
A Two-Stage Cascade for Increased High-Value Product Accumulation in Chlamydomonas asymmetrica

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Organism: Chlamydomonas asymmetrica
2.2. Preparation Modified AF-6 Medium
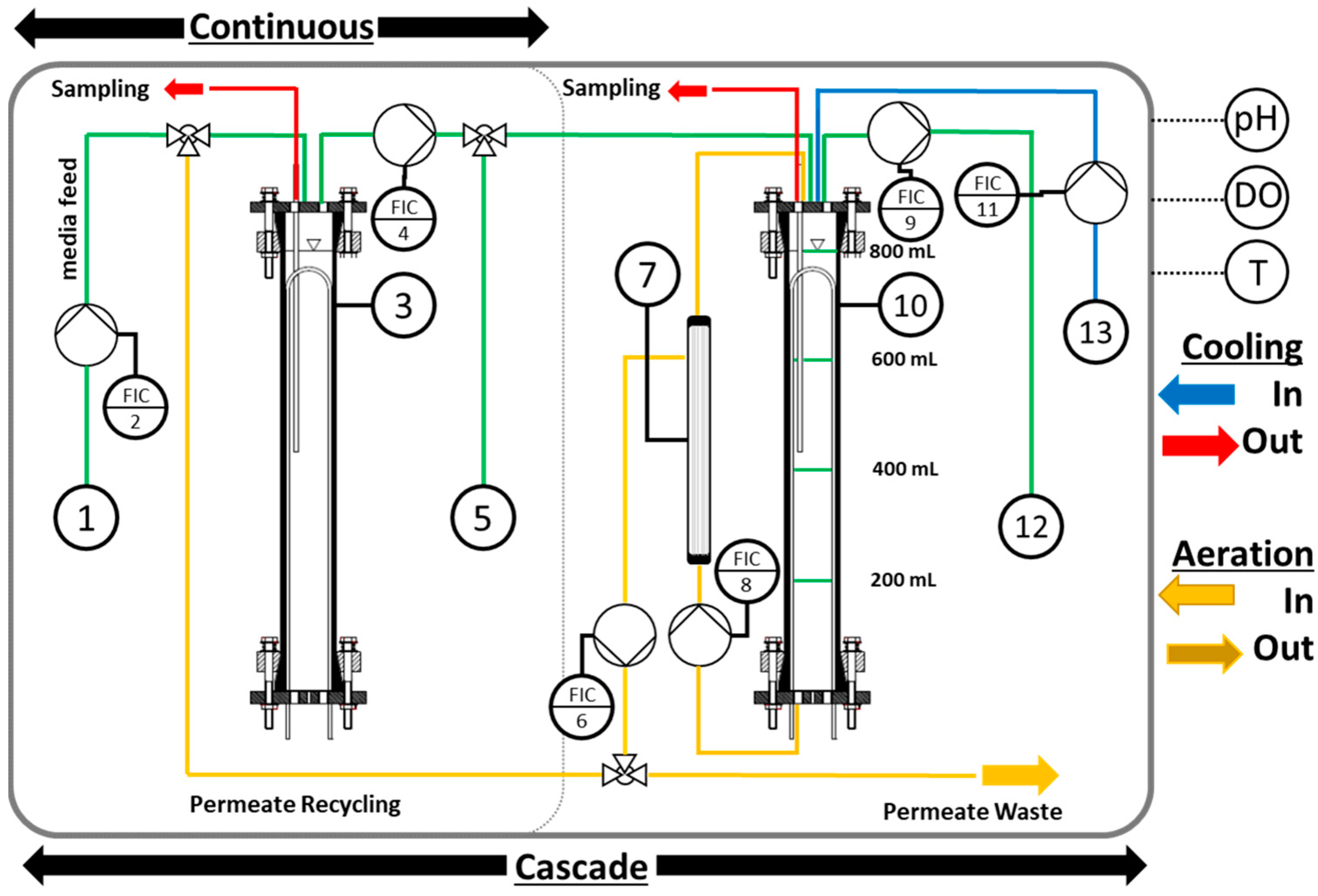
2.3. Reactor System Setup
2.4. Cultivation
2.5. Biomass and OD Quantification
2.6. Nitrate Analysis
2.7. High-Performance Liquid Chromatography (HPLC): Pigments
Gas Chromatography–Mass Spectrometry (GC-MS): Polyunsaturated Fatty Acids (PUFA)
2.8. Carbohydrate Content: EPS
3. Results and Discussion
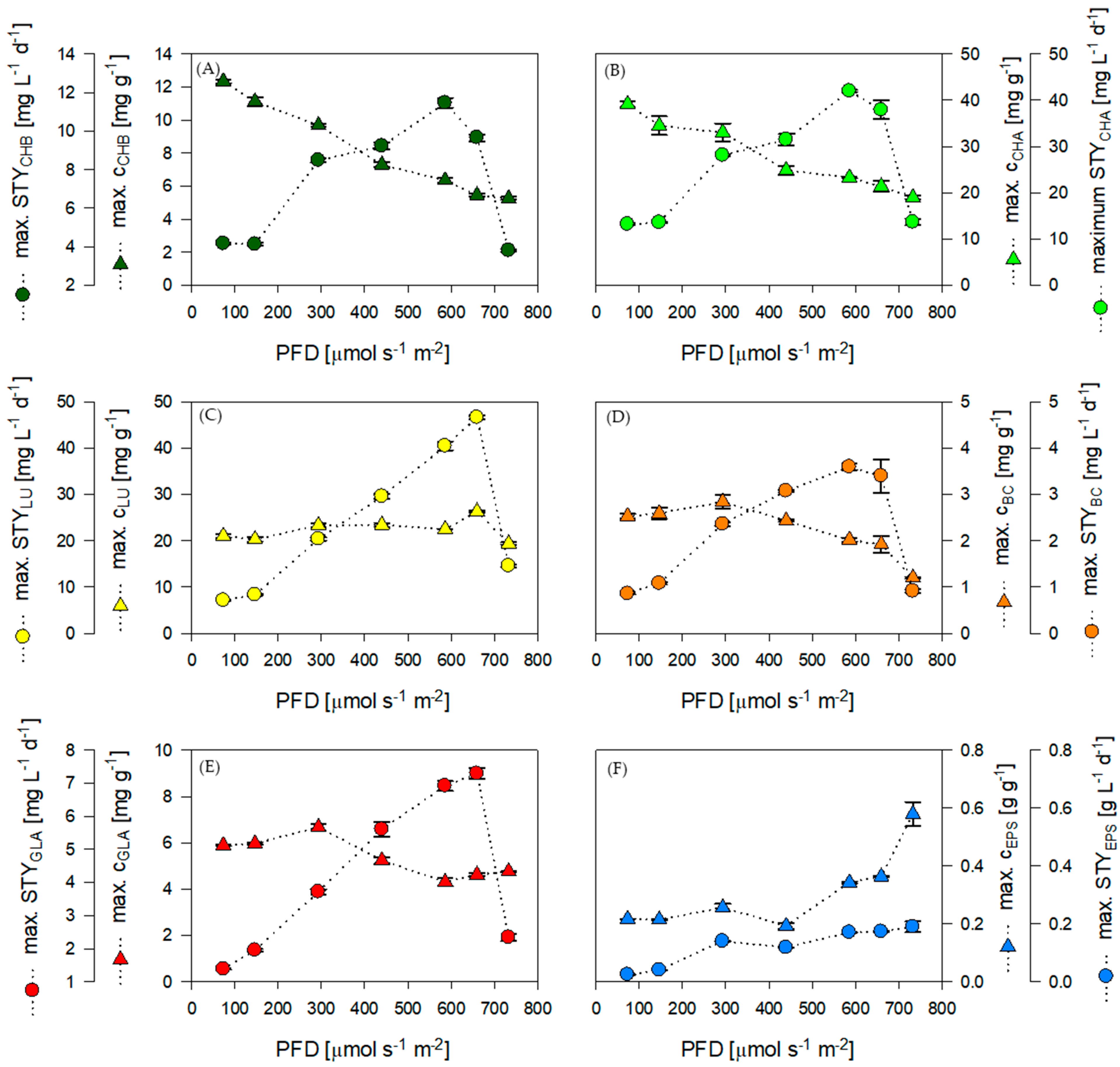
3.1. Chemostat Cultivation
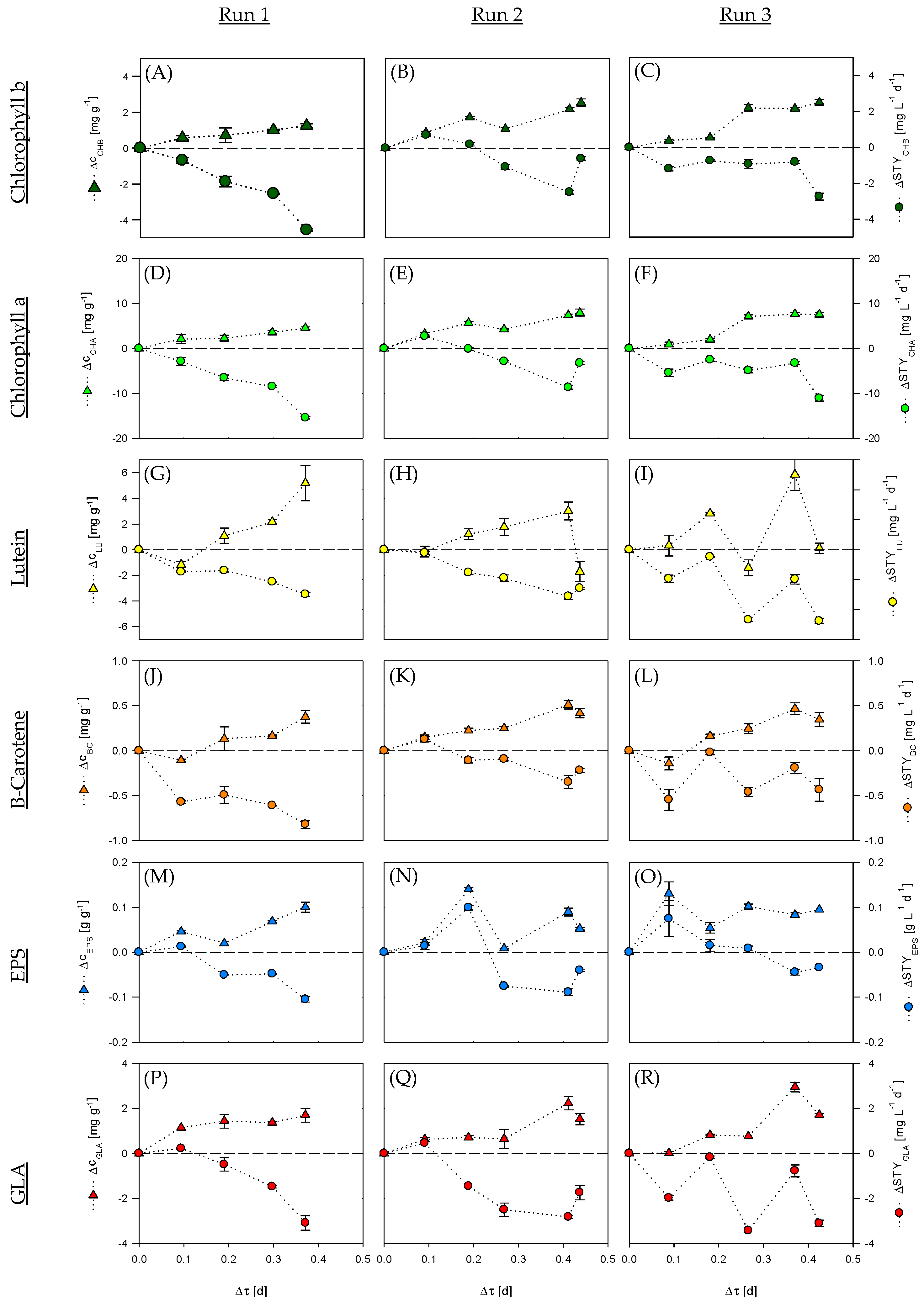
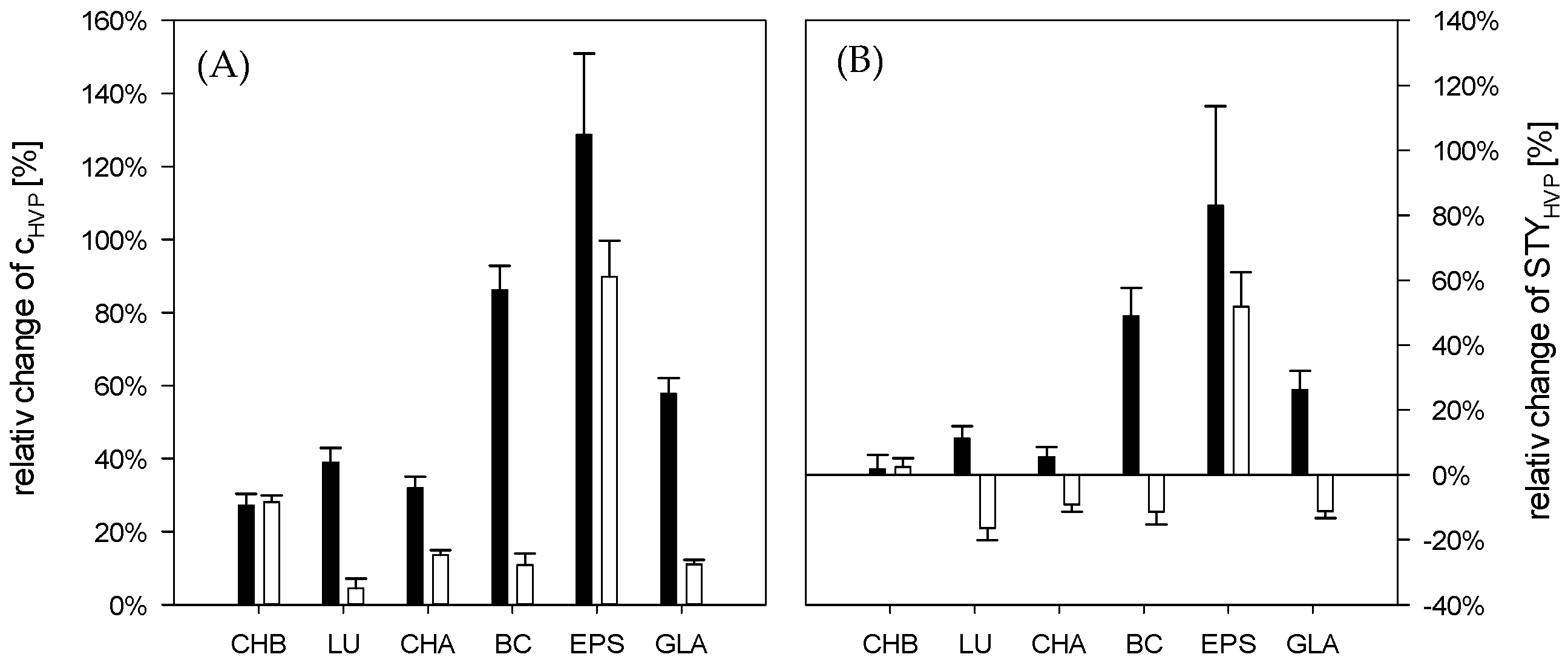
3.2. Cascade-System
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsygankov, A.; Solovchenko, A. “Microalgae as Converters of Light Energy into Biofuels and High-Value Products”: A Session of the Russian Photobiology Society 9th Congress (Shepsi, Krasnodar Region, Russia; September, 12–19, 2021). Biophys. Rev. 2022, 14, 761–763. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D. How Many Species of Algae Are There? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. High-Value Products from Microalgae-Their Development and Commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- McBride, R.C.; Lopez, S.; Meenach, C.; Burnett, M.; Lee, P.A.; Nohilly, F.; Behnke, C. Contamination Management in Low Cost Open Algae Ponds for Biofuels Production. Ind. Biotechnol. 2014, 10, 221–227. [Google Scholar] [CrossRef]
