Abstract
Biodiesel produced from waste cooking oil (WCO) is on the rise and inevitably leads to issues in managing glycerol waste. Due to the presence of colour, odour and other minor compounds, the refining costs for this type of glycerol are higher and uneconomical. The potential of biodiesel-derived glycerol waste (BDGW) obtained from WCO to produce the highly added product of docosahexaenoic acid (DHA), also known as omega-3 polyunsaturated fatty acid, via the marine microalga of Schizochytrium limacinum ATCC MYA-1381 under aerobic batch fermentation was investigated. Cell growth, as well as DHA production, were performed under various operating conditions, including aeration rates and BDGW concentrations. The effect of the substrate type on cell growth and DHA yield was evaluated. The optimum operating condition was obtained when the air flow of a 0.25 vvm and 50 g/L of the glycerol concentration was fed into the fermenter and maximum cell dry weight (11.40 g/L) and DHA yield (665.52 mg/g) were achieved. However, cell growth and DHA yield were not significantly different when S. limacinum was grown using various carbon sources. Successfully, it clearly demonstrates that the BDGW can be used as a cheap carbon source for DHA production via marine microalgae using aerobic batch fermentation.
1. Introduction
The ever-increasing demand and production worldwide over the past few years of biodiesel, an environmentally viable fuel, has inevitability led to a sharp increase in its associated co-product, glycerol, which represents 10% of the weight of the esters. This increase in glycerol production from biodiesel refining has created a glut in the glycerol market and has led to the price of glycerol falling significantly [1]. Typically, the crude glycerol obtained after biodiesel production contains many impurities such as soap and methanol, and thus, it is prohibitively expensive and has substantial commercial value if converted and purified into a raw material that can be used in food, cosmetics, or pharmaceutical industries. With the rapid growth in biodiesel production, the market is flooded with crude glycerol; as a result, biodiesel producers prefer to find out new uses for this waste stream. However, the cost to refine this product is too costly and energy-intensive. Extensive studies have been undertaken to use crude glycerol as a raw material to convert glycerol into products through thermochemical and biological methods, such as propylene glycol, acetol, 1,3-propanediol, lipid, and pigment. Furthermore, crude glycerol is also used as feedstock for producing hydrogen gas through photo-fermentation or gasification processes [2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18].
Naturally, microalgae can be grown in photoautotrophic conditions and can be classified into three growth patterns named autotrophic cultivation (AC), heterotrophic cultivation (HC), and mixotrophic cultivation (MC), respectively. The AC pattern occurs when they use light as the energy source and carbon dioxide (CO2) as the inorganic carbon source. However, the limitation of the AC is the low maximum biomass concentration attainable as the light available for growth with serious contamination by other microorganisms. In some cases, the pattern of HC means microalgal strains can use various organic carbons to grow in dark conditions. In addition, the MC is an alternative choice to AC, which combines the use of organic carbon and light as energy sources. For the MC, it has the potential to attain a high biomass concentration while maintaining the high content of the valuable products in the microalgal biomass that are usually lost under HC growth.
Typically, polyunsaturated fatty acids (PUFAs) are important long-chain omega-3 polyunsaturated fatty acids that have been shown to have beneficial influences on preventing human cognitive function, brain development function, cardiovascular diseases, cancer, schizophrenia, and Alzheimer’s disease [19,20,21,22,23,24,25]. Among the various PUFAs, the most important categories of PUFAs are omega-3 fatty acids (n-3 PUFAs) and omega-6 fatty acids (n-6 PUFAs). The form of n-3-PUFAs relevant for human health are α-linolenic acid (ALA, C18: 3n-3), eicosapentaenoic acid (EPA, C20: 5n-3) and docosahexaenoic acid (DHA, C22: 6n-3). Meanwhile, for the n-6 cateffamily, there are linoleic acid (LA, C18: 2n-6), di homo-γ-linolenic acid (DGLA, 20: 3n-6), and arachidonic acid (AA, C20: 4n-6) [26]. However, DHA has received much more attention from many researchers. In recent decades, oils derived from microorganisms such as microalgae, bacteria, yeast, fungi, and bacteria have been shown to possess oil-producing capabilities, which are often referred to as microbial oils, and have emerged as alternatives for useful PUFA productions. Compared to various traditional vegetable and animal sources, the production of microbial oils offers many interesting advantages, such as a shorter life cycle; less labour required; being affected less by season and climate change; and being easier to scale up. Therefore, DHA may become one of the value-added products for essential supplement production produced by microbial sources in the future [17,18,19,20,21,22,23,24,25,26,27,28].
