaerobic Condition for Lactic Acid Production
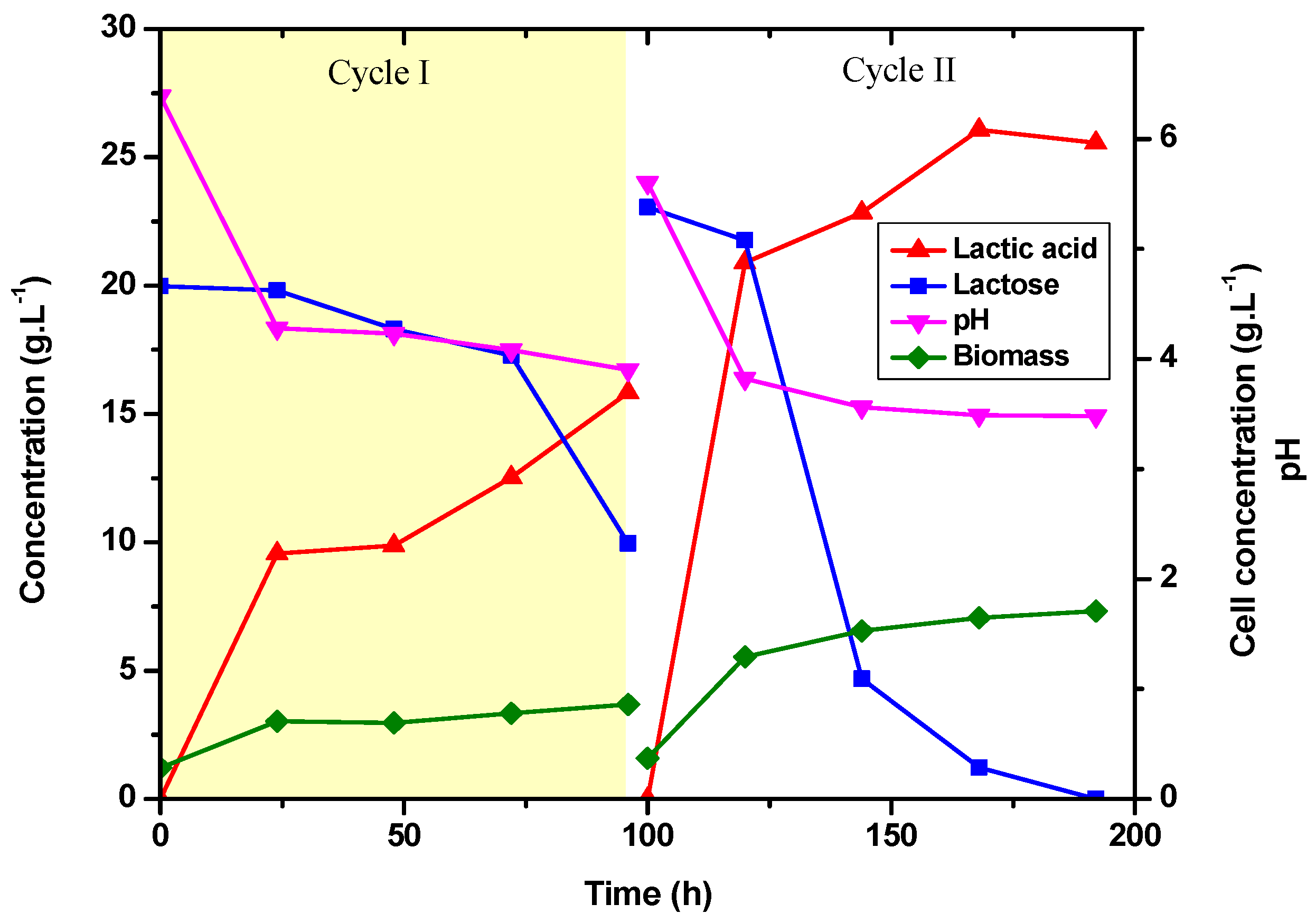
3.4. Evaluation of Cell Recycling for Lactic Acid Production with E. durans
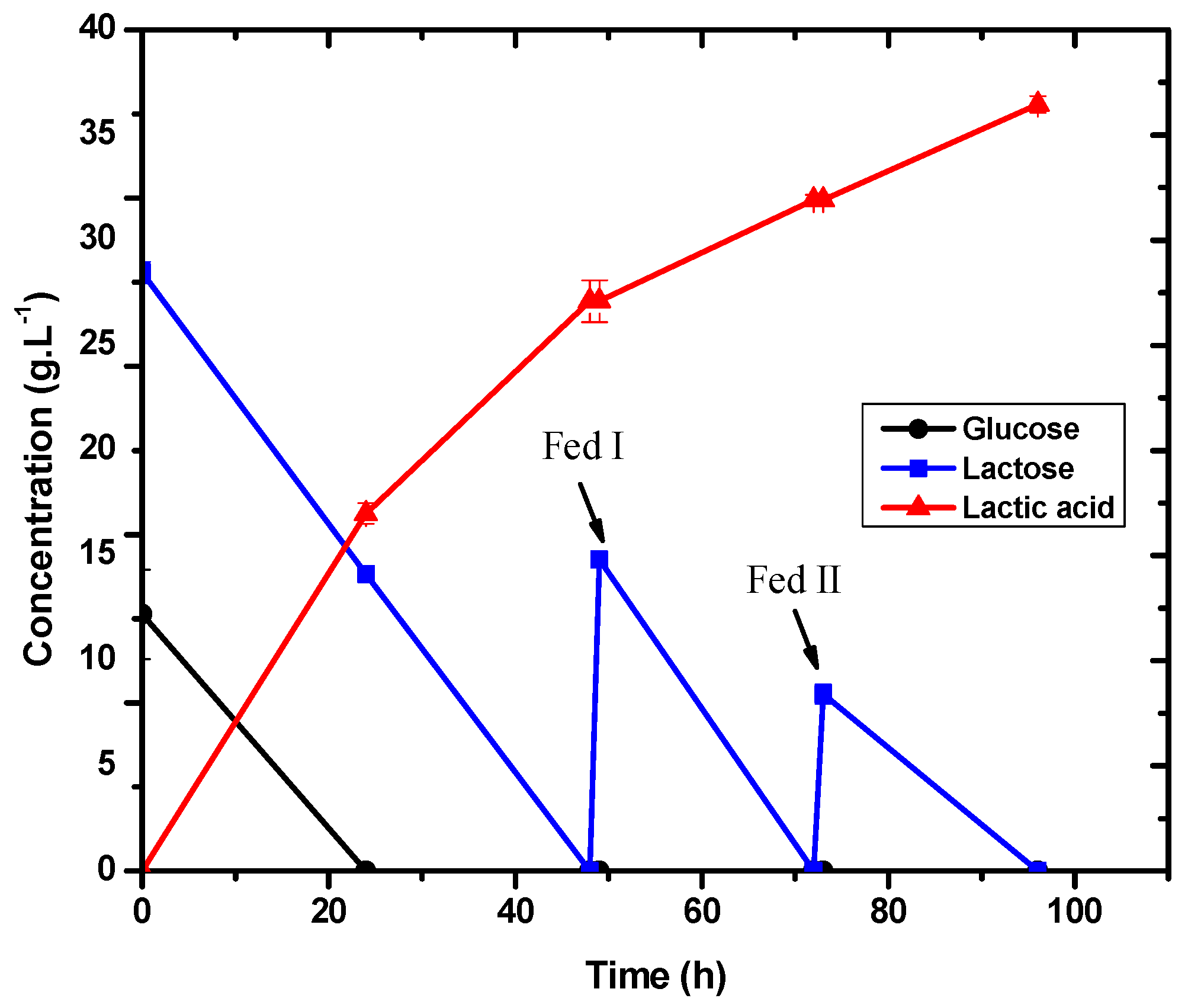
3.5. Evaluation of Batch with Pulse Lactic Acid Production with E. durans
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lactic Acid Market by Application (Biodegradable Polymers, Food & Beverages, Pharmaceutical Products), Form, and Region, Polylactic Acid Market, by Application (Packaging, Fiber & Fabrics, Agriculture), Form, and Region—Global Forecast to 2025. Available online: https://www.marketsandmarkets.com/Market-Reports/ (accessed on 22 February 2022).