- Pulz, O. Photobioreactors: Production Systems for Phototrophic Microorganisms. Appl. Microbiol. Biotechnol. 2001, 57, 287–293. [Google Scholar] [CrossRef]
- Li, H.B.; Chen, F.; Zhang, T.Y.; Yang, F.Q.; Xu, G.Q. Preparative Isolation and Purification of Lutein from the Microalga Chlorella Vulgaris by High-Speed Counter-Current Chromatography. J. Chromatogr. A 2001, 905, 151–155. [Google Scholar] [CrossRef]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-Producing Green Microalga Haematococcus Pluvialis: From Single Cell to High Value Commercial Products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.J.; Zijffers, J.W.; Nisworo, A.; Vaes, W.; van Schoonhoven, J.; Wijffels, R.H. Optimization of Biomass, Vitamins, and Carotenoid Yield on Light Energy in a Flat-Panel Reactor Using the A-Stat Technique. Biotechnol. Bioeng. 2005, 89, 233–242. [Google Scholar] [CrossRef]
- Zijffers, J.W.F.; Schippers, K.J.; Zheng, K.; Janssen, M.; Tramper, J.; Wijffels, R.H. Maximum Photosynthetic Yield of Green Microalgae in Photobioreactors. Mar. Biotechnol. 2010, 12, 708–718. [Google Scholar] [CrossRef]
- Shin, W.S.; Lee, H.; Sung, M.G.; Hwang, K.T.; Jung, S.M.G.; Kwon, J.H. Enrichment as a Screening Method for a High-Growth-Rate Microalgal Strain under Continuous Cultivation System. Biotechnol. Bioprocess Eng. 2016, 21, 268–273. [Google Scholar] [CrossRef]
- Lee, Y.-K.; Shen, H. Basic Culturing Techniques. In Handbook of Microalgal Culture; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 40–56. [Google Scholar]