Previously, several researchers reported on the production of DHA using marine microalgae such as Schizochytrium limacinum, Thraustochytrium sp., etc., via a fermentation process using crude glycerol as a substrate [2,17]. In 2007, Chi et al. and later Pyle et al., 2008 [2,17] used crude glycerol derived from soybean oil. During recent years, biodiesel feedstocks such as WCO have risen owing to the challenges that soybean oil faces: unstable prices, competing with food, etc. The WCO is basically cheap by-products from commercial and industrial processes. Compared to vegetable oils, the cost of WCO is anywhere from 60% less to free, depending on the source and availability. However, crude glycerol produced from WCO has lower sale values due to the presence of colour odour contaminants as well as other minor compounds. The refining costs for this type of glycerin are higher [24]. However, crude glycerol-derived algae have no heavy metal contaminations. Thus, in order to develop a mass glycerol-based algal culture to supply a large amount of biomass, an in-depth study of the cell growth, substrate utilisation, and total lipid (including DHA) production profile of this algal species is needed. The DHA-rich algal biomass can also be used as a functional food during the culture and production of marine fish. Presently, fish oil is widely used as a traditional source of omega-3 fatty acids, although it is facing challenges such as odour and taste problems, heavy metal contamination, and limited supply.
The utilization of the BDGW as a carbon source under the HC microalgal cultivation has been considered. Therefore, this research work aims to (1) evaluate the growth of S. limacinum on crude glycerol derived from the WCO during biodiesel production and (2) obtain the optimal condition of aeration rate and glycerol concentration for maximal cell growth and DHA accumulation in its cell under aerobic HC in a batch fermenter.
2. Materials and Methods
2.1. Algal Strain and Cultivation Medium
The pure microalgal strain Schizochytrium limacinum ATCC MYA-1381 was kindly obtained from the Laboratory of Chemical Engineering and Advanced Materials, Newcastle University, Newcastle upon Tyne, UK. The algal strain was maintained on nutrient agar slants with glucose–peptone–yeast extract (GPY)/agar medium, which contained various chemical substances in v such as 2% glucose, 0.4% of a yeast extract, 1% peptone, and 2% agar and was sub-cultured on a monthly basis. The basal medium was modified from Wu et al. (2005) [29] and consisted of 25 g/L NaCl; 1 g/L KH2PO4; 5 g/L MgSO4·7H2O; 0.2 g/L CaCl2; and 1.0 g/L KCL. The trace mineral solution contained 6.0 g/L of Na2EDTA; 0.29 g/L of FeCl3·6H2O; 6.84 g/L of H3BO3; 0.86 g/L of MnCl2·4H2O; 0.06 g/L of ZnCl2; 0.026 g/L of CoCl2·6H2O, and 0.002 g/L of CuSO4·5 H2O. It should be noted that the algal strain used naturally lives in sea or saline water. The alga was cultured in the medium, and its growth pattern was monitored.
2.2. Pretreatment of Crude Glycerol
The BDGW used throughout the experiment was obtained from the biodiesel production pilot plant, Khon Kaen University, Khon Kaen, Thailand, as a byproduct of the transesterification of methanol and the WCO, together with potassium hydroxide as the catalyst. The WCO was collected from households and restaurants within Khon Kaen City, Thailand, and contained various crop oils (such as palm oil, sunflower oil, and soybean oil) and animal fats. It comprised approximately 41.0% of saturated fatty acids and 59.0% of unsaturated fatty acids. Palmitic acid was a major saturated fatty acid, whilst cis-9-oleic acid was a major unsaturated fatty acid. The BDGW mainly contained glycerol, soap, methanol, and water at 52.6%, 28.7%, and 18.8% by weight, respectively. The BDGW was pretreated to avoid soap precipitation when it was mixed with artificial seawater. The pretreatment protocol was modified from Chi et al. (2007) as follows; the BDGW was mixed with distilled water at a ratio of 1:4 by volume to reduce the viscosity of the fluid. Then, the pH of the fluid was adjusted from pH 9.8 to pH 6.5 with 1 N of hydrochloric (HCL) acid to convert the soluble soap into insoluble free fatty acids, which precipitated from the liquid. After that, the precipitated solid was separated from the BDGW solution by centrifugation at 8000 rpm; finally, after separation, other nutrients and chemicals (mineral salts, nitrogen source, including glycerol, etc.) and additional water were added to reach the desired levels. For example, in this study, the BDGW concentrations varied at 25, 50, 75, 100, and 125 m/L, respectively.