- Komesu, A.; Allan Rocha de Oliveira, J.; Helena da Silva Martins, L.; Regina Wolf Maciel, M.; Maciel Filho, R. Lactic acid production to purification: A review. BioResources 2017, 12, 4364–4383. [Google Scholar] [CrossRef]
- Barroso, R.G.M.R.; Gonçalves, S.B.; Machado, F. A Novel Approach for the Synthesis of Lactic Acid-Based Polymers in an Aqueous Dispersed Medium. Sustain. Chem. Pharm. 2020, 15, 100211. [Google Scholar] [CrossRef]
- Esmaeili, N.; Jahandideh, A.; Muthukumarappan, K.; Åkesson, D.; Skrifvars, M. Synthesis and Characterization of Methacrylated Star-Shaped Poly(Lactic Acid) Employing Core Molecules with Different Hydroxyl Groups. J. Appl. Polym. Sci. 2017, 134, 45341. [Google Scholar] [CrossRef]
- Gupta, B.; Revagade, N.; Hilborn, J. Poly(Lactic Acid) Fiber: An Overview. Prog. Polym. Sci. 2007, 32, 455–482. [Google Scholar] [CrossRef]
- Pang, Y.; Ardagh, M.A.; Shetty, M.; Chatzidimitriou, A.; Kumar, G.; Vlaisavljevich, B.; Dauenhauer, P.J. On the Spatial Design of Co-Fed Amines for Selective Dehydration of Methyl Lactate to Acrylates. ACS Catal. 2021, 11, 5718–5735. [Google Scholar] [CrossRef]
- Pang, Y.; Lee, C.; Vlaisavljevich, B.; Nicholas, C.P.; Dauenhauer, P.J. Multifunctional Amine Modifiers for Selective Dehydration of Methyl Lactate to Acrylates. JACS Au 2023, 3, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Makshina, E.V.; Canadell, J.; van Krieken, J.; Sels, B.F. Potassium-Modified ZSM-5 Catalysts for Methyl Acrylate Formation from Methyl Lactate: The Impact of the Intrinsic Properties on Their Stability and Selectivity. ACS Sustain. Chem. Eng. 2021, 10, 6196–6204. [Google Scholar] [CrossRef]
- Hofvendahl, K.; Hahn-Hägerdal, B. Factors Affecting the Fermentative Lactic Acid Production from Renewable Resources. Enzym. Microb. Technol. 2000, 26, 87–107. [Google Scholar] [CrossRef] [PubMed]
- Barroso, R.G.M.R.; Lima, J.R.C.; Fávaro, L.C.L.; Machado, F.; Gonçalves, S.B. Bioconversion of Glycerol into Lactic Acid by a New Bacterial Strain from the Brazilian Cerrado Soil. Fermentation 2022, 8, 477. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent Advances in Lactic Acid Production by Microbial Fermentation Processes. Biotechnol. Adv. 2013, 31, 877–902. [Google Scholar] [CrossRef]
- Abedi, E.; Hashemi, S.M.B. Lactic Acid Production—Producing Microorganisms and Substrates Sources-State of Art. Heliyon 2020, 6, e04974. [Google Scholar] [CrossRef] [PubMed]
- Giraffa, G. Enterococci from Foods. FEMS Microbiol. Rev. 2002, 26, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.C.; Esteves, C.T.; Palazzo, I.C.V.; Darini, A.L.C.; Felis, G.E.; Sechi, L.A.; Franco, B.D.G.M.; de Martinis, E.C.P. Prevalence and Characterization of Enterococcus Spp. Isolated from Brazilian Foods. Food Microbiol. 2008, 25, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Cock, L.S.; De Stouvenel, A.R. Lactic Acid Production by a Strain of Lactococcus Lactis Subs Lactis Isolated from Sugar Cane Plants. Electron. J. Biotechnol. 2006, 9, 40–45. [Google Scholar] [CrossRef]
- Panesar, P.S.; Kennedy, J.F.; Knill, C.J.; Kosseva, M. Production of L(+) Lactic Acid Using Lactobacillus Casei from Whey. Braz. Arch. Biol. Technol. 2010, 53, 219–226. [Google Scholar] [CrossRef]
