
Self-Assembled Peptide Habitats to Model Tumor Metastasis

 , , ,
, , ,  , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
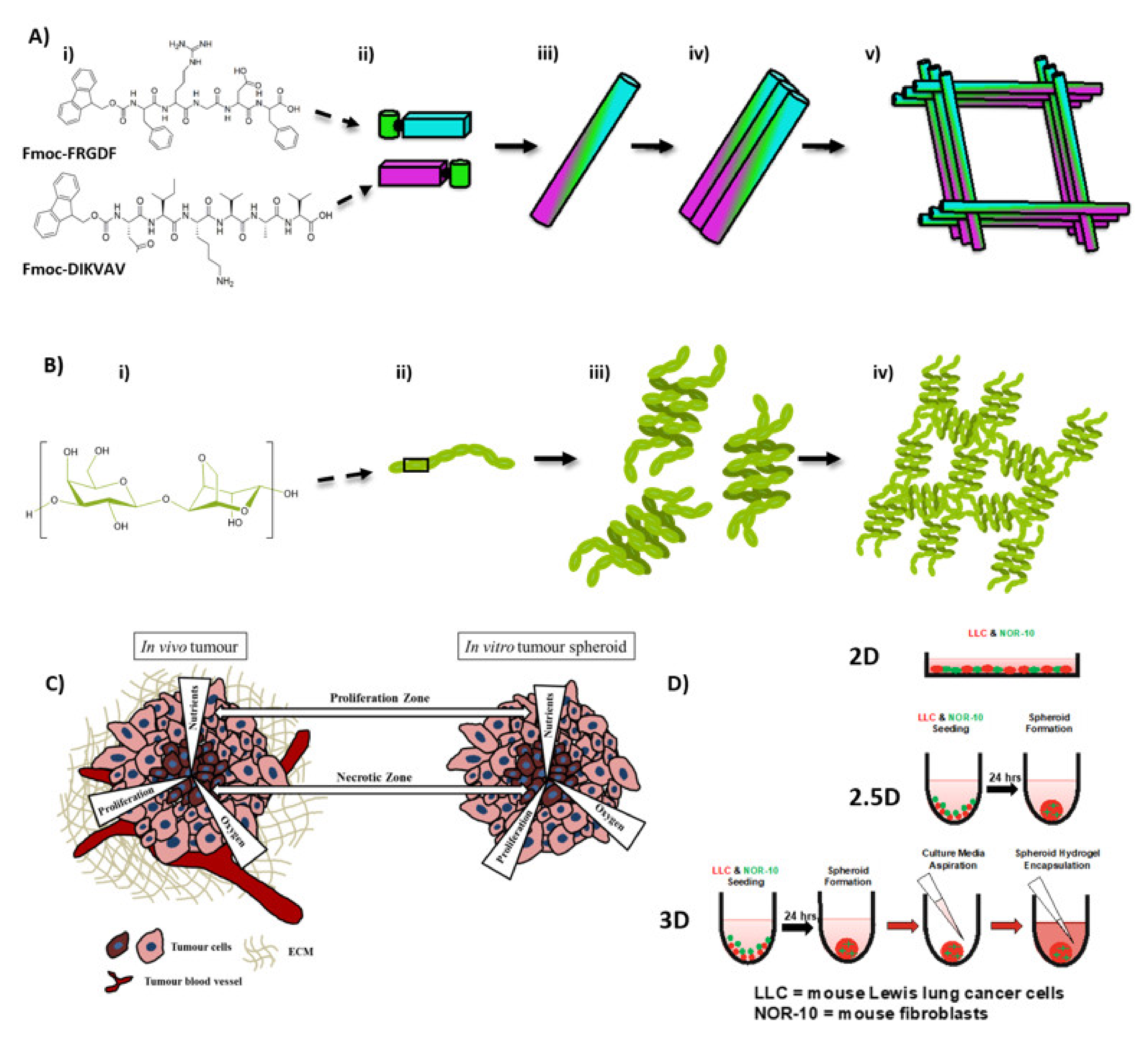
2.1. Co-Assembled Fmoc-FRGDF/Fmoc-DIKVAV Hydrogel Preparation
2.2. Agarose Preparation
2.3. Cryo-Scanning Electron Microscopy (CryoSEM)
2.4. Small-Angle X-ray Scattering (SAXS)
2.5. Cell Culture: LLC and NOR10
2.6. Culture Conditions in 2D, 2.5D and 3D Spheroids
2.7. Metabolic Activity
2.8. Flow Cytometry
2.9. Cell Migration Tracking
2.10. Immunofluorescence Staining for Vinculin and F-Actin
2.11. Statistical Analysis
3. Results and Discussion
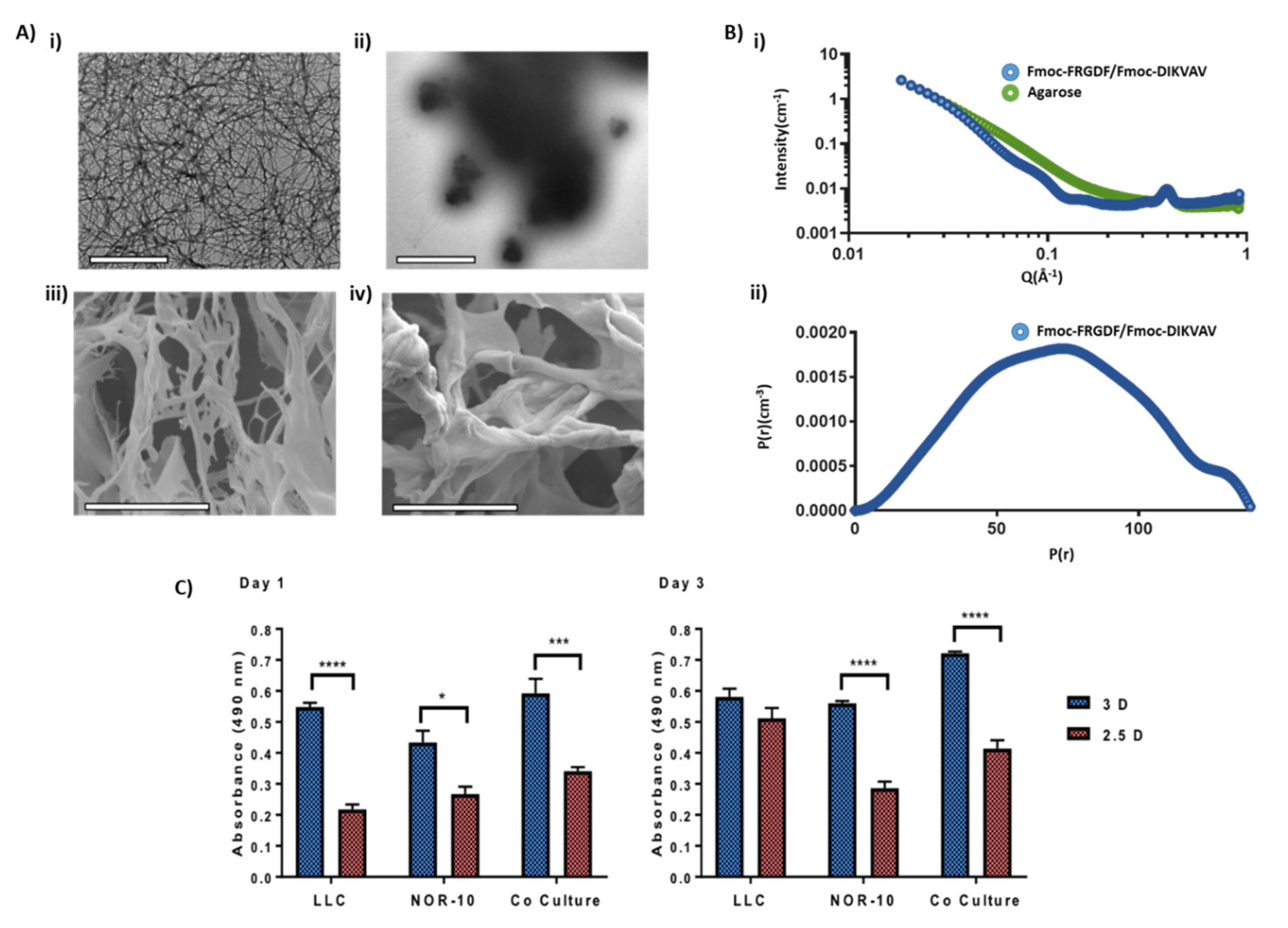
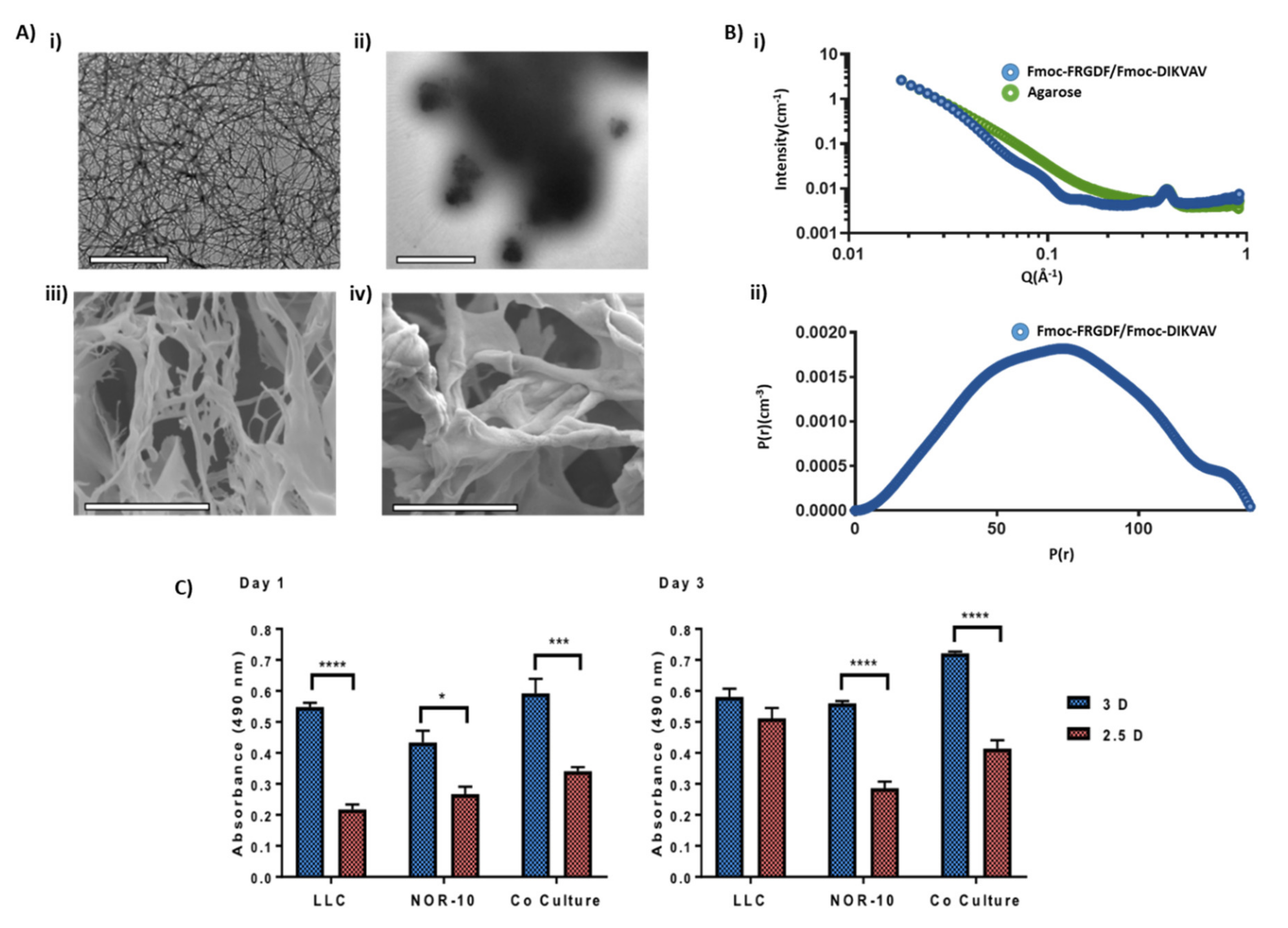
3.1. Comparison of Hydrogel Networks
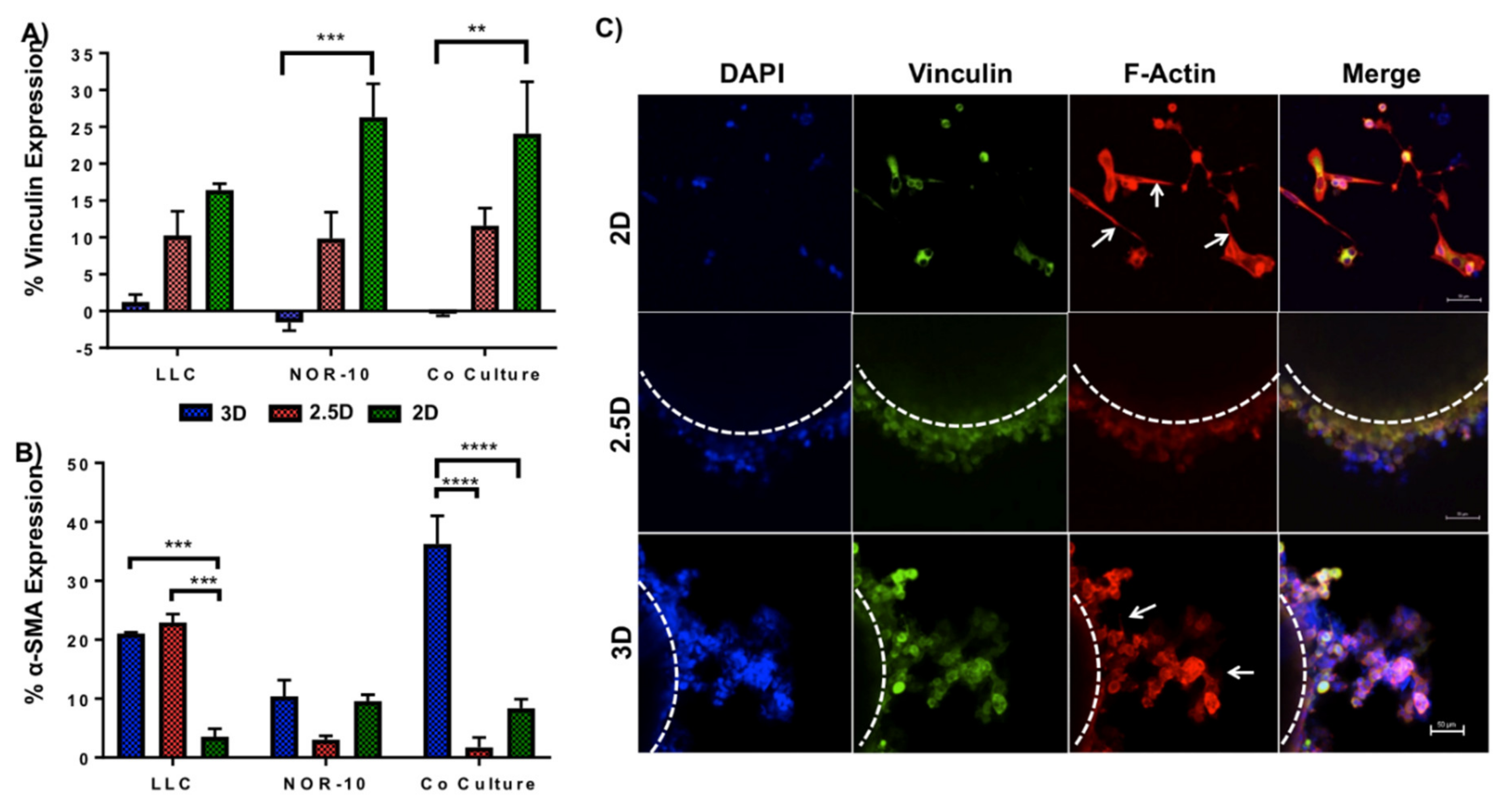
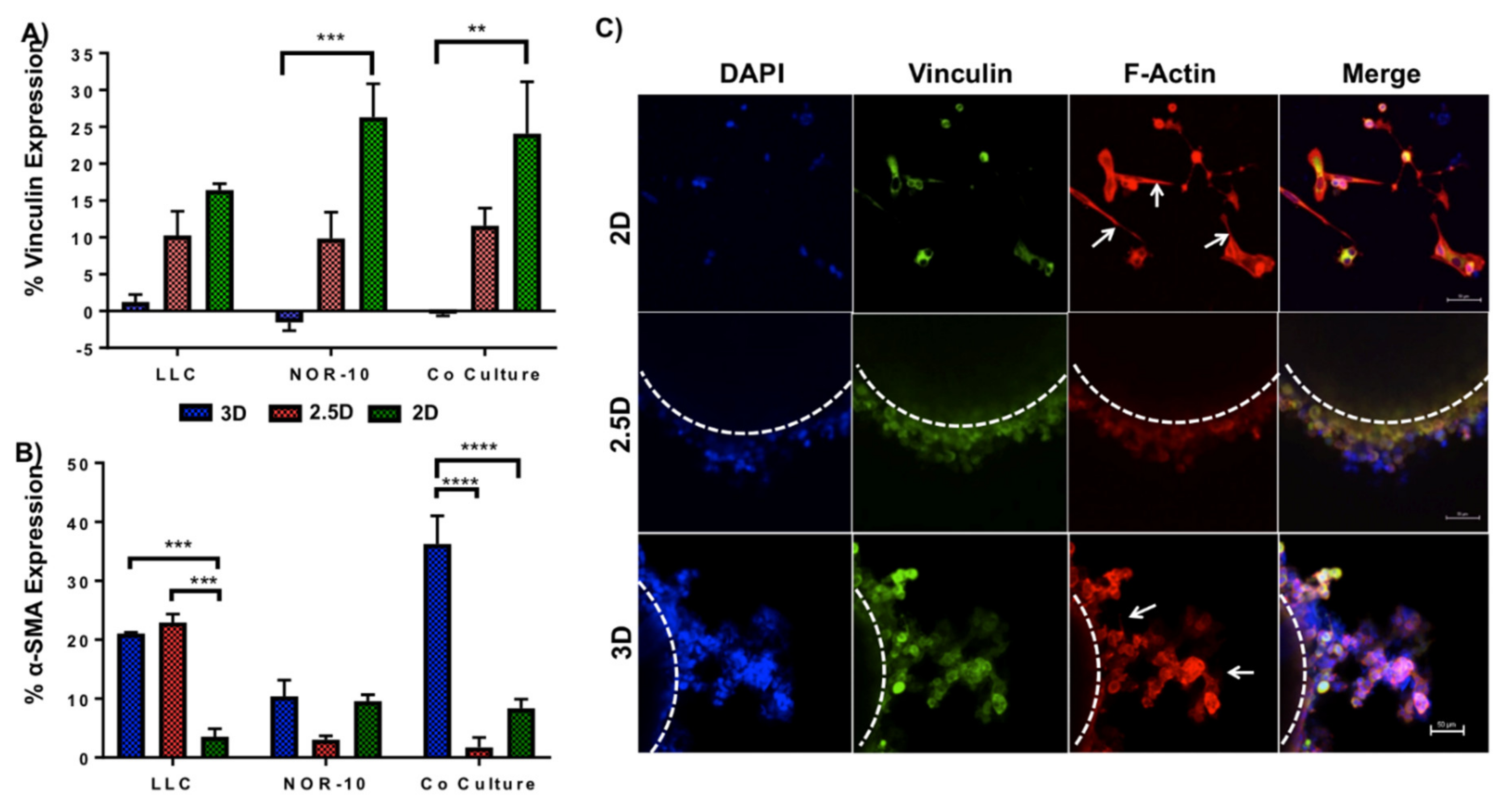
3.2. Evaluating Metabolic Activity of Lung Cancer and Stromal Cells Spheroids
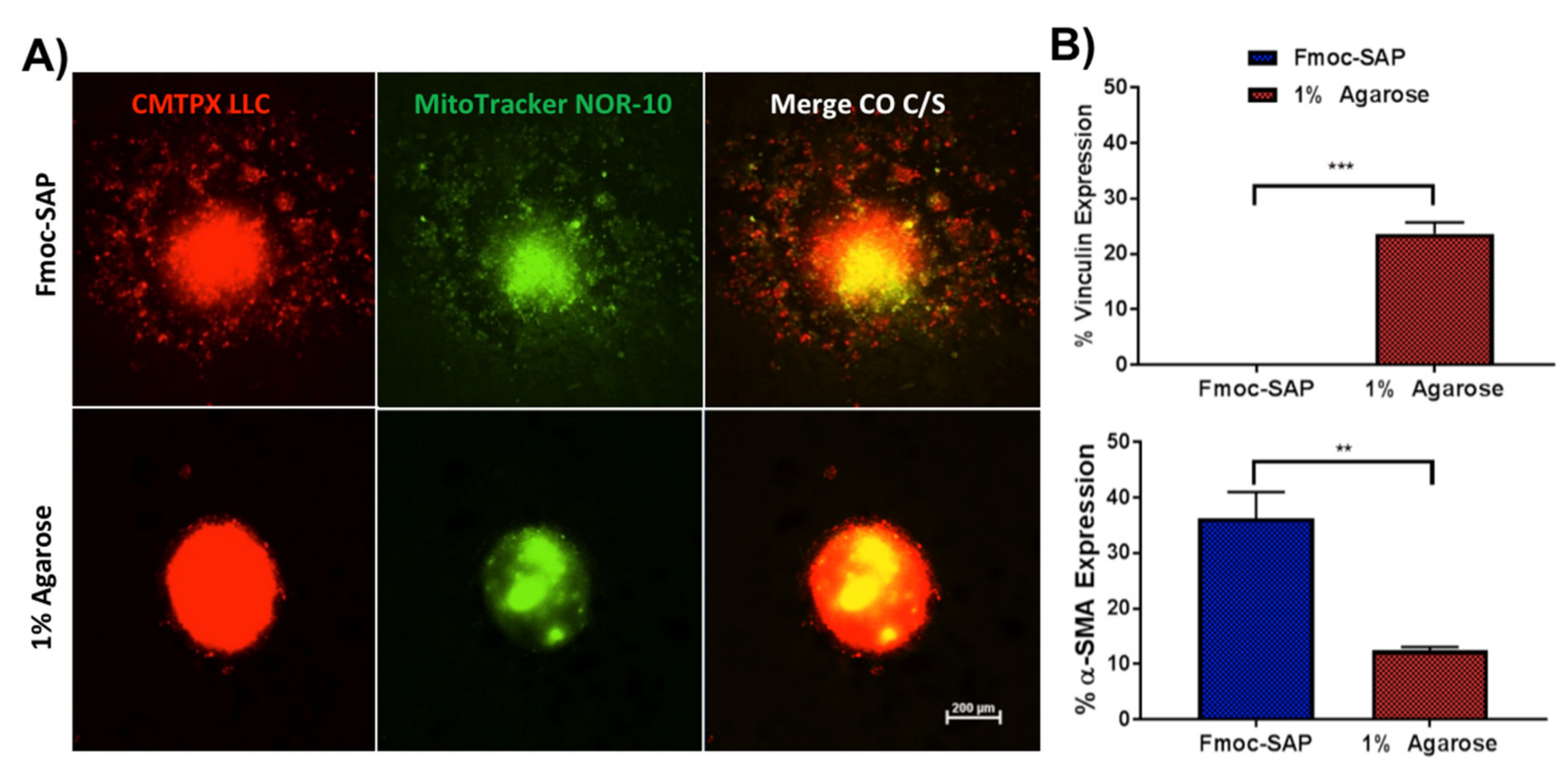
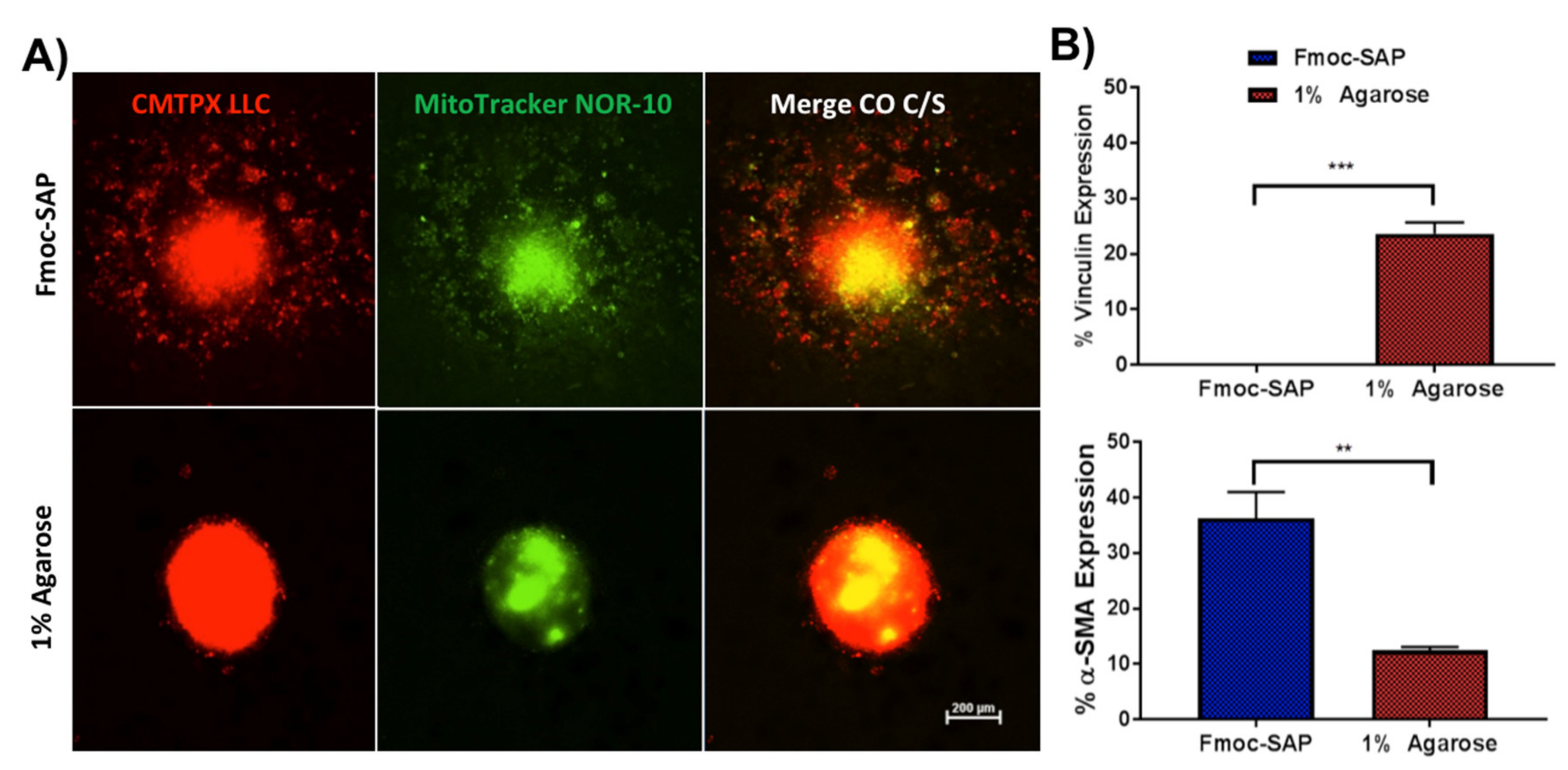
3.3. Enhanced Spheroid Adhesion within Functionalised Fmoc-SAP Hydrogel
3.4. Functionalised Fmoc-SAP Hydrogel Facilitates Lung Cancer Cell Migration
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sounni, N.E.; Noel, A. Targeting the tumor microenvironment for cancer therapy. Clin. Chem. 2013, 59, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Dart, A. Tumour microenvironment: Radical changes. Nat. Rev. Cancer 2018, 18, 65. [Google Scholar] [CrossRef]
- Antoni, D.; Burckel, H.; Josset, E.; Noel, G. Three-dimensional cell culture: A breakthrough in vivo. Int. J. Mol. Sci. 2015, 16, 5517–5527. [Google Scholar] [CrossRef]
- Jaganathan, H.; Gage, J.; Leonard, F.; Srinivasan, S.; Souza, G.R.; Dave, B.; Godin, B. Three-dimensional in vitro co-culture model of breast tumour using magnetic levitation. Sci. Rep. 2014, 4, 6468. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Farach-Carson, M.C.; Jia, X. Three-dimensional in vitro tumor models for cancer research and drug evaluation. Biotechnol. Adv. 2014, 32, 1256–1268. [Google Scholar] [CrossRef] [Green Version]
- Werb, Z.; Lu, P. The Role of Stroma in Tumor Development. Cancer J. 2015, 21, 250–253. [Google Scholar] [CrossRef] [Green Version]