2.3. Aerobic Batch Heterotrophic Fermentation
The microalgal cultures were grown in 250 mL Erlenmeyer flasks containing 50 mL of the basal medium with 2% (w/v) glucose and 0.4% (w/v) of the yeast extract for 48 h, which the algal cell could reach an exponential growth phase. Cultures were inoculated at a rate of 5 % (v/v) and were incubated on an orbital shaker at 150 rpm at 25 °C with pH 6.5 for 8 days in the dark condition or the HC condition. The initial pH of the culture was controlled with a buffer (potassium dihydrogen phosphate–sodium hydroxide). In later cell growth and DHA production studies, glucose was replaced as the carbon source by either laboratory-grade glycerol or pretreated glycerol at 50 g/L, unless stated otherwise, obtained from the biodiesel industry. Normally, the flask cultures were loaded with 30% of the medium (unless stated otherwise), which corresponded to an aeration flow rate of 0.25 vvm. All the other components were the same as those used in the subculture. The culture was daily withdrawn to determine the growth curve, cell dry weight (CDW), and DHA content. The results obtained in this study were average values based on the simultaneous cultivation of three replications for reproducibility.
In addition, a large-scale HC batch cultivation in the 5 L fermenter (Biostat B 8840334, B Braun, Germany) was further performed using the optimal condition obtained from the flask scale.
2.4. Cell Growth and Lipid Extraction
The biomass was determined using the CDW. The culture was harvested and washed with distilled water twice via centrifugation and then dried at 70 °C in a vacuum oven until it obtained a constant weight. Fatty acid methyl esters (FAMEs) were prepared from dried algal biomass according to a modified protocol developed by Rattanapoltee et al. (2021) [30]. This method involves a 4 mL mixture; methanol, concentrated sulfuric acid, and chloroform (1.7:0.3:2.0, v/v/v) was added into a tube containing dried cell biomass and 1 mg pentadecanoic acid with 15:0 as an internal standard. The 20 mL sample was heated in a water bath at 90 °C for 40 min and later cooled down to room temperature, at which point 1 mL of distilled water was added. The liquid in the tubes was thoroughly mixed through a vortex for 1 min and then settled for the separation of the two phases. The lower phase containing the FAMEs was transferred to a clean vial and dried with anhydrous Na2SO4. Subsequently, FAMEs were analysed using gas chromatography (GC-17A, Shimadzu, Japan). The GC was equipped with an FID (Flame Ionisation Detector) and Fame Wax Capillary Column (30 m × 0.32 mm × 0.25 µm). Nitrogen was used as the carrier gas. The initial column temperature program was raised from 195 °C to 240 °C in increments of 2.5 °C min−1. FAMEs were identified by comparing the retention times with those of standard fatty acids (Sigma Chemical Co., Ltd., Waltham, MA, USA). The quantity of DHA was estimated from the peak areas on the chromatogram using pentadecanoic acid (15:0) as an internal standard.
3. Results and Discussion
3.1. Growth Monitoring and Morphology
The growth pattern of pure algal strain S. limacinum ATCC MYA-1381, in terms of biomass as a function of time, was monitored, as shown in Figure 1. It was found that the cell culture came up and reached the log or exponential growth phase at about 10 h and reached the maximum biomass at about 8.5 g/L after 72 h cultivation.
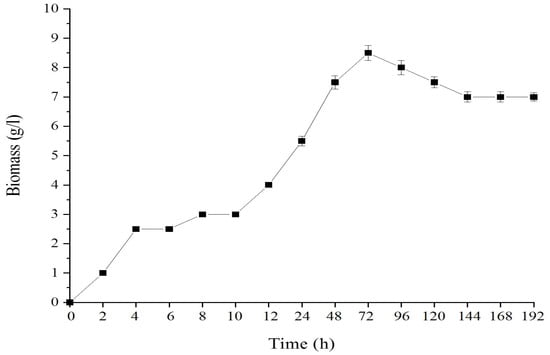
Figure 1.
Growth pattern of the pure algal strain of S. limacinum ATCC MYA-1381.