- Pessione, A.; Zapponi, M.; Mandili, G.; Fattori, P.; Mangiapane, E.; Mazzoli, R.; Pessione, E. Enantioselective Lactic Acid Production by an Enterococcus Faecium Strain Showing Potential in Agro-Industrial Waste Bioconversion: Physiological and Proteomic Studies. J. Biotechnol. 2014, 173, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Verma, D.; Subudhi, S. ‘Lactobacillus sp. Strain TERI-D3’, as Microbial Cell Factory for Fermentative Production of Lactic Acid’. Curr. Res. Green Sustain. Chem. 2021, 4, 100059. [Google Scholar] [CrossRef]
- Nagarajan, D.; Oktarina, N.; Chen, P.-T.; Chen, C.-Y.; Lee, D.-J.; Chang, J.-S. Fermentative Lactic Acid Production from Seaweed Hydrolysate Using Lactobacillus sp. and Weissella sp. Bioresour. Technol. 2022, 344, 126166. [Google Scholar] [CrossRef]
- Murakami, N.; Oba, M.; Iwamoto, M.; Tashiro, Y.; Noguchi, T.; Bonkohara, K.; Abdel-Rahman, M.A.; Zendo, T.; Shimoda, M.; Sakai, K.; et al. L-Lactic Acid Production from Glycerol Coupled with Acetic Acid Metabolism by Enterococcus Faecalis without Carbon Loss. J. Biosci. Bioeng. 2016, 121, 89–95. [Google Scholar] [CrossRef]
- Yuan, S.F.; Hsu, T.C.; Wang, C.A.; Jang, M.F.; Kuo, Y.C.; Alper, H.S.; Guo, G.L.; Hwang, W.S. Production of Optically Pure l(+)-Lactic Acid from Waste Plywood Chips Using an Isolated Thermotolerant Enterococcus Faecalis SI at a Pilot Scale. J. Ind. Microbiol. Biotechnol. 2018, 45, 961–970. [Google Scholar] [CrossRef]
- Hassan, S.E.D.; Abdel-Rahman, M.A.; Roushdy, M.M.; Azab, M.S.; Gaber, M.A. Effective Biorefinery Approach for Lactic Acid Production Based on Co-Fermentation of Mixed Organic Wastes by Enterococcus Durans BP130. Biocatal. Agric. Biotechnol. 2019, 20, 101203. [Google Scholar] [CrossRef]
- Hayat, Z.; Mukhtar, I.; Shair, F.; Ullah, K.; Hafeez, A.; Hafeez, F.Y.; Ullah, A. Production of Antifungal Gelatinase by Enterococcus Durans S2C and Its Potential Role in the Biological Control. Int. J. Agric. Biol. 2021, 25, 1043–1050. [Google Scholar] [CrossRef]
- Estilarte, M.L.; Tymczyszyn, E.E.; de los Ángeles Serradell, M.; Carasi, P. Freeze-Drying of Enterococcus Durans: Effect on Their Probiotics and Biopreservative Properties. LWT 2021, 137, 110496. [Google Scholar] [CrossRef]
- Hussein, W.E.; Abdelhamid, A.G.; Rocha-Mendoza, D.; García-Cano, I.; Yousef, A.E. Assessment of Safety and Probiotic Traits of Enterococcus Durans OSY-EGY, Isolated from Egyptian Artisanal Cheese, Using Comparative Genomics and Phenotypic Analyses. Front. Microbiol. 2020, 11, 608314. [Google Scholar] [CrossRef] [PubMed]
- Yerlikaya, O.; Akbulut, N. In Vitro Characterisation of Probiotic Properties of Enterococcus Faecium and Enterococcus Durans Strains Isolated from Raw Milk and Traditional Dairy Products. Int. J. Dairy Echnol. 2020, 73, 98–107. [Google Scholar] [CrossRef]
- Simonová, M.P.; Lauková, A.; Chrastinová, L.; Kandričáková, A.; Ščerbová, J.; Formelová, Z.; Chrenková, M.; Zitňan, R.; Miltko, R.; Bełzecki, G. Effect of Diet Supplementation with Enterococcus Durans ED26E/7 and Its Durancin ED26E/7 on Growth Performance, Caecal Enzymatic Activity, Jejunal Morphology and Meat Properties of Broiler Rabbits. Ann. Anim. Sci. 2022, 22, 221–235. [Google Scholar] [CrossRef]