- Thakuri, P.S.; Liu, C.; Luker, G.D.; Tavana, H. Biomaterials-Based Approaches to Tumor Spheroid and Organoid Modeling. Adv. Healthc. Mater. 2018, 7, 1700980. [Google Scholar] [CrossRef]
- Kim, C.H.; Suhito, I.R.; Angeline, N.; Han, Y.; Son, H.; Luo, Z.; Kim, T.H. Vertically Coated Graphene Oxide Micro-Well Arrays for Highly Efficient Cancer Spheroid Formation and Drug Screening. Adv. Healthc. Mater. 2020, 9, 1901751. [Google Scholar] [CrossRef]
- Charoen, K.M.; Fallica, B.; Colson, Y.L.; Zaman, M.H.; Grinstaff, M.W. Embedded multicellular spheroids as a biomimetic 3D cancer model for evaluating drug and drug-device combinations. Biomaterials 2014, 35, 2264–2271. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.-W.; Han, H.-W.; Tien, Y.-W.; Hsu, S.-h. Biomaterial substrate-derived compact cellular spheroids mimicking the behavior of pancreatic cancer and microenvironment. Biomaterials 2019, 213, 119202. [Google Scholar] [CrossRef]
- Burgos-Panadero, R.; Lucantoni, F.; Gamero-Sandemetrio, E.; Cruz-Merino, L.D.L.; Álvaro, T.; Noguera, R. The tumour microenvironment as an integrated framework to understand cancer biology. Cancer Lett. 2019, 461, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Meyer, C.M.; Joung, D.; Vallera, D.A.; McAlpine, M.C.; Panoskaltsis-Mortari, A. 3D bioprinted in vitro metastatic models via reconstruction of tumor microenvironments. Adv. Mater. 2019, 31, 1806899. [Google Scholar] [CrossRef] [PubMed]
- Bruggeman, K.; Zhang, M.; Malagutti, N.; Dehnavi, S.S.; Williams, R.; Tricoli, A.; Nisbet, D. Using UV-Responsive Nanoparticles to Provide In Situ Control of Growth Factor Delivery and a More Constant Release Profile from a Hydrogel Environment. ACS Appl. Mater. Interfaces 2022, 14, 12068–12076. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.; Kundu, B.; Silva-Correia, J.; Kundu, S.C.; Oliveira, J.M.; Reis, R.L.; Correlo, V.M. Emerging tumor spheroids technologies for 3D in vitro cancer modeling. Pharmacol. Ther. 2018, 184, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Finnberg, N.K.; Gokare, P.; Lev, A.; Grivennikov, S.I.; MacFarlane, A.W.; Campbell, K.S.; Winters, R.M.; Kaputa, K.; Farma, J.M.; Abbas, A.E.