- Markou, G.; Nerantzis, E. Microalgae for High-Value Compounds and Biofuels Production: A Review with Focus on Cultivation under Stress Conditions. Biotechnol. Adv. 2013, 31, 1532–1542. [Google Scholar] [CrossRef] [PubMed]
- Panis, G.; Carreon, J.R. Commercial Astaxanthin Production Derived by Green Alga Haematococcus Pluvialis: A Microalgae Process Model and a Techno-Economic Assessment All through Production Line. Algal Res. 2016, 18, 175–190. [Google Scholar] [CrossRef]
- Sarada, R.; Tripathi, U.; Ravishankar, G.A. Influence of Stress on Astaxanthin Production in Haematococcus Pluvialis Grown under Different Culture Conditions. Process Biochem. 2002, 37, 623–627. [Google Scholar] [CrossRef]
- Lemoine, Y.; Schoefs, B. Secondary Ketocarotenoid Astaxanthin Biosynthesis in Algae: A Multifunctional Response to Stress. Photosynth. Res. 2010, 106, 155–177. [Google Scholar] [CrossRef]
- Ludwig, W.; Ludwig, W. Protozoa. Flagellata. In Das Rechts-Links-Problem im Tierreich und Beim Menschen; Springer: Berlin/Heidelberg, Germany, 1932; pp. 62–71. [Google Scholar]
- Hahm, J.; Bauer, A.; Jung, S.-H.; Zell, N.; Boßle, F.; Buchholz, R.; Lindenberger, C. Process Parameter Screening for the Microalga Chlamydomonas Asymmetrica in Batch and Turbidostat Cultivations. Chemie Ing. Tech. 2021, 93, 1565–1572. [Google Scholar] [CrossRef]
- Jung, S.-H.; Zell, N.; Boßle, F.; Teipel, U.; Rauh, C.; McHardy, C.; Lindenberger, C. Influence of Process Operation on the Production of Exopolysaccharides in Arthrospira Platensis and Chlamydomonas Asymmetrica. Front. Sustain. Food Syst. 2022, 6, 883069. [Google Scholar] [CrossRef]