Meanwhile, Figure 2 shows the morphology of the algal strain (×40 magnification). It was found that the cell culture was increased from a single colony (lag phase) into clustered colonies (log phase).
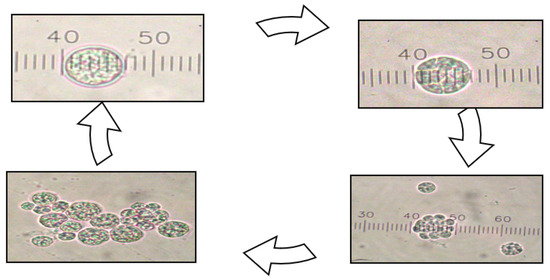
Figure 2.
Morphology of the pure algal strain of S. limacinum ATCC MYA-1381 in culture medium.
3.2. Characteristics of the BDWG
The BDWG before and after treatment was characterized. It was revealed that the pH was reduced from the basic value (9.98) to 6.5. Furthermore, the BDWG solution showed in mixing well after pretreatment. (See Table 1) since the water and soap were separated from the BDWG.

Table 1.
Characteristics of the BDWG before and after treatment.
Meanwhile, in Figure 3, the BDWG colour was obviously changed from dark brown and black into brown. Thus, the BDWG was ready for further use.

Figure 3.
(Left) the BDWG before treatment (middle) the BDWG separated from the soap layer when it was mixed in culture medium and (right) the BDWG after treatment.
3.3. Effect of the BDWG Concentration on Algal Biomass and DHA Accumulation
Before batch cultivation under the MC condition was performed, the preliminary test of optimal BDGW concentrations was varied at five levels (25, 50, 75, 100, 125 g/L) and considered. The results obtained (See Table 2) revealed that the maximal algal biomass, as well as the accumulated DHA in biomass, were obtained at 9.45 mg/L. However, the accumulated DHA was not much (357.288 mg/g). Therefore, the optimal products were 8.98 g/L of biomass and 510.527 mg/g of DHA when the BDGW was used at about 50 g/L. Therefore, the concentration of BDWG can be further used in both the flask and 5 L of the fermenter.
Table 2.
Biomass and DHA production by S. limacinum ATCC MYA-1381 under 5 levels of glycerol concentration.
3.4. Effect of Aeration Rates on Algal Biomass and DHA Production
An oxygen limitation has previously been reported in yeast culture [11]. To determine if oxygen is, in fact, a limiting factor, a range of aeration rates was investigated. Figure 4 shows the effect of aeration rates on cell growth performance and DHA production. As aeration increases, the algal growth and DHA production increase and reach an optimum at 0.25 vvm. A further increase in the aeration rate to 0.6 vvm led to a decline in both the algal growth and DHA production.
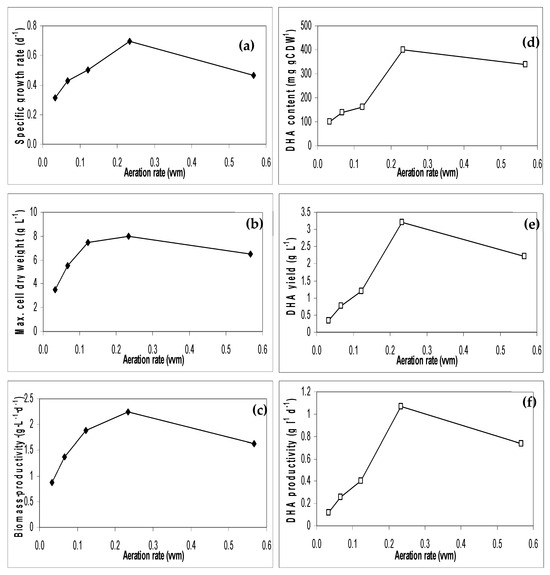
Figure 4.
Effect of aeration rate on cell growth performance (a–c) and DHA production (d–f) by S. limacinum on 2% glucose (Triplication).
The maximum biomass of 8 g/L, corresponding to the highest DHA yield of approximately 3.2 mg/L, was obtained at an aeration rate of 0.25 vvm. The increased biomass concentrations at higher aeration rates were also observed by other workers [3,18]. In addition, Chi et al., 2009 [3] illustrated that high oxygen levels were favourable to cell reproduction. High aeration rates intensified cell respiration and energy metabolism, which could trigger more carbon flux flow to the tricarboxylic acid cycle. The decline in algal cell growth at aeration above the optimum rate could be possibly caused by either nutrient limitation or inhibition. Therefore, appropriate aeration adjustment should be adopted to promote the efficiency of substrate utilisation efficiency in pilot-scale production.