-S.; et al. Application of 3D tumoroid systems to define immune and cytotoxic therapeutic responses based on tumoroid and tissue slice culture molecular signatures. Oncotarget 2017, 8, 66747–66757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Hu, W.; Matulay, J.T.; Silva, M.V.; Owczarek, T.B.; Kim, K.; Chua, C.W.; Barlow, L.J.; Kandoth, C.; Williams, A.B. Tumor evolution and drug response in patient-derived organoid models of bladder cancer. Cell 2018, 173, 515–528. [Google Scholar] [CrossRef] [Green Version]
- Dwyer, A.R.; Ellies, L.G.; Holme, A.L.; Pixley, F.J. A three-dimensional co-culture system to investigate macrophage-dependent tumor cell invasion. J. Biol. Methods 2016, 3, e49. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Chaimov, D.; Patel, S.N.; Liang, J.P.; Wiggins, S.C.; Samojlik, M.M.; Rubiano, A.; Simmons, C.S.; Stabler, C.L. 3-D physiomimetic extracellular matrix hydrogels provide a supportive microenvironment for rodent and human islet culture. Biomaterials 2019, 198, 37–48. [Google Scholar] [CrossRef]
- Fang, Y.; Eglen, R.M. Three-Dimensional Cell Cultures in Drug Discovery and Development. SLAS Discov. 2017, 22, 456–472. [Google Scholar] [CrossRef] [Green Version]
- Worthington, P.; Pochan, D.J.; Langhans, S.A. Peptide Hydrogels—Versatile Matrices for 3D Cell Culture in Cancer Medicine. Front. Oncol. 2015, 5, 92. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.E.; Bahlmann, L.C.; Tam, R.Y.; Liu, J.C.; Ganesh, A.N.; Mitrousis, N.; Marcellus, R.; Spears, M.; Bartlett, J.M.; Cescon, D.W. Benchmarking to the Gold Standard: Hyaluronan-Oxime Hydrogels Recapitulate Xenograft Models with In Vitro Breast Cancer Spheroid Culture. Adv. Mater. 2019, 31, 1901166. [Google Scholar] [CrossRef] [PubMed]
- Lan, S.F.; Safiejko-Mroczka, B.; Starly, B. Long-term cultivation of HepG2 liver cells encapsulated in alginate hydrogels: A study of cell viability, morphology and drug metabolism. Toxicol. Vitr. 2010, 24, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Silva, T.L.; Leach, D.G.; Azares, A.; Li, I.C.; Woodside, D.G.; Hartgerink, J.D. Chemical functionality of multidomain peptide hydrogels governs early host immune response. Biomaterials 2020, 231, 119667. [Google Scholar] [CrossRef]
- Kim, B.J.; Yang, D.; Xu, B. Emerging Applications of Supramolecular Peptide Assemblies. Trends Chem. 2020, 2, 71–83. [Google Scholar] [CrossRef]