- Watanabe, M.; Kasai, F.; Kawachi, M.; Erata, M.; Yumoto, K.; Sato, M. NIES-Collection List of Strains, 8th Edition. Jpn. J. Phycol 2009, 57, 1–350. [Google Scholar]
- Taucher, J.; Baer, S.; Schwerna, P.; Hofmann, D.; Hümmer, M.; Buchholz, R.; Becker, A. Cell Disruption and Pressurized Liquid Extraction of Carotenoids from Microalgae. J. Thermodyn. Catal. 2016, 7, 158. [Google Scholar] [CrossRef]
- Lewis, T.; Nichols, P.D.; McMeekin, T.A. Evaluation of Extraction Methods for Recovery of Fatty Acids from Lipid-Producing Microheterotrophs. J. Microbiol. Methods 2000, 43, 107–116. [Google Scholar] [CrossRef]
- Khoomrung, S.; Chumnanpuen, P.; Jansa-Ard, S.; Nookaew, I.; Nielsen, J. Fast and Accurate Preparation Fatty Acid Methyl Esters by Microwave-Assisted Derivatization in the Yeast Saccharomyces Cerevisiae. Appl. Microbiol. Biotechnol. 2012, 94, 1637–1646. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Takache, H.; Pruvost, J.; Cornet, J.-F. Kinetic Modeling of the Photosynthetic Growth of Chlamydomonas Reinhardtii in a Photobioreactor. Biotechnol. Prog. 2012, 28, 681–692. [Google Scholar] [CrossRef]
- Bonente, G.; Pippa, S.; Castellano, S.; Bassi, R.; Ballottari, M. Acclimation of Chlamydomonas Reinhardtii to Different Growth Irradiances. J. Biol. Chem. 2012, 287, 5833–5847. [Google Scholar] [CrossRef]
- Xie, S.-R.; Liang, M.-H.; Yan, B.; Jiang, J.-G. Creatinine Combined with Light Increases the Contents of Lutein and β-Carotene, the Main Carotenoids of Dunaliella Bardawil. Enzyme Microb. Technol. 2021, 151, 109913. [Google Scholar] [CrossRef]