3.5. Effect of Initial Glycerol Waste Concentration
The effect of initial glycerol waste concentrations on algal growth and DHA production was investigated at the optimum aeration rate of 0.25 vvm. As shown in Figure 5, in the lower range of crude glycerol concentrations (below 50 g/L), the algal growth rates and DHA production increased and reached an optimum level at 50 g/L. At glycerol concentrations above 50 g/L, the growth and DHA production were significantly inhibited. An explanation for these results may be as follows: (1) substrate inhibition as a low concentration of the substrate stimulates cell growth, whereas a high concentration of the substrate has an inhibitory effect. This is supported by the fact that during the first two days, the fastest biomass increase at 0.25 g/L glycerol concentration was achieved compared to the other concentrations; (2) salt content. S. limacinum is a marine alga. The medium used for its growth was 25 g/L NaCl. When this medium was used in conjunction with various glycerol concentrations, the overall NaCl concentration for each glycerol concentration was different.
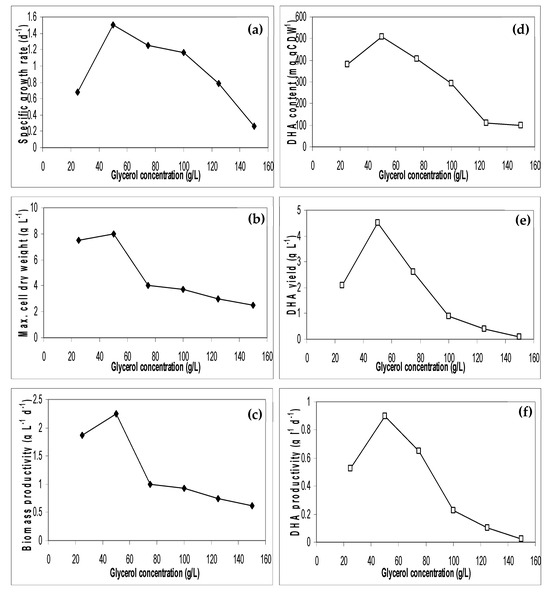
Figure 5.
Influence of glycerol concentrations on cell growth performance (a–c) and DHA production (d–f) using S. limacinum (Triplication).
The presence of an optimum glycerol concentration when S. limacinum is grown in a glycerol-containing medium is in agreement with other studies [2,7,17,31,32] (See Table 3). The difference in the glycerol inhibition concentrations found in this study to those in the literature could arise from employing different sources for crude glycerol and the various levels of impurities present. The compositions of crude glycerol samples varied according to several publications [1,10,15,17]. This variation mainly resulted from different oil feedstocks and biodiesel production processes. Liang et al. (2010) [10] found lower optimal glycerol doses compared to others and attributed these observations to the impurities in yellow grease-derived glycerol, which do not exist in soybean oil-derived crude glycerol.
Table 3.
Influence of various sources of crude glycerol concentrations on cell growth and DHA productions observed in the literature as well as using S. limacinum in this study.
3.6. Effect of Carbon Sources
Cell growth kinetics and DHA production by S. limacinum on different substrates during heterotrophic cultivation in a 5 L fermenter were further compared (see Table 4 and Table 5). The cells grew very well in crude glycerol compared to glucose and pure glycerol. The cell dry weight reached the highest level on days 5–6. The highest DHA production of circa 586 mg/g cell dry weight was observed in crude glycerol waste and glucose. Slightly lower levels of DHA production were observed in commercial glycerol. Similar trends in algal growth rates were also obtained for the different carbon sources. For a typical heterotrophic algal culture, glucose is usually expected as an easier-consumed carbon source versus other substrates. This is true when comparisons are made between pure glycerol and glucose. The higher specific cell growth rate obtained in crude glycerol, compared to that observed in pure glycerol, could be attributed to impurities, which may have beneficial effects on cell growth. The influence of the carbon source on cell growth and DHA production is in agreement with Chi et al. (2007) [2]. Elsewhere, the specific growth, lipid, and DHA productivities in 50% sweet sorghum juice-grown S. limacinum SR21 cells were reported to be higher than those from glucose-grown cells [10].
Table 4.