- Feng, Z.; Han, X.; Wang, H.; Tang, T.; Xu, B. Enzyme-Instructed Peptide Assemblies Selectively Inhibit Bone Tumors. Chem 2019, 5, 2442–2449. [Google Scholar] [CrossRef]
- Zhou, J.; Du, X.; Berciu, C.; He, H.; Shi, J.; Nicastro, D.; Xu, B. Enzyme-Instructed Self-Assembly for Spatiotemporal Profiling of the Activities of Alkaline Phosphatases on Live Cells. Chem 2016, 1, 246–263. [Google Scholar] [CrossRef] [Green Version]
- Hellmund, K.S.; Koksch, B. Self-Assembling Peptides as Extracellular Matrix Mimics to Influence Stem Cell’s Fate. Front. Chem. 2019, 7, 172. [Google Scholar] [CrossRef]
- Hainline, K.M.; Gu, F.; Handley, J.F.; Tian, Y.F.; Wu, Y.; Wet, L.D.; Griend, D.J.V.; Collier, J.H. Self-Assembling Peptide Gels for 3D Prostate Cancer Spheroid Culture. Macromol. Biosci. 2019, 19, 1800249. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.L.; Wang, T.Y.; Bruggeman, K.F.; Horgan, C.C.; Li, R.; Williams, R.J.; Parish, C.L.; Nisbet, D.R. In vivo assessment of grafted cortical neural progenitor cells and host response to functionalized self-assembling peptide hydrogels and the implications for tissue repair. J. Mater. Chem. B 2014, 2, 7771–7778. [Google Scholar] [CrossRef]
- Rodriguez, A.; Wang, T.; Bruggeman, K.; Li, R.; Williams, R.; Parish, C.; Nisbet, D. Tailoring minimalist self-assembling peptides for localised viral vector gene delivery. Nano Res. 2016, 9, 674–684. [Google Scholar] [CrossRef]
- Rodriguez, A.L.; Parish, C.L.; Nisbet, D.R.; Williams, R.J. Tuning the amino acid sequence of minimalist peptides to present biological signals via charge neutralised self assembly. Soft Matter. 2013, 9, 3915–3919. [Google Scholar] [CrossRef] [Green Version]
- Short, S.M.; Talbott, G.A.; Juliano, R.L. Integrin-mediated Signaling Events in Human Endothelial Cells. Mol. Biol. Cell 1998, 9, 1969–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horgan, C.C.; Rodriguez, A.L.; Li, R.; Bruggeman, K.F.; Stupka, N.; Raynes, J.K.; Day, L.; White, J.W.; Williams, R.J.; Nisbet, D.R. Characterisation of minimalist co-assembled fluorenylmethyloxycarbonyl self-assembling peptide systems for presentation of multiple bioactive peptides. Acta Biomater. 