- Solovchenko, A.E.; Khozin-Goldberg, I.; Didi-Cohen, S.; Cohen, Z.; Merzlyak, M.N. Effects of Light Intensity and Nitrogen Starvation on Growth, Total Fatty Acids and Arachidonic Acid in the Green Microalga Parietochloris Incisa. J. Appl. Phycol. 2008, 20, 245–251. [Google Scholar] [CrossRef]
- Hoober, J.K. The Chlamydomonas Sourcebook. A Comprehensive Guide to Biology and Laboratory Use. Elizabeth H. Harris. Academic Press, San Diego, CA, 1989. Xiv, 780 Pp., Illus. $145. Science 1989, 246, 1503–1504. [Google Scholar] [CrossRef] [PubMed]
- Kroen, W.K.; Rayburn, W.R. Influence of Growth Status and Nutrients on Extracellular Polysaccharide Synthesis by the Soil Alga Chlamydomonas Mexicana (Chlorophyceae). J. Phycol. 1984, 20, 253–257. [Google Scholar] [CrossRef]
- Kandilian, R.; Taleb, A.; Heredia, V.; Cogne, G.; Pruvost, J. Effect of Light Absorption Rate and Nitrate Concentration on TAG Accumulation and Productivity of Parachlorella Kessleri Cultures Grown in Chemostat Mode. Algal Res. 2019, 39, 101442. [Google Scholar] [CrossRef]
- Mogany, T.; Bhola, V.; Ramanna, L.; Bux, F. Photosynthesis and Pigment Production: Elucidation of the Interactive Effects of Nutrients and Light on Chlamydomonas Reinhardtii. Bioprocess Biosyst. Eng. 2022, 45, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Kamalanathan, M.; Pierangelini, M.; Shearman, L.A.; Gleadow, R.; Beardall, J. Impacts of Nitrogen and Phosphorus Starvation on the Physiology of Chlamydomonas Reinhardtii. J. Appl. Phycol. 2016, 28, 1509–1520. [Google Scholar] [CrossRef]