Comparison of cell growth and the DHA production of S. limacinum using different substrates.
Table 5.
Kinetic cultivation of S. limacinum ATCC MYA -1381 in the 5 L fermenter.
Figure 6 shows the cultivation of S. limacinum ATCC MYA-1381 in a 5 L fermenter under the HC or dark condition (for 24 h and until 240 h). It was noted that the fermenter was wrapped in black cloth.

Figure 6.
Cultivation of S. limacinum ATCC MYA-1381 in a 5 L fermenter under the dark condition.
Figure 7 shows the kinetic between algal biomass and DHA production during aerobic batch fermentation. It was revealed that the maximum biomass was obtained at 11.40 g at 120 h while the maximum DHA in algal biomass was achieved at 665.52 mg/g, which was equivalent to 6.27 g of the DHA yield/g BDWG.
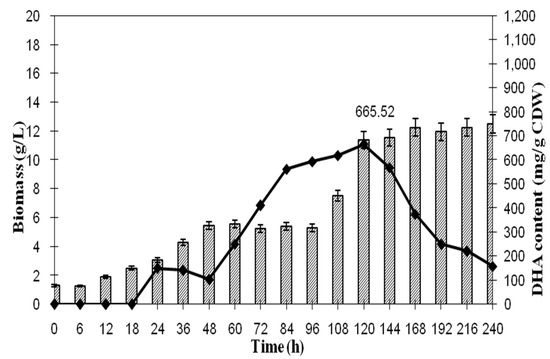
Figure 7.
The kinetic of biomass and DHA content as a function of time.
4. Conclusions
The potential of utilising the BDGW derived from WCO as a carbon source for the production of DHA via marine microalga and aerobic batch cultivation was studied. The cell growth and DHA content obtained from Schizochytrium limacinum ATCC MYA-1381 were shown to be affected by various operating conditions, such as aeration rates and the BDGW concentrations. The optimal culture conditions obtained in this study form the basis for future pilot scale work involving the optimisation of a mass algal production process for an accumulated high DHA yield via other types of fermentation, including a fed-batch, repeated fed-batch, and even a continuous system under the AC/HC and MC cultivations.
Author Contributions
This article was performed as collaborative research between V.T. and P.E. as well as P.K. For the first two authors, their work included the following: conceptualisation, resources, methodology, formal analysis, data curation, visualisation and writing—original draft preparation, P.E., considered and performed the experiment, the software, validation and investigation while, P.K. performed the main project administration, funding acquisition, review and corrections. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Khon Kaen University, Khon Kaen, Thailand, for partially article processing charge (ACP) cost and for collaborative research with P.E. (Newcastle University, Merz Court, Newcastle upon Tyne, UK).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are contained within the article.
Acknowledgments
The authors would like to thank a pilot plant of biodiesel production, Faculty of Engineering, Khon Kaen University, Thailand, for providing the crude glycerol samples. Further acknowledgements go to laboratories of the Faculty of Science and Technology, Nakhon Ratchasima Rajabhat University, and the Post-harvest and Processing Research and Development Division, Ministry of Agriculture and Cooperative for the facilities and their support in the analysis of the samples.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| AA | Arachidonic acid |
| AC | Autotrophic cultivation |
| ALA | Alpha linoleic acid |
| ATCC | American Type Culture Collection |
| ATCC MYA-1381 | Schizochytrium limacinum |
| BGDW | Biodiesel-derived glycerol waste |
| CDW | Cell dry weight |
| DHA | Docohexaenoic acid |
| DGLA | Homo gamma linoleic acid |
| EPA | Eicosapeentaenoic acid |
| HC | Heterotrophic cultivation |
| FAMEs | Fatty acid methyl esters |
| FID | Flame Ionisation Detector |
| GC | Gas chromatography |
| GPY | Glucose–Peptone–Yeast |
| LA | Linoleic acid |
| MC | Mixotrophic cultivation |
| PUAFs | Polyunsaturated fatty acids |
| v/v | Volume by volume |
| v/v/m | Volume by volume by minute |
| WCO | Waste cooking oil |
| w/v | Weight by volume |
| w/w | Weight by weight |