2016, 38, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Williams, C.G.; Wang, D.A.; Lee, H.; Manson, P.N.; Elisseeff, J. The effect of incorporating RGD adhesive peptide in polyethylene glycol diacrylate hydrogel on osteogenesis of bone marrow stromal cells. Biomaterials 2005, 26, 5991–5998. [Google Scholar] [CrossRef] [PubMed]
- Modepalli, V.N.; Rodriguez, A.L.; Li, R.; Pavuluri, S.; Nicholas, K.R.; Barrow, C.J.; Nisbet, D.R.; Williams, R.J. In vitro response to functionalized self-assembled peptide scaffolds for three-dimensional cell culture. Biopolymers 2014, 102, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Aye, S.-S.; Li, R.; Boyd-Moss, M.; Long, B.; Pavuluri, S.; Bruggeman, K.; Wang, Y.; Barrow, C.; Nisbet, D.; Williams, R. Scaffolds Formed via the Non-Equilibrium Supramolecular Assembly of the Synergistic ECM Peptides RGD and PHSRN Demonstrate Improved Cell Attachment in 3D. Polymers 2018, 10, 690. [Google Scholar] [CrossRef] [Green Version]
- Boyd-Moss, M.; Firipis, K.; O’Connell, C.D.; Rifai, A.; Quigley, A.; Boer, G.; Long, B.M.; Nisbet, D.R.; Williams, R.J. Shining a light on the hidden structure of gelatin methacryloyl bioinks using small-angle X-ray scattering (SAXS). Mater. Chem. Front. 2021, 5, 8025–8036. [Google Scholar] [CrossRef]
- Hammouda, B. Probing Nanoscale Structures—The Sans Toolbox; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2008; pp. 1–717. [Google Scholar]
- Wisotzki, E.I.; Tempesti, P.; Fratini, E.; Mayr, S.G. Influence of high energy electron irradiation on the network structure of gelatin hydrogels as investigated by small-angle X-ray scattering (SAXS). Phys. Chem. Chem. Phys. 2017, 19, 12064–12074. [Google Scholar] [CrossRef]
- Li, R.; Pavuluri, S.; Bruggeman, K.; Long, B.M.; Parnell, A.J.; Martel, A.; Parnell, S.R.; Pfeffer, F.M.; Dennison, A.J.C.; Nicholas, K.R.; et al. Coassembled nanostructured bioscaffold reduces the expression of proinflammatory cytokines to induce apoptosis in epithelial cancer cells. Nanomedicine 2016, 12, 1397–1407. [Google Scholar] [CrossRef] [Green Version]