- Molina Grima, E.; Belarbi, E.H.; Acién Fernández, F.G.; Robles Medina, A.; Chisti, Y. Recovery of Microalgal Biomass and Metabolites: Process Options and Economics. Biotechnol. Adv. 2003, 20, 491–515. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PFD (1) [µmol·m−2·s−1] | PFD (2) [µmol·m−2·s−1] | V (2) [mL] | Nitrate Addition (2) | |
|---|---|---|---|---|
| Run 1 | 659 | 73 | 200–800 | No |
| Run 2 | 1319 | 73 | 200–900 | No |
| Run 3 | 1319 | 73 | 200–900 | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hahm, J.; Jung, S.-H.; Kandaswamy, S.; Talwar, P.; Verma, N.; Vivekanand, V.; Lindenberger, C. A Two-Stage Cascade for Increased High-Value Product Accumulation in Chlamydomonas asymmetrica. Fermentation 2024, 10, 38. https://doi.org/10.3390/fermentation10010038
Hahm J, Jung S-H, Kandaswamy S, Talwar P, Verma N, Vivekanand V, Lindenberger C. A Two-Stage Cascade for Increased High-Value Product Accumulation in Chlamydomonas asymmetrica. Fermentation. 2024; 10(1):38. https://doi.org/10.3390/fermentation10010038
Chicago/Turabian StyleHahm, Joachim, Sun-Hwa Jung, Saikrishnan Kandaswamy, Prakhar Talwar, Nikita Verma, Vivekanand Vivekanand, and Christoph Lindenberger. 2024. "A Two-Stage Cascade for Increased High-Value Product Accumulation in Chlamydomonas asymmetrica" Fermentation 10, no. 1: 38. https://doi.org/10.3390/fermentation10010038
APA StyleHahm, J., Jung, S.-H., Kandaswamy, S., Talwar, P., Verma, N., Vivekanand, V., & Lindenberger, C. (2024). A Two-Stage Cascade for Increased High-Value Product Accumulation in Chlamydomonas asymmetrica. Fermentation, 10(1), 38. https://doi.org/10.3390/fermentation10010038