- Lauková, A.; Tomáška, M.; Kmet’, V.; Strompfová, V.; Simonová, M.P.; Dvorožňáková, E. Slovak Local Ewe’s Milk Lump Cheese, a Source of Beneficial Enterococcus Durans Strain. Foods 2021, 10, 3091. [Google Scholar] [CrossRef]
- Bosco, F.; Cirrincione, S.; Carletto, R.; Marmo, L.; Chiesa, F.; Mazzoli, R.; Pessione, E. Pha Production from Cheese Whey and “Scotta”: Comparison between a Consortium and a Pure Culture of Leuconostoc mesenteroides. Microorganisms 2021, 9, 2426. [Google Scholar] [CrossRef]
- García-Depraect, O.; León-Becerril, E. Use of a Highly Specialized Biocatalyst to Produce Lactate or Biohydrogen and Butyrate from Agro-Industrial Resources in a Dual-Phase Dark Fermentation. Fermentation 2023, 9, 787. [Google Scholar] [CrossRef]
- Dareioti, M.A.; Vavouraki, A.I.; Tsigkou, K.; Zafiri, C.; Kornaros, M. Dark Fermentation of Sweet Sorghum Stalks, Cheese Whey and Cow Manure Mixture: Effect of PH, Pretreatment and Organic Load. Processes 2021, 9, 1017. [Google Scholar] [CrossRef]
- Bustamante, D.; Tortajada, M.; Ramón, D.; Rojas, A. Production of D-lactic acid by the fermentation of orange peel waste hydrolysate by lactic acid bacteria. Fermentation 2020, 6, 1. [Google Scholar] [CrossRef]
- Boonmee, M.; Leksawasdi, N.; Bridge, W.; Rogers, P.L. Batch and Continuous Culture of Lactococcus Lactis NZ133: Experimental Data and Model Development. Biochem. Eng. J. 2003, 14, 127–135. [Google Scholar] [CrossRef]
- Jangra, M.; Belur, P.D.; Oriabinska, L.B.; Dugan, O.M. Multistrain Probiotic Production by Co-Culture Fermentation in a Lab-Scale Bioreactor. Eng. Life Sci. 2016, 16, 247–253. [Google Scholar] [CrossRef]
- Zotta, T.; Ricciardi, A.; Ianniello, R.G.; Parente, E.; Reale, A.; Rossi, F.; Iacumin, L.; Comi, G.; Coppola, R. Assessment of Aerobic and Respiratory Growth in the Lactobacillus Casei Group. PLoS ONE 2014, 9, e99189. [Google Scholar] [CrossRef]
- Zotta, T.; Parente, E.; Ricciardi, A. Aerobic Metabolism in the Genus Lactobacillus: Impact on Stress Response and Potential Applications in the Food Industry. J. Appl. Microbiol. 2017, 122, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.Y.; Xu, Y.; Chen, X.; Zhang, L.Y. Regulation of Metabolic Flux in Lactobacillus Casei for Lactic Acid Production by Overexpressed LdhL Gene with Two-Stage Oxygen Supply Strategy. J. Microbiol. Biotechnol. 2015, 25, 81–88. [Google Scholar] [CrossRef]
- Joulak, I.; Concórdio-Reis, P.; Torres, C.A.V.; Sevrin, C.; Grandfils, C.; Attia, H.; Freitas, F.; Reis, M.A.M.; Azabou, S. Sustainable use of agro-industrial wastes as potential feedstocks for exopolysaccharide production by selected Halomonas strains. Environ. Sci. Pollut. Res. 2022, 29, 22043–22055. [Google Scholar] [CrossRef] [PubMed]
- Alfano, A.; D’ambrosio, S.; D’agostino, A.; Finamore, R.; Schiraldi, C.; Cimini, D. Concentrated Buffalo Whey as Substrate for Probiotic Cultures and as Source of Bioactive Ingredients: A Local Circular Economy Approach towards Reuse of Wastewaters. Fermentation 2021, 7, 281. [Google Scholar] [CrossRef]