- Loessner, D.; Stok, K.S.; Lutolf, M.P.; Hutmacher, D.W.; Clements, J.A.; Rizzi, S.C. Bioengineered 3D platform to explore cell-ECM interactions and drug resistance of epithelial ovarian cancer cells. Biomaterials 2010, 31, 8494–8506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria, O.M.; Maria, O.; Liu, Y.; Komarova, S.V.; Tran, S.D. Matrigel improves functional properties of human submandibular salivary gland cell line. Int. J. Biochem. Cell Biol. 2011, 43, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Luca, A.C.; Mersch, S.; Deenen, R.; Schmidt, S.; Messner, I.; Schäfer, K.-L.; Baldus, S.E.; Huckenbeck, W.; Piekorz, R.P.; Knoefel, W.T.; et al. Impact of the 3D Microenvironment on Phenotype, Gene Expression, and EGFR Inhibition of Colorectal Cancer Cell Lines. PLoS ONE 2013, 8, e59689. [Google Scholar] [CrossRef] [PubMed]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-Dimensional Cell Culture Systems and Their Applications in Drug Discovery and Cell-Based Biosensors. Assay Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Coll, J.L.; Adamson, E.D. Rescue of the mutant phenotype by reexpression of full-length vinculin in null F9 cells; effects on cell locomotion by domain deleted vinculin. J. Cell Sci. 1998, 111, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Guo, H.; Song, Y.; Zhao, X.; Shi, Y.; Lu, Y.; Hu, S.; Nie, Y.; Fan, D.; Wu, K. Loss of vinculin and membrane-bound beta-catenin promotes metastasis and predicts poor prognosis in colorectal cancer. Mol. Cancer 2014, 13, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toma-Jonik, A.; Widlak, W.; Korfanty, J.; Cichon, T.; Smolarczyk, R.; Gogler-Piglowska, A.; Widlak, P.; Vydra, N. Active heat shock transcription factor 1 supports migration of the melanoma cells via vinculin down-regulation. Cell. Signal. 2015, 27, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Hellman, K.; Alaiya, A.A.; Becker, S.; Lomnytska, M.; Schedvins, K.; Steinberg, W.; Hellström, A.C.; Andersson, S.; Hellman, U.; Auer, G. Differential tissue-specific protein markers of vaginal carcinoma. Br. J. Cancer 2009, 100, 1303. [Google Scholar] [CrossRef] [Green Version]
- Lifschitz-Mercer, B.; Czernobilsky, B.; Feldberg, E.; Geiger, B. Expression of the adherens junction protein vinculin in human basal and squamous cell tumors: Relationship to invasiveness and metastatic potential. Hum. Pathol. 1997, 28, 1230–1236. [Google Scholar] [CrossRef]