References
- Saman, A.-u.-R.; Wijesekara, R.G.; Nomura, N.; Sato, S.; Matsumura, M. Pre-treatment and utilization of raw glycerol from sunflower oil biodiesel for growth and 1,3-propanediol production by Clostridium butyricum. J. Chem. Technol. Biotechnol. 2008, 83, 1072–1080. [Google Scholar]
- Chi, Z.; Pyle, D.; Wen, Z.; Frear, C.; Chen, S. A laboratory study of producing docosahexaenoic acid from biodiesel-waste glycerol by microalgal fermentation. Process Biochem. 2007, 42, 1537–1545. [Google Scholar] [CrossRef]
- Chi, Z.Y.; Liu, Y.; Frear, C.; Chen, S.L. Study of a two-stage growth of DHA-producing marine algae Schizochytrium limacinum SR21 with shifting dissolved oxygen level. Appl. Microbiol. Biotechnol. 2009, 81, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.W.; Dasari, M.A.; Sutterlin, W.R.; Suppes, G.J. Removal of residual catalyst from simulated biodiesel’s crude glycerol for glycerol hydro-genolysis to propylene glycol. Ind. Eng. Chem. Res. 2006, 45, 791–795. [Google Scholar] [CrossRef]
- Dasari, M.A.; Kiatsimkul, P.-P.; Sutterlin, W.R.; Suppes, G.J. Low-pressure hydrogenolysis of glycerol to propylene glycol. Appl. Catal. A Gen. 2005, 281, 225–231. [Google Scholar] [CrossRef]
- Dharmadi, Y.; Murarka, A.; Gonzalez, R. Anaerobic fermentation of glycerol by Escherichia coli: A new platform for metabolic engineering. Biotechnol. Bioeng. 2006, 94, 821–829. [Google Scholar] [CrossRef]
- Ethier, S.; Woisard, K.; Vaughan, D.; Wen, Z. Continuous culture of the microalgae Schizochytrium limacinum on biodiesel-derived crude glycerol for producing docosahexaenoic acid. Bioresour. Technol. 2011, 102, 88–93. [Google Scholar] [CrossRef]
- Johnson, D.T.; Taconi, K.A. The glycerin glut: Options for the value-added conversion of crude glycerol resulting from biodiesel production. Environ. Prog. 2007, 26, 338–348. [Google Scholar] [CrossRef]
- Johnson, M.B.; Wen, Z. Production of biodiesel fuel from the microalga Schizochytrium limacinum by direct transesterification of algal biomass. Energy Fuel 2009, 10, 5179–5183. [Google Scholar] [CrossRef]
- Liang, Y.; Sarkany, N.; Cui, Y.; Blackburn, J.W. Batch stage study of lipid production from crude glycerol derived from yellow grease or animal fats through microalgal fermentation. Bioresour. Technol. 2010, 101, 6745–6750. [Google Scholar] [CrossRef]
- Liu, Y.-S.; Wu, J.-Y.; Ho, K.-P. Characterization of oxygen transfer conditions and their effects on Phaffia rhodozyma growth and carotenoid production in shake-flask cultures. Biochem. Eng. J. 2006, 27, 331–335. [Google Scholar] [CrossRef]
- Indarti, E.; Majid, M.I.A.; Hashim, R.; Chong, A. Direct FAME synthesis for rapid total lipid analysis from fish oil and cod liver oil. J. Food Compos. Anal. 2005, 18, 161–170. [Google Scholar] [CrossRef]
- Meesters, P.A.E.P.; Huijberts, G.N.M.; Eggink, G. High-cell-density cultivation of the lipid accumulating yeast Cryptococcus curvatus using glycerol as a carbon source. Appl. Microbiol. Biotechnol. 1996, 45, 575–579. [Google Scholar] [CrossRef]
- Narayan, M.S.; Manoj, G.P.; Vatchravelu, K.; Bhagyalakshmi, N.; Mahadevaswamy, M. Utilization of glycerol as carbon source on the growth, pigment and lipid production in Spirulina platensis. Int. J. Food Sci. Nutr. 2005, 56, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Lipid production by Yarrowia lipolytica growing on industrial glycerol in a single-stage continuous culture. Bioresour. Technol. 2002, 82, 43–49. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Fakasb, S.; Ficka, M.; Chevalota, I.; Galiotou-Panayotoub, M.; Komaitisb, M.; Marca, I.; Aggelisc, G. Biotechnological valorisation of raw glycerol discharged after bio-diesel (fatty acid methyl esters) manufacturing process: Production of 1,3-propanediol, citric acid and single cell oil. Biomass Bioenergy 2008, 32, 60–71. [Google Scholar] [CrossRef]