- Aso, Y.; Tsubaki, M.; Dang Long, B.H.; Murakami, R.; Nagata, K.; Okano, H.; Phuong Dung, N.T.; Ohara, H. Continuous Production of D-Lactic Acid from Cellobiose in Cell Recycle Fermentation Using β-Glucosidase-Displaying Escherichia coli. J. Biosci. Bioeng. 2019, 127, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zheng, Z.; Xu, Q.; Qian, Z.; Liu, J.; Ouyang, J. Valorization of Dairy Waste for Enhanced D-Lactic Acid Production at Low Cost. Process. Biochem. 2018, 71, 18–22. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Xiao, Y.; Tashiro, Y.; Wang, Y.; Zendo, T.; Sakai, K.; Sonomoto, K. Fed-Batch Fermentation for Enhanced Lactic Acid Production from Glucose/Xylose Mixture without Carbon Catabolite Repression. J. Biosci. Bioeng. 2015, 119, 153–158. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Source | Fermentation Strategies | Lactic Acid (g·L−1) | Yield (g·g−1) | Reference |
|---|---|---|---|---|---|
| Lactococcus lactis subs lactis | Glucose | Batch | 35.0 | 0.45 | Cock [15] |
| Lactobacillus casei | Lactose | Batch | 33.73 | 0.88 | Panesar [16] |
| Enterococcus faecium | Glucose | Batch | 18 | 0.9 | Pessione [17] |
| Enterococcus faecium | Fructose | Batch | 18 | 0.9 | Pessione [17] |
| Enterococcus faecium | Cellobiose | Batch | 13.5 | 0.71 | Pessione [17] |
| Enterococcus faecium | Xylose | Batch | 4.5 | 0.27 | Pessione [17] |
| Lactobacillus sp. TERI-D3 | Sucrose | Batch | 19.4 | 0.86 | Verma [18] |
| Lactobacillus sp. TERI-D3 | Galactose | Batch | 15.8 | 0.8 | Verma [18] |
| Lactobacillus sp. TERI-D3 | Lactose | Batch | 18.1 | 0.9 | Verma [18] |
| Lactobacillus sp. TERI-D3 | Glucose | Batch | 17.5 | 0.9 | Verma [18] |
| Weissella paramesenteroides | Glucose | Batch | 20 | 0.68 | Nagarajan [19] |
| Enterococcus faecalis | Glycerol | Fed-batch | 55.3 | 0.99 | Murakami [20] |
| Enterococcus faecalis | Waste plywood chips | Batch | 92 | 0.92 | Yuan [21] |
| Enterococcus durans BP130 | Food waste | Batch | 28.8 | 0.85 | Hassan [22] |
| Operation Mode | Lactic Acid (g·L−1) | Yield (g·g−1) | Productivity (g·L−1·h−1) |
|---|---|---|---|
| Simple Batch | 10.87 | 0.94 | 0.54 |
| Anaerobiosis | 9.17 | 0.69 | 0.21 |
| Cell recycling | 26.07 | 0.82 | 0.36 |
| Batch with pulse and pH control | 36.47 | 0.57 | 0.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barroso, R.G.M.R.; Damaso, M.C.T.; Machado, F.; Gonçalves, S.B. Lactic Acid Production by Enterococcus durans Is Improved by Cell Recycling and pH Control. Fermentation 2024, 10, 149. https://doi.org/10.3390/fermentation10030149
Barroso RGMR, Damaso MCT, Machado F, Gonçalves SB. Lactic Acid Production by Enterococcus durans Is Improved by Cell Recycling and pH Control. Fermentation. 2024; 10(3):149. https://doi.org/10.3390/fermentation10030149
Chicago/Turabian StyleBarroso, Raissa Gabriela Martins Reis, Mônica Caramez Triches Damaso, Fabricio Machado, and Sílvia Belém Gonçalves. 2024. "Lactic Acid Production by Enterococcus durans Is Improved by Cell Recycling and pH Control" Fermentation 10, no. 3: 149. https://doi.org/10.3390/fermentation10030149
APA StyleBarroso, R. G. M. R., Damaso, M. C. T., Machado, F., & Gonçalves, S. B. (2024). Lactic Acid Production by Enterococcus durans Is Improved by Cell Recycling and pH Control. Fermentation, 10(3), 149. https://doi.org/10.3390/fermentation10030149