- Théry, M. Micropatterning as a tool to decipher cell morphogenesis and functions. J. Cell Sci. 2010, 123, 4201–4213. [Google Scholar] [CrossRef] [Green Version]
- Bays, J.L.; DeMali, K.A. Vinculin in cell–cell and cell–matrix adhesions. Cell. Mol. Life Sci. 2017, 74, 2999–3009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Chen, H.; Li, H.; Wu, Y. 3D culture increases pluripotent gene expression in mesenchymal stem cells through relaxation of cytoskeleton tension. J. Cell Mol. Med. 2017, 21, 1073–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benzoubir, N.; Mussini, C.; Lejamtel, C.; Santos, A.D.; Guillaume, C.; Desterke, C.; Samuel, D.; Bréchot, C.; Bourgeade, M.-F.; Guettier, C. Gamma-Smooth Muscle Actin Expression Is Associated with Epithelial-Mesenchymal Transition and Stem-Like Properties in Hepatocellular Carcinoma. PLoS ONE 2015, 10, e0130559. [Google Scholar]
- Li, H.; Li, Y.; Liu, D.; Sun, H.; Su, D.; Yang, F.; Liu, J. Extracellular HSP70/HSP70-PCs Promote Epithelial-Mesenchymal Transition of Hepatocarcinoma Cells. PLoS ONE 2013, 8, e84759. [Google Scholar] [CrossRef] [Green Version]
- Bolm, L.; Cigolla, S.; Wittel, U.A.; Hopt, U.T.; Keck, T.; Rades, D.; Bronsert, P.; Wellner, U.F. The Role of Fibroblasts in Pancreatic Cancer: Extracellular Matrix Versus Paracrine Factors. Transl. Oncol. 2017, 10, 578–588. [Google Scholar] [CrossRef]
- Apte, M.V.; Xu, Z.; Pothula, S.; Goldstein, D.; Pirola, R.C.; Wilson, J.S. Pancreatic cancer: The microenvironment needs attention too! Pancreatology 2015, 15, S32–S38. [Google Scholar] [CrossRef]
- Ulrich, T.A.; Jain, A.; Tanner, K.; MacKay, J.L.; Kumar, S. Probing cellular mechanobiology in three-dimensional culture with collagen-agarose matrices. Biomaterials 2010, 31, 1875–1884. [Google Scholar] [CrossRef]
- Firipis, K.; Footner, E.; Boyd-Moss, M.; Dekiwadia, C.; Nisbet, D.; Kapsa, R.M.I.; Pirogova, E.; Williams, R.J.; Quigley, A. Biodesigned bioinks for 3D printing via divalent crosslinking of self-assembled peptide-polysaccharide hybrids. Mater. Today Adv. 2022, 14, 100243. [Google Scholar] [CrossRef]