- Pyle, D.J.; Garcia, R.A.; Wen, Z. Producing Docosahexaenoic Acid (DHA)-Rich Algae from Biodiesel-Derived Crude Glycerol: Effects of Impurities on DHA Production and Algal Biomass Composition. J. Agric. Food Chem. 2008, 56, 3933–3939. [Google Scholar] [CrossRef]
- Ren, L.J.; Ji, X.J.; Huang, H.; Qu, J.; Feng, Y.; Tong, Q.Q.; Ouyang, P.K. Development of a stepwise aeration control strategy for efficient docosahexaenoic acid production by Schizochytrium sp. Appl. Microbiol. Biotechnol. 2010, 87, 1649–1656. [Google Scholar] [CrossRef]
- Zhang, Y.; Ward, V.; Dennis, D.; Plechkova, N.V.; Armenta, R.; Rehmann, L. Efficient Extraction of a Docosahexaenoic Acid (DHA)-Rich Lipid Fraction from Thraustochytrium sp. Using Ionic Liquids. Materials 2018, 11, 1986. [Google Scholar] [CrossRef]
- Yang, Y.; Li, G.; Li, F.; Xu, F.; Hu, P.; Xie, Z.; Lu, X.; Ding, Y.; Wang, Z. Impact of DHA from Algal Oil on the Breast Milk DHA Levels of Lactating Women: A Randomized Controlled Trial in China. Nutrients 2022, 14, 3410. [Google Scholar] [CrossRef]
- Sittiprapaporn, P.; Bumrungpert, A.; Suyajai, P.; Stough, C. Effectiveness of fish oil-DHA supplementation for cognitive function in Thai children: A randomized, doubled-blind, two-dose, placebo-controlled clinical trial. Foods 2022, 11, 2595. [Google Scholar] [CrossRef]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.B.S.; Ciappolino, V.; Agostoni, C. DHA Effects in Brain Development and Function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Essential fatty acids in health and chronic disease. Am. J. Clin. Nutr. 1999, 70, 560S–569S. [Google Scholar] [CrossRef] [PubMed]
- Tyson, K.S.; Bozell, J.; Wallace, R.; Petersen, E.; Moens, L. Biomass Oil Analysis: Research Needs and Recommendations; NREL/TP-510-34796; National Renewable Energy Laboratory: Golden, CO, USA, 2004. [Google Scholar]
- Ursin, V.M. Modification of Plant Lipids for Human Health: Development of Functional Land-Based Omega-3 Fatty Acids. J. Nutr. 2003, 133, 4271–4274. [Google Scholar] [CrossRef] [PubMed]
- Bozzatello, P.; Blua, C.; Rocca, P.; Bellino, S. Mental Health in Childhood and Adolescence: The Role of Polyunsaturated Fatty Acids. Biomedicines 2021, 9, 850. [Google Scholar] [CrossRef]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72–76. [Google Scholar] [CrossRef]
- Liang, Y.; Cui, Y.; Trushenski, J.; Blackburn, J.W. Converting crude glycerol derived from yellow grease to lipids through yeast fermentation. Bioresour. Technol. 2010, 101, 7581–7586. [Google Scholar] [CrossRef]
- Wu, S.T.; Yu, S.T.; Lin, L.P. Effect of culture conditions on docosahexaenoic acid production by Schizochytrium sp. 31. Proc. Biochem. 2005, 40, 3103–3108. [Google Scholar] [CrossRef]
- Rattanapoltee, P.; Dujjanutat, P.; Muanruksa, P.; Kaewkannetra, P. Biocircular platform for third generation biodiesel production: Batch/fed batch mixotrophic cultivations of microalgae using glycerol waste as a carbon source. Biochem. Eng. J. 2021, 175, 108128. [Google Scholar] [CrossRef]
- Rattanapoltee, P.; Kaewkannetra, P. Cultivation of microalga, Chlorella vulgaris under different auto-hetero-mixo trophic growths as a raw material during biodiesel production and cost evaluation. Energy 2014, 78, 4–8. [Google Scholar] [CrossRef]
- Rattanapoltee, P.; Kaewkannetra, P. Utilization of Agricultural Residues of Pineapple Peels and Sugarcane Bagasse as Cost-Saving Raw Materials in Scenedesmus acutus for Lipid Accumulation and Biodiesel Production. Appl. Biochem. Biotechnol. 2014, 173, 1495–1510. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).