- Mani, V.; Lyu, Z.; Kumar, V.; Ercal, B.; Chen, H.; Malhotra, S.V.; Demirci, U. Epithelial-to-Mesenchymal Transition (EMT) and Drug Response in Dynamic Bioengineered Lung Cancer Microenvironment. Adv. Biosyst. 2019, 3, 1800223. [Google Scholar] [CrossRef]
- Penna, V.; Moriarty, N.; Wang, Y.; Law, K.C.L.; Gantner, C.W.; Williams, R.J.; Nisbet, D.R.; Parish, C.L. Extracellular Matrix Biomimetic Hydrogels, Encapsulated with Stromal Cell-Derived Factor 1, Improve the Composition of Foetal Tissue Grafts in a Rodent Model of Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 4646. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.C.; Grant, J.; Wang, Y.; Sandhu, M.; Williams, R.J.; Nisbet, D.R.; Perriman, A.W.; Lupton, D.W.; Jackson, C.J. Hydrogel-Immobilized Supercharged Proteins. Adv. Biosyst. 2018, 2, 1700240. [Google Scholar] [CrossRef]
- Firipis, K.; Boyd-Moss, M.; Long, B.; Dekiwadia, C.; Hoskin, W.; Pirogova, E.; Nisbet, D.R.; Kapsa, R.M.I.; Quigley, A.F.; Williams, R.J. Tuneable hybrid hydrogels via complementary self-assembly of a bioactive peptide with a robust polysaccharide. ACS Biomater. Sci. Eng. 2021, 7, 3340–3350. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Balushi, N.; Boyd-Moss, M.; Samarasinghe, R.M.; Rifai, A.; Franks, S.J.; Firipis, K.; Long, B.M.; Darby, I.A.; Nisbet, D.R.; Pouniotis, D.; et al. Self-Assembled Peptide Habitats to Model Tumor Metastasis. Gels 2022, 8, 332. https://doi.org/10.3390/gels8060332
Al Balushi N, Boyd-Moss M, Samarasinghe RM, Rifai A, Franks SJ, Firipis K, Long BM, Darby IA, Nisbet DR, Pouniotis D, et al. Self-Assembled Peptide Habitats to Model Tumor Metastasis. Gels. 2022; 8(6):332. https://doi.org/10.3390/gels8060332
Chicago/Turabian StyleAl Balushi, Noora, Mitchell Boyd-Moss, Rasika M. Samarasinghe, Aaqil Rifai, Stephanie J. Franks, Kate Firipis, Benjamin M. Long, Ian A. Darby, David R. Nisbet, Dodie Pouniotis, and et al. 2022. "Self-Assembled Peptide Habitats to Model Tumor Metastasis" Gels 8, no. 6: 332. https://doi.org/10.3390/gels8060332
APA StyleAl Balushi, N., Boyd-Moss, M., Samarasinghe, R. M., Rifai, A., Franks, S. J., Firipis, K., Long, B. M., Darby, I. A., Nisbet, D. R., Pouniotis, D., & Williams, R. J. (2022). Self-Assembled Peptide Habitats to Model Tumor Metastasis. Gels, 8(6), 332. https://doi.org/10.3390/gels8060332